

Volume1
.pdf58 Часть 1. Введение в мир клетки
Рис. 1.51. Дупликация гена как инструмент мутационного анализа для выяснения функций генов.
Вэтомгипотетическомпримерепредковыймногоклеточныйорганизмимеетгеном,содержащийединственнуюкопиюгенаG,которыйвыполняетсвоюфункциюнанесколькихучасткахтела,обозначенных зеленым. а) За счет дупликации гена современный потомок предкового организма имеет две копии генаG,названныеG1 иG2.Онивнекоторойстепениразошлисьпопрофилямэкспрессииипоуровням ихактивностинаучасткахэкспрессии,нопо-прежнемусохраняютважныечертысходства.Внекоторых участках они экспрессируются совместно и независимо друг от друга выполняют ту же самую старую функцию, что и предковый ген G (чередующиеся голубые и желтые полосы); на других участках они экспрессируютсяиндивидуальноимогутвыполнятьновыезадачи.б)Из-заналоженияфункцийпотеря одногоизэтихдвухгеноввследствиемутации(красныйкрестик)выявляеттолькочастьегороли;илишь припотереобоихгеноввдвойноммутантераскрываетсяполныйрепертуарпроцессов,закоторыеэти геныответственны.Аналогичныепринципыприменимыикдуплицированнымгенам,которыеработают в одном и том же месте (например, в одноклеточном организме), но приводятся в действие — вместе или индивидуально — в ответ на изменяющиеся условия. Таким образом, наличие дуплицированных геновсильноусложняетихгенетическийанализ.
лекулярной биологии позвоночных, и с ними мы будем неоднократно встречаться в этой книге.
1.3.16. Мышь как модель млекопитающих
По сравнению с дрозофилой у млекопитающих обычно в три или четыре раза больше генов, геном больше в 20 раз, а в телах их взрослых особей в миллионы или даже в миллиарды раз больше клеток. В отношении размера и функции генома, биологии клетки и молекулярных механизмов млекопитающие, тем не менее, чрезвычайно однородная группа организмов. Даже в анатомическом плане различия между млекопитающими, в основном, касаются размера и пропорций; трудно представить какую-либо часть тела человека, которая не имела бы своего аналога у слона и мыши, и наоборот. Эволюция свободно играет с количественными признаками, но не так легко изменяет закономерности, лежащие в основе строения.
Для более точной оценки того, насколько близко виды млекопитающих схожи друг с другом генетически, мы можем сравнивать последовательности нуклеотидов
Глава 1. Клетки и геномы 59
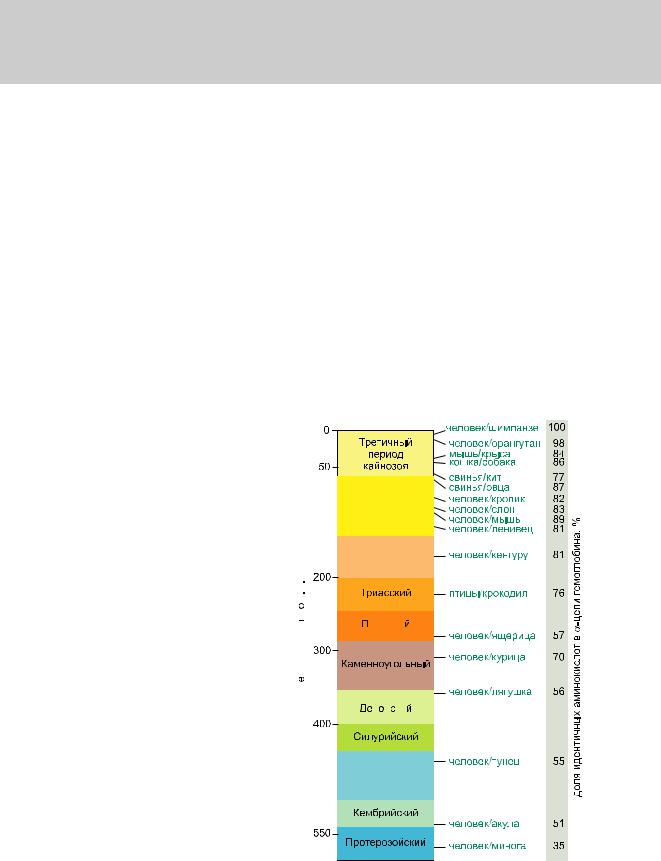
соответствующих (ортологичных) генов или же аминокислотные последовательности белков, кодируемых этими генами. Результаты для отдельных генов и белков имеют широкий диапазон значений. Но обычно, если мы выравниваем аминокислотную последовательность какого-либо белка человека с таковой ортологичного ему белка, принадлежащего, скажем, слону, то около 85 % аминокислот окажутся идентичными. Подобное сравнение между человеком и птицей показывает идентичность аминокислот приблизительно на уровне 70 % — вдвое больше различий, потому что линии родословной птиц и млекопитающих разошлись вдвое раньше, чем отошли друг от друга предки слона и человека (рис. 1.52).
Мышь в силу малого размера, выносливости и быстрого размножения стала лучшим модельным организмом для экспериментальных исследований в молеку-
Рис.1.52.Временнáяшкаладивергенцииразличныхпозвоночных.Нашкалеслевауказаноцениваемый периодвремениигеологическаяэражизнипоследнегообщегопредкакаждойуказаннойпарыживотных. Все временные оценки основаны на сравнении аминокислотных последовательностей ортологичных белков; чем дольше два животных эволюционировали независимо друг от друга, тем меньшая доля аминокислотостаетсяидентичной.Данныепомногимразличнымклассамбелковпришлосьусреднить, чтобыприйтикзаключительнымоценкам,авременнáяшкалаоткалиброванасучетомпалеонтологических данных, согласно которым последний общий предок млекопитающих и птиц жил
310 миллионов лет назад. Цифрами справа представленыданныеодивергенциипоследовательностейодногоопределенногобелка (выбранногонаугад)—α-цепигемоглобина. Обратитевнимание,чтодляэтогобелка,хотя
и наблюдается явная общая тенденция уве- 


 личения расхождения с течением времени, имеетсятакжеинескольконарушений.Они отражают хаотичность эволюционного про-
личения расхождения с течением времени, имеетсятакжеинескольконарушений.Они отражают хаотичность эволюционного про- 
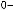 цесса и, вероятно, действие естественного
цесса и, вероятно, действие естественного 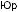
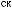
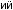 отбора, вызвавшего особенно быстрые изменения последовательности гемоглобина
отбора, вызвавшего особенно быстрые изменения последовательности гемоглобина  у некоторых организмов, которые испыты-
у некоторых организмов, которые испыты-  валиособыефизиологическиепотребности.
валиособыефизиологическиепотребности. 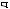
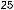
 В среднем, в пределах любой отдельно взя-
В среднем, в пределах любой отдельно взя- 
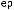

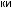 тойэволюционнойлинии,гемоглобинынака-
тойэволюционнойлинии,гемоглобинынака- 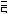
 пливаютизменениясоскоростьюоколо6-ти
пливаютизменениясоскоростьюоколо6-ти 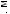 аминокислотныхзаменна100аминокислот
аминокислотныхзаменна100аминокислот  за каждые 100 миллионов лет. Некоторые
за каждые 100 миллионов лет. Некоторые 
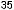
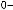
 белки,подвергнутыеболеестрогимфункциональным ограничениям, эволюционируют
белки,подвергнутыеболеестрогимфункциональным ограничениям, эволюционируют 

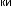 намногомедленнее,аиныев5разбыстрее. Всеэтопорождаетсерьезныесомнениявдостоверностивременн йоценкидивергенции,
намногомедленнее,аиныев5разбыстрее. Всеэтопорождаетсерьезныесомнениявдостоверностивременн йоценкидивергенции, 
 инекоторыеэкспертыполагают,чтоосновные
инекоторыеэкспертыполагают,чтоосновные 
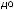
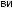
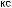

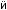 ветвимлекопитающихотошлидруготдруга на целых 60 миллионов лет позже, чем по-
ветвимлекопитающихотошлидруготдруга на целых 60 миллионов лет позже, чем по- 
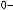

казаноздесь.(ПереработаноизS.Kumarand S. B. Hedges,Nature392:917–920,1998.Сраз-
решенияMacmillanPublishersLtd.)

60 Часть 1. Введение в мир клетки
лярной генетике позвоночных. Известно много происходящих в природе мутаций, которые являются зеркальным отражением эффектов, наблюдаемых при соответствующих мутациях у людей (рис. 1.53). Кроме того, разработаны методы, позволяющие проверить функцию любого выбранного нами гена мыши или любой некодирующей части генома мыши путем искусственного внесения мутаций в них; подробно мы объясним это позже.
Рис. 1.53. Человек и мышь: подобные гены и подобное развитие. Младенец человека и мышь, пока-
занныенаэтомрисунке,имеютподобныебелыепятнаналбу,потомучтообаимеютмутацииводном итомжегене(названномKit),необходимомдляразвитияиподдержанияпигментныхклеток.(Любез-
ностьR. A. Fleischman.)
Одна «сделанная под заказ» мышь-мутант может дать цитобиологу море информации. Можно установить эффекты заданной мутации в легионе различных контекстов, одновременно проверяя действие гена в различных видах клеток тела, которое, в принципе, может пострадать от этой мутации.
1.3.17. Человек сам являет миру свои особенности
Будучи людьми, мы проявляем особый интерес к геному человека. Мы хотим знать полный набор деталей, из которых мы собраны, и открыть механизм их работы. Но даже если бы мы были мышами, озабоченными молекулярной биологией мышей, люди были бы привлекательны для нас в качестве модельных генетических организмов благодаря лишь одной характерной особенности: в ходе медосмотров и жалобных исповедей перед врачами мы создаем каталог своих собственных генетических (и иных) расстройств. Человеческое население огромно, насчитывает на сегодняшний день приблизительно 6 миллиардов индивидов, а самодокументация, накопленная человечеством, означает, что собрана огромная база данных с информацией о мутациях у человека. Полная последовательность генома человека из более чем 3 миллиардов пар нуклеотидов уже определена, облегчая, как никогда прежде, опознание на молекулярном уровне именно того гена, который отвечает за интересующий нас мутантный признак у человека.
Сводя воедино данные исследований, проведенных на людях, мышах, мухах, червях, дрожжах, растениях и бактериях — используя подобия последовательностей генов, чтобы отобразить соответствия между тем или иным модельным организмом, — мы обогащаем свои знания по всем этим организмам.

Глава 1. Клетки и геномы 61
1.3.18. В деталях все мы различны
Что в точности мы имеем в виду, когда говорим о геноме человека? О чьем геноме идет речь? В среднем любые два человека, взятые наугад, отличаются примерно одной или двумя из 1 000 пар нуклеотидов в последовательности ДНК. В проекте «Геном человека» ДНК для секвенирования была произвольно выбрана из небольшой группы анонимных индивидов. Геном человека — геном биологического вида «человек», — строго говоря, это более сложная вещь, охватывающая полную совокупность различных вариантов генов, которые встречаются в человеческом населении и непрерывно обмениваются и перераспределяются в ходе полового размножения. В конечном счете мы можем надеяться задокументировать также
иэто разнообразие. Это поможет нам понять, например, почему некоторые люди предрасположены к одной болезни, а другие — к другой; почему одни хорошо переносят тот или иной лекарственный препарат, а другие — плохо. Оно также дает нам новые ключи к приоткрытию многих тайн нашей истории: переселению людей и смешению наших предков, инфекциям, которыми они страдали, рационам питания, которых они придерживались. Все эти моменты оставляют «следы» в различных формах генов, которые выжили в человеческих общинах.
Знание и понимание дают власть, позволяющую вмешиваться в биологию человека — чтобы избежать или предупредить болезнь, в биологию растений — чтобы получать более обильные урожаи, в биологию бактерий — чтобы использовать их для наших собственных нужд. Все эти биологические предприятия взаимосвязаны, потому что генетическая информация всех живых организмов написана на одном
итом же языке. Недавно обретенная молекулярными биологами способность читать
ирасшифровывать этот язык уже начала преобразовывать наши отношения с живым миром. Сведения по биологии клетки, приведенные в последующих главах, мы надеемся, подготовят читателя к ее пониманию, а возможно, и к тому, чтобы внести свой вклад в это невероятное по своим масштабам научное приключение
XXI века.
Заключение
Эукариотические клетки, по определению, хранят свою ДНК в отдельной,
окруженной мембраной полости — ядре. Кроме этого, у них есть цитоскелет для опоры и движения, замысловатые внутриклеточные полости для перева- ривания и выделения, способность (у многих видов) поглощать другие клетки и метаболизм, который зависит от окисления органических молекул митохон- дриями. Эти свойства предполагают, что эукариоты, возможно, зародились как хищники по отношению к другим клеткам. Митохондрии — как и хлоропласты у растений — имеют свой собственный генетический материал и, очевидно, эволюционировали от бактерий, которые были захвачены в цитоплазму клеткиэукариота и выжили как симбионты. В эукариотических клетках, как правило, в 3–30 раз больше генов, чем в прокариотических, и часто в тысячи раз больше
некодирующей ДНК. Некодирующая ДНК позволяет осуществлять сложное регулирование экспрессии генов, что необходимо для построения многоклеточных организмов. Многие эукариоты, однако, являются одноклеточными — среди них дрожжи Saccharomyces cerevisiae, которые служат простым модельным организмом для изучения цитобиологии эукариот, обеспечивая молекулярную базу консервативных фундаментальных процессов, таких как цикл клеточного

62 Часть 1. Введение в мир клетки
деления эукариот. Несколько других организмов выбраны в качестве основных моделей многоклеточных растений и животных, а секвенирование их полных геномов открыло путь к систематическому и всестороннему анализу функций генов, механизмов регулирования генов и генетического разнообразия. В резуль- тате дупликаций генов в ходе эволюции позвоночных их геномы содержат много- численные близкородственные гомологи большинства генов. Такая генетическая избыточность обусловила разнообразие и специализацию генов, приспособив их к выполнению новых задач, но вместе с тем она непомерно затрудняет рас- шифровку функций генов. В геномах нематоды Caenorhabditis elegans и мухи Drosophila melanogaster генетическая избыточность меньше, благоларя чему
эти два вида сыграли ключевую роль в раскрытии универсальных генетических механизмов развития животных.
Задачи
Какие утверждения являются правильными? Обоснуйте свой ответ
1.1.Гены гемоглобина человека, которые расположены двумя кластерами на двух хромосомах, представляют хороший пример набора ортологичных генов.
1.2.Горизонтальный перенос генов более распространен среди одноклеточных организмов, чем у многоклеточных.
1.3.Бóльшая часть последовательностей ДНК в бактериальном геноме кодирует белки, тогда как бóльшая часть последовательностей в геноме человека является некодирующей.
Обсудите следующие проблемы
1.4. С тех пор как генетический код был расшифрован четыре десятилетия назад, одни утверждают, что он, должно быть, представляет собой «замороженный случай», тогда как другие возражают, что он сформировался путем естественного отбора. Поразительная особенность генетического кода – это свойственное ему сопротивление воздействиям мутаций. Например, кодон с изменением в третьей позиции часто определяет ту же самую аминокислоту или другую, но с подобными химическими свойствами. Естественный код сопротивляется мутации более эффективно (то есть менее восприимчив к ошибкам), чем большинство других возможных кодов, как показано на рис. Q1.1. Только один из миллиона машинногенерируемых «случайных» кодов более устойчив к ошибкам, чем естественный генетический код. Говорит ли столь необычайная устойчивость генетического кода к мутациям в пользу его происхождения как «замороженного случая» или как
результата естественного отбора? Объясните ход ваших рассуждений.
Рис. Q1.1. Восприимчивость естественного (природного) кода относительно миллионов машинно-генерируемых кодов (задача 1.4). Восприимчивость является мерой среднего числа изменений в свойствах аминокислот, вызванных случайными мутациями. Низкое значение указывает, что мутации, как правило,вызываютнезначительныеизменения.(Данныелюбезно предоставленыSteveFreeland.)

Глава 1. Клетки и геномы 63
1.5.Вы начали характеризовать образец, добытый из глубин океанов Европы — одной из лун Юпитера. К вашему большому удивлению, образец содержит форму жизни, которая хорошо растет в питательном бульоне. Ваш предварительный анализ показывает, что это — клеточный организм, состоящий из ДНК, РНК
ибелка. Когда Вы показываете свои результаты коллеге, он заявляет, что ваш образец, скорее всего, был загрязнен организмом с Земли. Какие подходы Вы могли бы перепробовать, чтобы отличить загрязнение от новой клеточной формы жизни, основанной на ДНК, РНК и белке?
1.6.Не так трудно представить, что значит питаться органическими молекулами, которые производят живые организмы. То есть, в конечном счете, делать именно то, что делаем мы. Но что означает «питаться» солнечным светом, как это делают фототрофы? Или, что еще более странно, «питаться» камнями, чем занимаются
литотрофы? Что является «пищей», например, в смеси химикатов (H2S, H2, CO, Мn+, Fe2+, Ni2+, CH4 и NH4+), исторгающейся из гидротермального источника?
1.7.Сколько различных возможных деревьев (схем ветвления) с ветвями эубактерий, архей и эукариот получится вырастить на почве допущений о том, что все они произошли от некоторого общего предка?
1.8.Гены рибосомной РНК высококонсервативны (обладают относительно малой изменчивостью последовательности) у всех живущих на Земле организмов; таким образом, они эволюционировали очень медленно с течением времени. Говорит ли это о том, что гены рибосомной РНК были «рождены» совершенными?
1.9.Гены, участвующие в таких информационных процессах, как репликация, транскрипция и трансляция, переносятся между видами намного реже, чем гены, вовлеченные в метаболизм. В настоящее время причина такого неравенства еще не ясна, но есть одно предположение, согласно которому это связано с различным уровнем сложности информационных и метаболических процессов. Информационные процессы, как правило, вовлекают крупные ансамбли различных генных продуктов, тогда как метаболические реакции обычно катализируются ферментами, состоящими из единственного белка. Почему уровень сложности, заложенный в этих процессах — информационных или метаболических, — так или иначе влияет на скорость горизонтального переноса генов?
1.10.Процесс переноса генов из митохондриального генома в ядерный может быть проанализирован на растениях. Ген дыхательной цепи Cox2, который кодирует субъединицу 2 цитохромоксидазы, был функционально перенесен в ядро в ходе эволюции цветковых растений. Широкомасштабный анализ родов растений позволил точно установить время появления ядерной формы этого гена и определить несколько вероятных промежуточных продуктов на пути безвозвратного ухода из митохондриального генома. Сводные данные о распределении гена Cox2 между митохондриями и ядрами, наряду с данными об их транскрипции, представлены в филогенетическом контексте на рис. Q1.2.
A. Предполагая, что перенос митохондриального гена в ядро произошел только однажды (это предположение подтверждается структурами ядерных генов), укажите на филогенетическом дереве точку, в которой произошел этот перенос.
B. Известны ли какие-либо примеры родов, в которых как перенесенный ген, так и митохондриальный ген оказываются функционально активными? Приведите такие примеры.
C. Каково минимальное число событий инактивации или потери митохондриального гена? Укажите эти события на филогенетическом дереве.

64 Часть 1. Введение в мир клетки
Рис.Q1.2.Сводныеданныепораспределению гена Cox2 и сведения о соответствующих транскриптах в филогенетическом контексте (задача 1.10). Присутствие неповрежденного гена или функционально активного транскриптаобозначено(+);отсутствиенеповрежденногогенаилифункционально активноготранскриптаобозначено(–). Сокращения:мт—митохондрии;яд— ядра.
D. Каково минимальное
число событий инактивации или потери ядерного гена? Укажите эти события на филогенетиче-
ском дереве.
E. На основании этих дан-
ных предложите общую схему
передачи митохондриальных генов в ядерный геном.
1.11. Когда кодирующие гемоглобин гены были открыты в бобах, это было настолько удивительно – обнаружить в растении ген, типичный для крови животного, – что была выдвинута гипотеза, согласно кото-
рой этот ген растения возник в результате горизонтального переноса от животного. К настоящему времени секвенировано множество генов
гемоглобина, и филогенетическое дерево, основанное на некоторых из этих последовательностей, показано на рис. Q1.3.
A.Подтверждает ли это дерево или опровергает гипотезу о том, что гемоглобины растений возникли вследствие горизонтального переноса генов?
B.Если предположить, что кодирующие гемоглобин гены растений были первоначально получены, например, от паразитической нематоды, то как, по-вашему, будет выглядеть филогенетическое дерево?
1.12. Как известно, в различных ветвях генеалогического дерева скорость эволюции разная. Например, скорость эволюции в линии крыс значительно больше, чем в линии человека. Такая разница в скорости становится заметной, когда мы смотрим на изменения в белковых последовательностях, подвергающихся давлению естественного отбора, или на изменения в некодирующих последовательностях нуклеотидов, которые избавлены от заметного воздействия естественного отбора. Можете ли Вы предложить одно или несколько возможных объяснений более медленной скорости эволюционных изменений в ветви человека по сравнению
светвью крыс?

Глава 1. Клетки и геномы 65
Рис.Q1.3.Филогенетическоедеревогеновгемоглобинаразличныхвидов(кзадаче1.11).Бобовыевыделенырозовымцветом.
Литература
Общая
Alberts B., Bray D., Hopkin K. et al. (2004) Essential Cell Biology, 2nd ed. New York: Garland Science.
Barton N. H., Briggs D. E. G., Eisen J. A. et al. (2007) Evolution. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press.
Darwin C. (1859) On the Origin of Species. London: Murray.
Graur D. & Li W.-H. (1999) Fundamentals of Molecular Evolution, 2nd ed. Sunderland, MA: Sinauer Associates.
Madigan M. T. & Martinko J. M. (2005) Brock’s Biology of Microorganisms, 11th ed. Englewood Cliffs, NJ: Prentice Hall.
Margulis L. & Schwartz K. V. (1998) Five Kingdoms: An Illustrated Guide to the Phyla of Life on Earth, 3rd ed. New York: Freeman.
Watson J. D., Baker T. A., Bell S. P. et al. (2007) Molecular Biology of the Gene, 6th ed. Menlo Park, C. A: Benjamin-Cummings.
Клетки на Земле: характерные черты
Andersson S. G. E. (2006) The bacterial world gets smaller. Science 314: 259–
260.
Brenner S., Jacob F. & Meselson M. (1961) An unstable intermediate carrying information from genes to ribosomes for protein synthesis. Nature 190: 576–581.
Fraser C. M., Gocayne J. D., White O. et al. (1995) The minimal gene complement of Mycoplasma genitalium. Science 270: 397–403.

66 Часть 1. Введение в мир клетки
Harris J. K., Kelley S. T., Spiegelman et al. (2003) The genetic core of the universal ancestor. Genome Res. 13: 407–413.
Koonin E. V. (2005) Orthologs, paralogs, and evolutionary genomics. Annu. Rev. Genet. 39: 309–338.
Watson J. D. & Crick F. H. C. (1953) Molecular structure of nucleic acids. A structure for deoxyribose nucleic acid. Nature 171: 737–738.
Yusupov M. M., Yusupova G. Z., Baucom A. et al. (2001) Crystal structure of the ribosome at 5.5 Å resolution. Science 292: 883–896.
Разнообразие геномов и древо жизни
Blattner F. R., Plunkett G., Bloch C. A. et al. (1997) The complete genome sequence of Escherichia coli K-12. Science 277: 1453–1474.
Boucher Y., Douady C. J., Papke R. T. et al. (2003) Lateral gene transfer and the origins of prokaryotic groups. Annu. Rev. Genet. 37: 283–328.
Cole S. T., Brosch R., Parkhill J. et al. (1998) Deciphering the biology of Mycobacterium tuberculosis from the complete genome sequence. Nature 393: 537–544.
Dixon B. (1994) Power Unseen: How Microbes Rule the World. Oxford: Free-
man.
Kerr R. A. (1997) Life goes to extremes in the deep earth—and elsewhere? Sci- ence 276: 703–704.
Lee T. I., Rinaldi N. J., Robert F. et al. (2002) Transcriptional regulatory networks in Saccharomyces cerevisiae. Science 298: 799–804.
Olsen G. J. & Woese C. R. (1997) Archaeal genomics: an overview. Cell 89: 991–994.
Pace N. R. (1997) A molecular view of microbial diversity and the biosphere. Science 276: 734–740.
Woese C. (1998) The universal ancestor. Proc. Natl. Acad. Sci. USA 95: 6854–6859.
Генетическая информация эукариот
Adams M. D., Celniker S. E., Holt R. A. et al. (2000) The genome sequence of
Drosophila melanogaster. Science 287: 2185–2195.
Andersson S. G., Zomorodipour A., Andersson J. O. et al. (1998) The genome sequence of Rickettsia prowazekii and the origin of mitochondria. Nature 396: 133–140.
The Arabidopsis Initiative (2000) Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 408: 796–815.
Carroll S. B., Grenier J. K. & Weatherbee S. D. (2005) From DNA to Diversity: Molecular Genetics and the Evolution of Animal Design, 2nd ed. Maldon, MA: Blackwell Science.
de Duve C. (2007) The origin of eukaryotes: a reappraisal. Nature Rev. Genet. 8: 395–403.
Delsuc F., Brinkmann H. & Philippe H. (2005) Phylogenomics and the reconstruction of the tree of life. Nature Rev. Genet. 6: 361–375.
DeRisi J. L., Iyer V. R. & Brown P. O. (1997) Exploring the metabolic and genetic control of gene expression on a genomic scale. Science 278: 680–686.
Gabriel S. B., Schaffner S. F., Nguyen H. et al. (2002) The structure of haplotype blocks in the human genome. Science 296: 2225–2229.

Глава 1. Клетки и геномы 67
Goffeau A., Barrell B. G., Bussey H. et al. (1996) Life with 6000 genes. Science
274:546–567.
International Human Genome Sequencing Consortium (2001) Initial sequencing
and analysis of the human genome. Nature 409: 860–921.
Kellis M., Birren B. W. & Lander E. S. (2004) Proof and evolutionary analysis of ancient genome duplication in the yeast Saccharomyces cerevisiae. Nature 428: 617–624.
Lynch M. & Conery J. S. (2000) The evolutionary fate and consequences of duplicate genes. Science 290: 1151–1155.
Mulley J. & Holland P. (2004) Comparative genomics: Small genome, big insights. Nature 431: 916–917.
National Center for Biotechnology Information. http://www.ncbi.nlm.nih.gov/ Owens K. & King M. C. (1999) Genomic views of human history. Science 286:
451–453.
Palmer J. D. & Delwiche C. F. (1996) Second-hand chloroplasts and the case of
the disappearing nucleus. Proc. Natl. Acad. Sci. USA 93: 7432–7435. Pennisi E. (2004) The birth of the nucleus. Science 305: 766–768. Plasterk R. H. (1999) The year of the worm. BioEssays 21: 105–109.
Reed F. A. & Tishkoff S. A. (2006) African human diversity, origins and migrations. Curr. Opin. Genet. Dev. 16: 597–605.
Rubin G. M., Yandell M. D., Wortman J. R. et al. (2000) Comparative genomics of the eukaryotes. Science 287: 2204–2215.
Stillman B. & Stewart D. (2003) The genome of Homo sapiens (Cold Spring Harbor Symp. Quant. Biol. LXVIII). Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press.
The C. elegans Sequencing Consortium (1998) Genome sequence of the nematode C. elegans: a platform for investigating biology. Science 282: 2012–2018.
Tinsley R. C. & Kobel H. R. (eds.) (1996) The Biology of Xenopus. Oxford: Clarendon Press.
Tyson J. J., Chen K. C. & Novak B. (2003) Sniffers, buzzers, toggles and blinkers: dynamics of regulatory and signaling pathways in the cell. Curr. Opin. Cell Biol. 15: 221–231.
Venter J. C., Adams M. D., Myers E. W. et al. (2001) The sequence of the human genome. Science 291: 1304–1351.