

91
стероидные гормоны.
∙Защитная функция: липиды создают механическую защиту внутренних органов от повреждений и травм.
∙Терморегуляторная функция: липиды подкожной клетчатки снижают теплоотдачу организма.
∙Участвуют в проведении нервных импульсов, формируют миелиновые оболочки нервных пучков, играющие роль «электроизолятов».
∙Липиды растворяют жирорастворимые витамины.
∙Липиды являются важными источниками эндогенной воды.
7.5.Структура и функции клеточных мембран
Состав клеточных мембран. В состав клеточных мембран в различных соотношениях входят белки, жиры и углеводы. На долю белков в среднем приходится 50%, липидов - 30%, углеводов - 10%.
Белки представлены ферментами, структурными, транспортными, рецепторными белками. Около половины липидов мембран составляют глицерофосфолипиды, треть приходится на холестерин, меньшая часть - на сфинголипиды. Углеводы клеточных мембран представлены компонентами гликосфинголипидов, гликопротеидов.
Структура клеточных мембран. В настоящее время общепринятой является мозаичная структура клеточной мембраны. Согласно этой модели, основу клеточной мембраны составляют глицерофосфолипиды, которые ориентированы в мембране таким образом, что гидрофильные участки находятся на поверхности, а гидрофобные в глубине клеточной мембраны. В силу дифильности глицерофосфолипиды образуют билипидный слой. Фосфолипиды в клеточных мембранах располагается ассимитрично, на поверхности плазматической мембраны находятся в основном фосфатидилхолин, а внутри фосфатидилколамин и фосфатидилсерин.
Белки в клеточных мембранах делятся на поверхностные белки и интергральные. Интегральные белки обычно расположены в мембране асимметрично. Толщину мембраны пронизывает гидрофобные участки белка, чаще всего уложенные в виде альфа - спирали, С-конец полипептидной цепи находится на внутренней поверхности, а N-конец на внешней поверхности мембраны. Очень часто к N-концевому фрагменту присоединяются углеводы, выполняющие рецепторную функцию. Гидрофобные части белка связываются с гидрофобными участками липидов, а гидрофильные с гидрофильными участками липидов.
Физико-химические свойства мембран определяются химическим составом мембран и температурой окружающей среды. Жёсткость мембранам придают холестерин и насыщенные жирные кислоты. Непредельные жирные кислоты определяют текучесть липидов клеточной мембраны. При низкой температуре фосфолипиды достаточно жёстко зафиксированы в составе мембраны, при повышении температуры возможно перемещение липидов. При температуре тела липиды находятся в жидко – кристаллическом состоянии.
Функции клеточных мембран
1. Разделительная функция – мембраны придают форму клеткам,

92
формируют внутренние отсеки, взаимодействуют со структурой цитоскелета.
2.Коммуникативная функция – мембраны обеспечивают межклеточные контакты с помощью рецепторов.
3.Метаболическая функция – в клеточные мембраны встроены мембранные ферменты.
4.Транспортная функция – через мембрану осуществляется транспорт веществ.
5.Рецепторная функция – избирательное взаимодействие рецепторов мембран с различными веществами.
Транспорт веществ через клеточные мембраны
1.Пассивный транспорт веществ, который осуществляется по градиенту концентрации через соответствующие мембранные каналы.
2.Активный транспорт против градиента концентрации с использованием энергии АТФ.
3.Облегчённый транспорт, в котором участвуют особые дополнительные транспортные белки, осуществляющие или однонаправленное перемещение двух веществ, или разнонаправленное перемещение двух веществ через мембрану.
4.Транспорт макромолекул осуществляется путём эндоцитоза или экзоцитоза.
7.6.Переваривание липидов
Для взрослого человека суточная потребность в липидах составляет от
70-80 г до 100 г, для детей 5 – 7 г/кг.
У взрослых людей процесс переваривания липидов происходит в тонком кишечнике. Необходимыми условиями для этого являются:
-наличие ферментов;
-оптимальное рН;
-эмульгирование жиров.
Необходимость эмульгирования липидов связана с водонерастворимостью жиров. Водорастворимые ферменты могут действовать на липиды только на поверхности капли. Эмульгирование повышает поверхность раздела липид / вода и обеспечивает большую поверхность контакта фермента и липида. В эмульгировании липидов основную роль играют жёлчные кислоты, выделяемые в просвет кишечника в составе жёлчи.
Различают простые и парные, первичные и вторичные жёлчные кислоты: Простые жёлчные кислоты являются производными холановой кислоты.
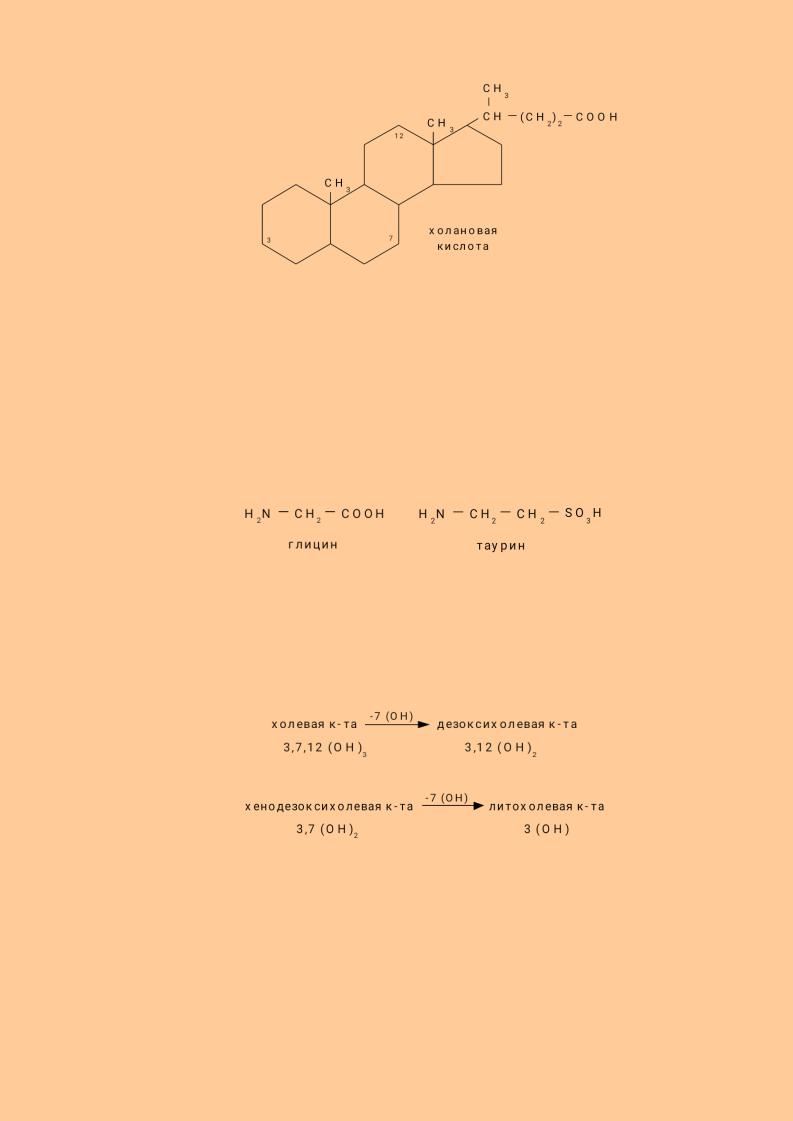
93
К простым жёлчным кислотам относятся холевая, дезоксихолевая кислота, хенодезоксихолевая и литохолевая кислоты.
Синтез жёлчных кислот из холестерина происходит в печени. Ключевым ферментом является 7 – α -гидроксилаза. Она переводит холестерин при участии цитохрома Р450 в 7-α - холестерин - 3,7 (ОН)2. Он, в свою очередь, переходит в хенодезоксихолевую кислоту - 3,7 (ОН)2 путём укорочения бокового радикала и в холевую кислоту - 3,7,12 (ОН)3. Эти две кислоты являются первичными жёлчными кислотами.
Полярность простых жёлчных кислот увеличивается при образовании парных жёлчных кислот путём присоединения глицина (гликокола) и таурина.
У взрослого человека до 80% всех жёлчных кислот представлено гликохолевой и таурохолевой кислотами. В кишечнике под действием микрофлоры происходит отцепление таурина, гликокола и ОН группы в 7 положении с образованием вторичных желчных кислот: дезоксихолевой и литохолевой.
Все жёлчные кислоты относятся к поверхностно активным веществам, имеющим в своем составе гидрофобные и гидрофильные участки. Гидрофильными являются ОН - группы, остатки таурина и гликокола, а гидрофобными – радикал жёлчной кислоты. Благодаря дифильности жёлчные кислоты располагаются в поверхностном слое жировой капли и уменьшают поверхностное натяжение.

94
Врезультате снижения поверхностного натяжения под действием перистальтики кишечника, выделения СО2 в результате нейтрализации бикарбонатами соляной кислоты желудочного содержимого происходит дробление крупных капель липида на множество мелких – эмульгирование, резко возрастает поверхность соприкосновения липидных капель и ферментов.
Липолитические ферменты, участвующие в переваривании липидов, активны при pН 8 – 8,5. Такая среда обеспечивается секрецией бикарбонатов поджелудочной железой.
Основные ферменты переваривания липидов вырабатываются поджелудочной железой и стенкой тонкого кишечника.
Впереваривании ТАГ участвует поджелудочная липаза. Она вырабатывается в неактивной форме, и в тонком кишечнике взаимодействует с дополнительным белком колипазой, который повышает активность липазы и обеспечивает контакт фермента с соответствующими жирами. Поджелудочная липаза отщепляет последовательно остатки жирных кислот из α - положений с образованием β – моноацилглицерина (β -МАГ)
Образующиеся бета-МАГ в случае изомеризации в α –МАГ могут в дальнейшем подвергаться расщеплению под действием липазы до глицерина и жирных кислот. Около 50% МАГ подвергается всасыванию.
Переваривание глицерофосфолипидов происходит под действием ферментов фосфолипаз поджелудочной железы. Под действием фосфолипазы А2 отщепляется остаток жирной кислоты из β – положения с образованием продукта неполного распада глицерофосфолипида – лизофосфолипида, содержащего в β – положении гидрофильную гидроксильную группу. Лизофосфолипиды являются поверхностно активными веществами и усиливают процессы эмульгирования жиров. Лизофосфолипиды расщепляются лизофосфолипазой на жирную кислоту и глицерофосфохолин, который хорошо всасывается без гидролиза. Таким образом, при распаде глицерофосфолипидов образуются жирные кислоты и глицерофосфохолин.

95
Эфиры холестерина расщепляются ферментом холестеролэстеразой.
Переваривание сфинголипидов осуществляется ферментами эстеразами, фосфатазами, амидазами, гликозидазами.
7.7. Всасывание продуктов расщепления липидов Всасывание продуктов расщепления липидов происходит в тонком
кишечнике и определяется водорастворимостью или водонерастворимостью образовавшихся продуктов. Водорастворимые вещества (глицерин, холин, Н3РО4) легко всасывается по градиенту концентрации.
Водонерастворимые вещества (β-МАГ, холестерин, длинноцепочечные жирные кислоты) не могут всасываться самостоятельно. В их всасывании участвуют жёлчные кислоты, которые образуют в просвете кишечника особые сферические водорастворимые структуры – мицеллы, в которые встраиваются гидрофобные жирные кислоты, холестерин. В процессе всасывания мицеллы распадаются, все водонерастворимые вещества подвергаются всасыванию. Жёлчные кислоты частично возвращаются в просвет кишечника, но в основном проходят процесс гемато-гепато-энтеральной циркуляции: всасываются, с током крови возвращаются в печень и повторно выделяются в состав жёлчи в просвет кишечника. Вследствие многократной циркуляции жёлчных кислот их небольшого количества (4 - 6 г) оказывается достаточным для всасывания большого количества гидрофобных продуктов. 10% короткоцепочечных жирных кислот может всасываться в виде тончайшей эмульсии путём иноцитоза.
7.8. Ресинтез липидов в слизистой тонкого кишечника Ресинтез – синтез липидов, характерных для организма человека из
компонентов переваривания липидов пищи. Недостающие жирные кислоты, спирты, необходимые для ресинтеза, могут синтезироваться в клетках слизистой кишечника (энтероцитах) и выделяться в составе жёлчи. Ресинтез триацилглицеринов происходит из активной формы глицерина и жирных кислот последовательно через стадию моноацилглицерина, диацилглицерина. Ресинтез

96
глицерофосфолипидов происходит из фосфатидной кислоты, фосфохолина и диацилглицеринов. Ресинтезу подвергаются и другие виды липидов.
7.9. Особенности переваривания и всасывании липидов у детей Главная особенность переваривания липидов в раннем детском возрасте
заключается в том, что примерно половина их частично расщепляется в желудке. Данная особенность обусловлена следующими обстоятельствами:
1.липиды молока находится в эмульгированном состоянии;
2.при грудном вскармливании в переваривании липидов участвует липаза грудного молока;
3.в процессе сосания у грудного ребёнка вырабатываются лингвальная и фарингальная липазы, которые оказывают эффект в желудке;
4.активно вырабатывается желудочная липаза с оптимумом рН около 5,0;
5.у детей в желудке менее кислая среда, приближенная к оптимуму рН для липаз;
6.активность панкреатической липазы у детей снижена;
7.в детском возрасте менее активен синтез жёлчных кислот, повышена их потеря через кишечник и замедлена гепато – энтеральная циркуляция.
Всасывание продуктов гидролиза жиров у детей происходит с большей скоростью, чем у взрослых в связи с высокой проницаемостью слизистой кишечника, а также в связи с наличием в липидах грудного молока большого количества среднецепочечных жирных кислот.
7.10. Транспорт липидов кровью Гидрофобные липиды не могут транспортироваться кровью
самостоятельно. Они переносятся в следующих формах:
1.липопротеиды (липопротеины) – белково-липидные комплексы;
2.хиломикроны –липидные капли, образующиеся в млечном соке;
3.свободные жирные кислоты транспортируются в комплекте с альбуминами.
Хиломикроны - это мельчайшие капельки липида с размерами около 500 нм, плотностью 0,95 г/см3, состоящие из 2% белка и 90% ТАГ. Хиломикроны синтезируются в слизистой кишечника, считаются транспортной формой пищевых (экзогенных) липидов в организме. Хиломикроны попадают сначала в лимфу, а затем разносятся кровью в основном в жировые депо (>50%), а также в печень, лёгкие, мышечную ткань.
Липопротеиды (ЛП) являются основной транспортной формой липидов. По электрофоретической подвижности различают: пре β - ЛП, β - ЛП, α - ЛП По плотности выделяют:
-ЛП очень низкой плотности (ЛПОНП);
-ЛП низкой плотности (ЛПНП);
-ЛП высокой плотности (ЛПВП);
-ЛП промежуточной плотности;
-ЛП очень высокой плотности.
Все ЛП построены по общему принципу. В центре частицы находится гидрофобное ядро, в которое входят ТАГ и эфиры холестерина (Эх), вокруг него

97
формируется гидрофильная оболочка, в которую входят фосфолипиды (ФЛ), холестерин (Х-ОН). На поверхности располагаются белки - апопротеины (АроPt).
Различают несколько видов АроPt: A, B, C, E. Они формируют структуру липопротеидных частиц, взаимодействуют с тканевыми рецепторами к ЛП, являются активаторами ферментов обмена ЛП.
ЛП осуществляют транспорт липидов, жирорастворимых витаминов и гидрофобных гормонов.
Закономерности строения липопротеидов в ряду: ЛПОНП →ЛПНП→ЛПВП представлены в таблице.
Липопротеиды
|
ЛПОНП |
ЛПНП |
ЛПВП |
|
Размеры |
80 нм |
25 нм |
10 нм |
|
уменьшаются |
||||
|
|
|
||
Плотность |
1,006 г/см3 |
1,006-1,06 г/см3 |
1,25 г/см3 |
|
возрастает |
|
|
|
|
% содержания |
|
|
|
|
белков |
10% |
25% |
50-60% |
|
увеличивается |
|
|
|
|
% содержания |
|
45-48% |
30% холестерин |
|
липидов |
60% ТАГ |
|||
холестерин |
30% ФЛ |
|||
уменьшается |
|
|||
|
|
|
ЛПОНП – синтезируется в печени, считаются основной транспортной формой эндогенных липидов. В эндотелии сосудов ЛПОНП и хиломикроны подвергаются действию фермента липопротеидной липазы, которая расщепляет в их составе ТАГ. В результате в составе ЛП повышается доля холестерина, и ЛПОНП превращаются в ЛПНП.
ЛПНП считаются транспортной формой холестерина от печени к органам и тканям. В тканях имеются рецепторы к ЛПНП, при участии которых происходит поглощение холестерина с последующим использованием его на построение мембран, синтез стероидов, депонированием в виде эфиров.

98
ЛПВП синтезируется в печени в виде дисковидных структур. Они считается транспортной формой холестерина из тканей к печени. В кровотоке при контакте с эндотелием происходит поглощение холестерина из тканей и переход его в ЛПВП. Они постепенно превращаются в сферические структуры и переносят холестерин в печень. В поглощении холестерина частицами ЛПВП участвует фермент ЛХАТ (лецитинхолестеролацилтрансфераза), который в составе ЛПВП переносит остатки жирных кислот с фосфолипидов на холестерин с образованием эфиров холестерина. Эфиры холестерина более гидрофобны по сравнению со свободным холестерином и, в силу этого, погружаются внутрь ЛП частицы.
У детей общее содержание ЛП ниже, чем у взрослых людей. В детском возрасте снижена концентрация хиломикронов и ЛПОНП, но повышено содержание ЛПВП, в которых выше доля гидрофильных компонентов.
Содержание липидов в крови взрослых людей
Общие липиды |
4-8 г/л |
ТАГ |
1-2,5 ммоль/л |
ФЛ |
2,5-3,0 ммоль/л |
Холестерин |
3,5-5,2 ммоль/л |
Свободные жирные |
0,5 – 1,0 мэкв/л |
кислоты |
|
У детей содержание общих липидов ниже, чем у взрослых. В то же время концентрация свободных жирных кислот у детей повышено примерно в два раза.
Большая часть переносимых кровью липидов откладывается в жировых депо, к которым относятся подкожно-жировая клетчатка, большой и малый сальники. У детей наиболее активно депонирование липидов происходит в возрасте 1 года, 7 лет и в пубертатном периоде. В раннем детском возрасте у детей важным видом жировой ткани является бурая жировая ткань. Она локализована в основном на спине, на груди, имеет бурый оттенок, который обусловлен большим содержанием митохондрий и Fе - содержащих цитохромов. В бурой жировой ткани происходит нефосфорилирующее окисление липидов, которое сопровождается выделением тепловой энергии (данная ткань является органом термогенеза). Жировое депо у детей легко истощается при нарушении питания, болезнях, стрессе. Липиды в жировых депо постоянно обновляются.
7.11. Обмен триацилглицеринов
7.11.1. Распад триацилглицеринов в тканях (липолиз) Триацилглицерины поэтапно расщепляется тканевыми липазами.
Ключевым ферментом липолиза является гормональнозависимая ТАГ-липаза. Образующиеся на этом этапе распада жиров глицерин и жирные кислоты окисляются в тканях с образованием энергии.

99
7.11.1.1. Окисление жирных кислот.
Различают несколько вариантов окисления жирных кислот: α - окисление, β - окисление, ω - окисление. Основным вариантом окисления жирных кислот является β - окисление. Оно наиболее активно протекает в жировой ткани, печени, почках и сердечной мышце.
β - окисление заключается в постепенном отщеплении от жирной кислоты двух углеродных атомов в виде ацетил - КоА с освобождением энергии. Запас жирных кислот в клетках сосредоточен в цитозоле, где протекает активация жирных кислот с образованием ацил - КоА
Сам процесс β-окисления ацил-КоА происходит в митохондриях. Митохондриальная мембрана непроницаема для длинноцепочечных ацил - КоА. В переносе их внутрь митохондрий участвует специальный переносчик карнитин (метил, гидропроизводное аминомасляной кислоты). Ацил - КоА образует с карнитином комплекс, который после переноса жирной кислоты внутрь митохондрий распадается.
Энергетическая эффективность бета - окисления жирных кислот складывается из энергии окисления ацетил - КоА в цикле Кребса и энергии, освобождающейся в самом бета – цикле. Энергия окисления жирной кислоты тем выше, чем длиннее её углеродная цепь. Количество молекул ацетил - КоА из насыщенной жирной кислоты и количество образующихся из них молекул АТФ определяется по формулам:
n=N/2, где n-количество молекул ацетил - КоА, N- число атомов углерода в жирной кислоте.
Количество молекул АТФ за счёт окисления молекул ацетил-КоА = (N/2)*12 Число β - циклов окисления на один меньше, чем количество образующихся молекул ацетил-КоА, поскольку в последнем цикле масляная кислота за один цикл переходит в две молекулы ацетил-КоА, и рассчитывается
по формуле
Количество β -циклов = (N/2)-1
Количество молекул АТФ в β - цикле рассчитывается, исходя из последующего окисления образовавшихся в нём НАДН2 (3 АТФ) и ФАДН2 (2 АТФ) по формуле
Количество молекул АТФ, образующихся в β - циклах = ((N/2)-1)*5
2 макроэргические связи АТФ расходуются на активацию жирной кислоты
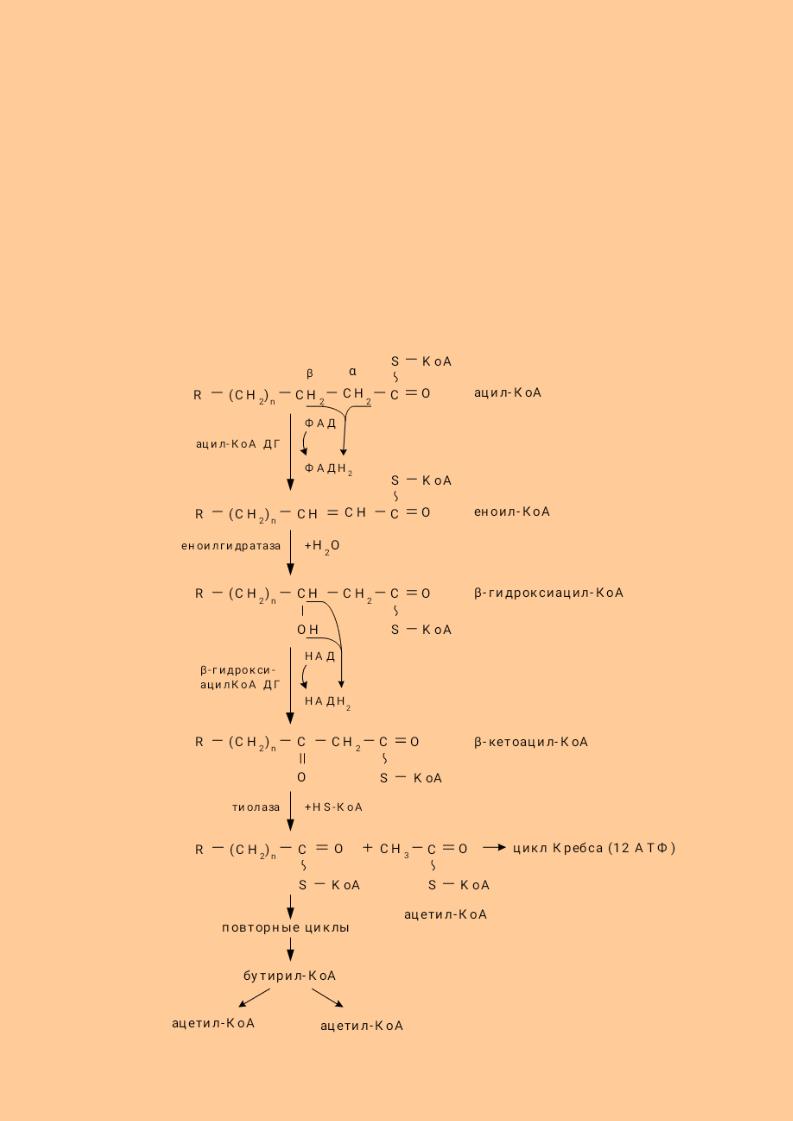
100
Суммарная формула для подсчёта выхода АТФ при окислении насыщенной жирной кислоты имеет вид: 17(N/2)-7.
Окисление ненасыщенных жирных кислот на начальных стадиях представляет обычное β - окисление до места двойной связи. Если эта двойная связь находится в β - положении, то продолжается окисление жирной кислоты со второго этапа (минуя стадию восстановления ФАД→ ФАДН2). Если двойная связь находится не β - положении, то ферментами еноилтрансферазами связь перемещается в β – положение. Таким образом, при окислении ненасыщенных жирных кислот образуется меньше энергии (теряется образование ФАДН2 на каждую двойную связь). Она рассчитывается по формуле:
7(N/2)-7-2m, где m-число двойных связей.
Химизм β -окисления насыщенных жирных кислот