

6 курс / Медицинская реабилитация, ЛФК, Спортивная медицина / Физиотерапия, лазерная терапия / Многоканальная_электростимуляция_Давиденко_В_Ю_
.pdfИз литературы, посвященной физическим нагрузкам, хорошо известно, что тип тренировки скелетных мышц зависит от характера выполняемой работы. Так, если просто поднимать вес несколько раз в день, то мышечная сила не возрастает. Для того, чтобы эта сила увеличилась, необходима нагрузка с возрастающим сопротивлением. Тяжелоатлет, например, для увеличения мышечной силы должен очень медленно поднимать и опускать вес, равный, примерно, 2/3 максимально возможного усилия. Именно такая (изокинетическая) нагрузка оптимальна для повышения силы скелетных мышц, т.к. характер требований, предъявляемых к метаболическим процессам в мышцах, может играть первостепенную роль в тех биохимических изменениях, которые наступают в них при длительной физической тренировке. Если тяжелоатлет будет поднимать карандаш даже несколько сот раз в день, то увеличения силы не произойдет.
Еще в 1941 г. в своих ранних исследованиях Нобелевский лауреат по электрофизиологии Дж. Экклз [20] на обширном экспериментальном материале доказал, что качество электростимуляционного воздействия на мышцы зависит от того, насколько физиологично проводится это воздействие. Так, даже 2-х часовая электростимуляция (в день) тенотомированной и укороченной мышцы не предотвращает атрофических процессов в ней. В то время, как при обеспечении условия удлиннения и укорочения мышцы, для предупреждения атрофических процессов достаточно стимулировать эти мышцы несколько минут в день.
Для более глубокого понимания механизма действия ЭС при выборе оптимальных режимов воздействия очень важно выявить биохимические и морфофункциональные сдвиги при различных сроках электростимуляции.
Всвязи с этим нами [21] на кафедре физиологии НУФК, совместно с кафедрой функциональной анатомии и отделом биохмии НАНУ были проведены исследования, целью которых было изучение в эксперименте на животных функциональных сдвигов в мышцах при различной продолжительности эксперимента и длительности сеансов ЭС.
4.3АТФ-азная активность и интенсивность накопления Са2+ при различной продолжительности электростимуляции
Впроведенных исследованиях с целью характеристики состояния обменных процессов определяли содержание аденозинтрифосфорной кислоты (АТФ), креатинфосфата (КФ) и неорганического фосфата в мышечной ткани. Исходя из важной роли саркоплазматического ретикулума мышечного волокна
в процессе сокращения-расслабления изучали также АТФ-азную активность и потребление кальция (Са 2+) в этой структуре.
Исследования проводили на взрослых кроликах весом 1800-2500 г. Стимуляция осуществлялась с помощью электростимулятора ПМС-2. Длительность одиночного импульса 0,7-0,8 мс. Длительность фазы модулированного сигнала 2 с (период напряжения 0.6-0.8 с, расслабления – 1.2- 1.3 с, т.е. 30 сокращений в минуту). Стимулировали мышцы правого бедра
80

кроликов. Левая конечность служила контролем. Первый сеанс длился 7 минут, остальные по 10 минут. Раздражения были сверхпорпоговыми, но не вызывали оборонительной реакции животных. Применяли следующие режимы стимуляции: 18 сеансов, 5 сеансов, 2 раза по 5 сеансов с двухдневным перерывом после 5-и сеансов и четырех-дневным отдыхом после второго курса стимуляции.
Фрагменты саркоплазматического ретикулума (ФСР) выделяли из мышц кролика по [22,23,24 ] .
Количество белка, АТФ-азную активность и интенсивность накопления Са2+ – по [25,26]. Содержание АТФ и креатинфосфата определяли по [27,28]. Интенсивность накопления Са2+ выражали в микромолях Са2+ на 1 миллиграмм белка за одну минуту, содержание АТФ и креатинфосфата – в миллиграммпроцентах на 1 г свежей ткани. Все данные обработаны статистически.
Для изучения возбудимости мышц определяли реобазу и хронаксию с помощью хронаксиметра ИС-1.
Для гистологических исследований материал фиксировали в 10%-ом формалине и системе спирт-формалин (2:1). Срезы обрабатывали гематокси- лин-эозином, пикрофусином и импрегнировали азотнокислым серебром по Бильшовскому или Бильшовскому-Гросс [29].
Прежде всего мы исследовали накопление Са2+, АТФ-азную активность саркоплазматического ретикулума, содержание АТФ и КФ тканей мышц. Как видно из таблицы 4.1, все исследованные показатели скелетных мышц правой и левой конечности нормальных кроликов не отличаются один от другого ( >0.5) .
Таблица 4.1. Накопление Са2+ и АТФ-азная активность в фрагментах саркоплазмати-
ческого ретикулума, содержание АТФ и креатинфосфата в симметричных мышцах нормальных кроликов /M m/.
Исследуемый |
Правая |
Левая |
||
показатель |
конечность |
конечность |
||
Интенсивность |
1,2 |
0,15 |
1,2 |
0,3 |
накопления Са2+ |
||||
АТФ-азная активность |
1,9 |
0,3 |
2,0 |
0,2 |
Содержание АТФ |
36,8 |
1,7 |
35,8 |
2,2 |
Содержание креатинофосфата |
19,7 |
2,2 |
18,2 |
1,0 |
В следующей серии были исследованы мышцы кроликов, у которых стимулировали мышцы правого бедра в течение 18-и дней.
Данные этой серии исследований указывают на то, что накопление Са2+, АТФ-азная активность ФСР, содержание АТФ и КФ стимулируемых мышц существенно не изменяется при сравнении с симметричными мышцами
( >0.5).
Однако, если учитывать, что тренировки вызывают изменения не только в стимулируемых мышцах, но и в других органах и тканях [30], то полученные
81

данные следует сравнивать с одноименными мышцами контрольных животных.
Такое сопоставление (табл. 4.2) показывает, что содержание Са2+ ФСР понижено по сравнению с нормой как в стимулируемых мышцах ( <0.01), так и в симметричных мышцах другой конечности ( =0,01). Кроме того, в мышцах обеих конечностей подопытных животных значительно снижено количество АТФ ( =0,001).
Для оценки возбудимости электростимулируемых мышц измерялись реобаза и хронаксия (рис. 4.1). В фоновых исследованиях реобаза была в пределах 6 - 133 вольт, хронаксия – 0,22 - 0,33 мс.
Одноразовая стимуляция вызывала снижение реобазы и хронаксии. Это отчетливо проявлялось и дальше в течение 4-5 сеансов. Уровень возбудимости мышц превышал фоновые показатели до 30 %. Таким образом, на протяжении 5-и сеансов наблюдалось повышение возбудимости мышц. Начиная с 6-го сеанса, величина реобазы повышается и возвращается к исходному уровню, повышение же хронаксии даже его превышает.
Таблица 4.2. Накопление Са2+ и АТФ - азная активность в фрагментах саркоплазма-
тического ретикулума, содержание АТФ и креатинфосфата в скелетных мышцах кроликов после 18-и дневной электростимуляции /М m/.
Исследуемый показа- |
Стимулированная |
Симметричная к сти- |
Нормальная |
||||
тель |
мышца (n=6) |
мулированной (n=6) |
мышца (n=9) |
||||
Интенсивность накоп- |
0,82 |
0,1 |
0,7 |
0,1 |
1,2 |
0,1 |
|
ления Са2+ |
|||||||
|
|
|
|
|
|
||
АТФ-азная активность |
1,2 |
0,4 |
1,35 |
0,36 |
2,0 |
0,1 |
|
Содержание АТФ |
15,7 |
1,9 |
17,4 |
1,4 |
36,3 |
1,0 |
|
Содержание креатино- |
22,4 |
3,5 |
20,0 |
2,6 |
18,9 |
1,0 |
|
фосфата |
|||||||
|
|
|
|
|
|
||
4.4Морфологические изменения нервно-мышечных образований при различных режимах ЭС
Приведенные данные говорят о том, что длительная ЭС может отрицательно влиять на работоспособность организма. Подтверждается это таже данными морфологических исследований, которые показали, что длительная ЭС вызывает реактивные изменения со стороны мышечных волокон, двигательных аксонов, моторных нервных окончаний и стволов седалищных нервов как стимулированной, так и симметричной конечностей (рис. 4.2.).
Уже на пятый день эксперимента отмечается, увеличение агирофилии в большинстве моторных аксонов, появление во многих из них выраженных варикозностей, одно и двусторонних выпячиваний в претерминальных участках. Эти изменения являются еще обратимыми и свидетельствуют о высокой функциональной напряженности структур.
82

Рис. 4.1. Динамика показателей хронаксии (1,3) и реобазы (2,4) мышц при электростимуляции без введения (1,3) и с введением (3,4) АТФ.
Рис 4.2. Двуглавая мышца бедра кролика после 10(а) и 18 (б) сеансов электростимуляции:
83
а - повышена аргирофилия и наплывы нейроплазмы притерминаль-ных отделов деятельных отделов двигательных аксомов. В одной двигательной бляшке резко повышена аргирофилия яер, которые плотно размещены одно к одному.
б - фрагментация и комочкообразный распад осевоцилиндрических отростков двигательных аксонов.
Импрегнация по Бильшовскому. .МБ 16. ок.15. об 40.
Через десять суток эксперимента реактивные изменения моторных аксонов становятся более резкими: появляются массивные натеки нейроплазмы, чередующиеся с резким ее сужением. Кроме того, отдельные нервные волокна подвергаются фрагментации. Моторные нервные окончания характеризуются различным состоянием: часть не проявляет никакой реакции на раздражение, большинство находится в раздраженном состоянии, во многих двигательных бляшках происходят патологические изменения – их терминали огрубевают, резко сокращаются, многие из них фрагментируются по типу автономии.
Последний феномен трактуется, как защитная реакция со стороны периферической нервной системы, которая путем ослабления контакта между нервными и мышечными волокнами обеспечивает предохранение передачи импульсации, не отвечающей функциональному состоянию иннервируемого объекта. Об этом же свидетельствует уменьшение размеров многих двигательных бляшек, ядра которых собираются в комочки (рис. 4.2. а).
Через 18 суток эксперимента эти реакции становятся еще более отчетливыми. Наблюдается распад отдельных двигательных бляшек, а также моторных аксонов не только внутримышечных нервных стволах, но и в стволе седалищного нерва. Это свидетельствует о восходящем характере дегенеративного процесса (рис. 4.2. б). Через 10, а особенно 18 дней эксперимента на многих мышечных волокнах возникают узлы локальных сокращений (контрактуры). Плотность поперечной исчерченности в участках контрактур резко увеличивается за счет уменьшения толщины изотропных дисков, что происходит в результате глубокого вхождения ниток актина между нитками миозина. Между контрактурами мышечные волокна имеют другой вид. Здесь изотропные диски слишком широкие и сцепление ниток миозина с нитями актина незначительное. Если рассматривать такие мышечные волокна по длине, то можно заметить, что различные их участки характеризуются различным функциональным состоянием – резким сокращением, расслаблением и растяжением (рис. 4.3.).
Это может происходить вследствие локальных нарушений метаболических процессов в мышечных волокнах в связи с изменениями эндоплазматической сетки. Как правило, такие изменения являются обратимыми. Однако, очень часто в участках значительных контрактур разрушается молекулярная организация ниток актина и миозина (рис. 4.3.).
84

Рис. 4.3. Двуглавая мышца бедра кролика после 10 сеансов электростимуляции. Локальные контрактуры на мышечных волокнах. В участке крупных контрактур исчезла поперечная исчерченность и резко проявилась фибриллярность. Гематоксилин - эозин. МБ 16, ок.20, об.20.
Поперечная исчерченность полностью исчезает, наступает так называемая гомогенизация мышечного волокна. Эти изменения не обратимы. Они ведут к нарушению мышечных волокон, которые в дальнейшем фрагментируются и подвергаются восковидной дегенерации. Фрагменты таких волокон были выявлены в двуглавой и четырехглавой мышцах конечностей, после 18-и сеансов ЭС (рис. 4.4.). В поле зрения на 100 волокон 2-3 нарушения.
После прекращения ЭС в течение 5-10 дней контрактуры мышечных волокон постепенно исчезают, происходит внутриклеточная регенерация мышечных волокон, они приобретают нормальную структуру. Фрагменты мышечных волокон, что распадались, рассасываются и на их месте развивается соединительная ткань.
85
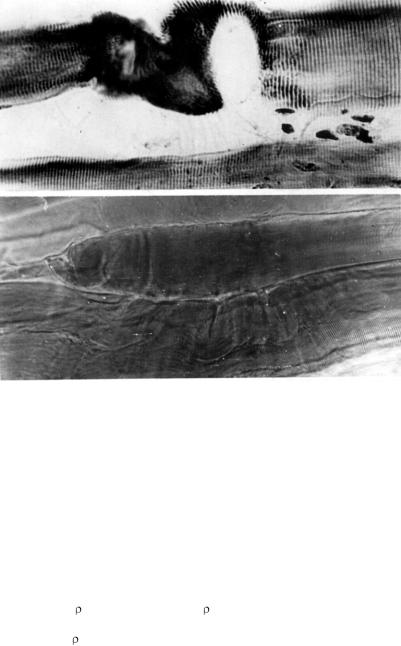
Рис. 4.4. Двуглавая мышца бедра после 18 сеансов электростиму-ляции. Дегенерация отдельных мышечных волокон. Импрегнация по Бильшовскому (вверху), гематоксилин - эозин (внизу), МБ16, ок. 20, об. 20.
Явление раздражения периферических отделов двигательных аксонов сохраняется до 15-и суток. Наблюдается прямая или коллатеральная регенерация периферических отделов моторных аксонов, которые подверглись дегенеративным изменениям.
Все сказанное дает основание считать нецелесообразным применение длительного режима ЭС (в частности 18 сеансов для животых).
Поэтому в следующей серии исследований мы уменьшили количество сеансов до 5-и.
Как видно из таблицы 4.3, содержание АТФ, креатинфосфата и активность АТЗ-азы фрагментов саркоплазматического ретикулума мышц, стимулированных на протяжении 5-и дней, достоверно не изменяется по сравнению с контрольными ( >0,5) и нормальными ( >0,5) мышцами, достоверно уменьшается только интенсивность накопления Са2+ в фрагментах саркоплазматического ретикулума ( >0,05).
86

Таблица 4.3 Накопление Са2+ и АТФ-азная активность в фрагментах саркопазматиче-
ского ретикулума, содержание АТФ и креатинфосфата в скелетных мышцах кроликов после пятидневной ЭС (М m).
|
Стимулиро- |
Симметричная к |
Нормальная |
||||
Исследуемые показатели |
ван–ная мыш- |
стимулиро–ванной |
|||||
мышца (n=9) |
|||||||
|
ца (n=5) |
мышце (n=5) |
|||||
|
|
|
|||||
Интенсивность накопления Са2+ |
0,9 |
0,1 |
1,3 |
0,1 |
1,2 |
0,1 |
|
АТФ-азная активность |
2,2 |
0,3 |
2,2 |
0,4 |
2,0 |
0,1 |
|
Содержание АТФ |
40,7 |
3,7 |
41,0 |
3,4 |
36,3 |
1,0 |
|
Содержание креатинфосфата |
26,5 |
3,6 |
25,3 |
3,4 |
18,9 |
1,0 |
|
Интенсивность накопления Са2+ |
1,2 |
0,18 |
1,28 |
0,2 |
–– |
|
|
при введении АТФ |
|
||||||
|
|
|
|
|
|
||
Пятидневная ЭС (по 10 минут в день) не вызывает заметных под свето- |
|||||||
вым микроскопом изменений мышечной ткани. |
|
|
|
||||
|
|
|
|
|
Таблица 4.4 |
||
Накопление Са2+ и АТФ-азная активность в фрагментах саркопазматиче- |
|||||||
ского ретикулума, содержание АТФ и креатинфосфата в скелетных мышцах кроликов при электростимуляции два раза по пять дней с периодами отдыха /М m/.
Исследуемые показа- |
Стимулирован- |
Симметричная к стиму- |
Нормальная |
||||
тели |
ная мышца (n=5) |
лированной мышце (n=5) |
мышца (n=9) |
||||
Интенсив–ность |
0,81 |
0,1 |
1,3 |
0,2 |
1,2 |
0,1 |
|
накопления Са2+ |
|||||||
АТФ-азная активность |
1,65 |
0,2 |
2,0 |
0,3 |
2,0 |
0,1 |
|
Содержание АТФ |
33,4 |
4,0 |
39,3 |
4,2 |
36,3 |
1,0 |
|
Содержание |
креатин- |
25,4 |
4,2 |
24,8 |
4,5 |
18,9 |
1,0 |
фос–фата |
|
||||||
|
|
|
|
|
|
|
|
Интенсив–ность |
|
|
|
|
|
|
|
накопления |
Са2+ при |
1,2 |
0,1 |
1,2 |
0,2 |
--- |
|
введении АТФ |
|
|
|
|
|
|
|
Поскольку в этой серии исследований наблюдается уменьшение АТФзависимого накопления Са2+, что характерно при наследственной мышечной дистрофии [31,32], решено было испытать режим стимуляции с включением периода отдыха, чтобы создать возможность для восстановления нарушенных после нагрузки процессов.
После 5-и сеансов ЭС животные два дня отдыхали, потом еще 5 дней стимулировались и 4 дня отдыхали. В таблице 4.4 приведены данные этих исследований.
Содержание АТФ, креатинфосфата и АТФ - азная активность фрагментов саркоплазматического ретикулума заметно не меняются ( >0,2), а интенсивность накопления Са2+ в них как и при стимуляции на протяжении 18-и и 5-и суток достоверно снижается ( <0,05). Мышцы контрольных конечностей по этим показателям не отличаются от нормы ( >0,5).
87
Таким образом, включение отдыха при десятидневной электростимуляции не способствует полной нормализации этого важного для сократительного акта процесса.
Исходя из данных, полученных в отделе биохимии мышц института биохимии НАНУ о нормализующем влиянии АТФ на протекание обменных процессов в мышцах [33-35], было решено провести исследования с введением АТФ на протяжении всего периода ЭС. Одновременно стимулировали 2 кролика, одному из которых вводили ежедневно по 1 мл 1%-го раствора натриевой соли АТФ. Применяли последние два режима стимуляции, а именно: 5 дней и 2 раза по 5 дней с периодами отдыха. Выяснилось (таблицы 4.3 и 4.4), что введение кроликам АТФ предохраняет снижение интенсивности накопления Са2+ фрагментами саркоплазматического ретикулума. Содержание АТФ и креатинфосфата остается без изменений. Величина физиологических показателей при этом уменьшается, что свидетельствует о повышени возбудимости. По данным морфологических исследований контрактуры мышечных волокон у животных, которым на протяжении всего периода ЭС вводили АТФ, возникали очень редко.
4.5Функциональные изменения нервно-мышечной системы при различных режимах электростиму-ляции
Электростимуляция не только успешно внедряется в клиническую, спортивную и космическую медицину и реабилитацию, но и успешно применяется в косметологии. Однако ряд важных вопросов, касающихся оптимизации режимов миостимуляции, остаётся нерешенным. На наш взгляд, наиболее остро стоит вопрос подбора наиболее адекватных характеристик стимулирующих сигналов и методик стимуляции. В этой связи В.Б.Ласков [36] выполнил работу, в которой стояла задача сравнить эффективность использования приборов Тонус-'1", "Тонус-2", "Саноматик" (типов SХ-3 и SХ-4) и аппарата "Миоритм". Приборы "Тонус-1", " Тонус-2" и "Саноматик" предназначенны для стимуляции мышц диадинамическими токами, аппарат "Миоритм" генерирует размыкательные фарадические импульсы с амплитудно-частотной модуляцией, причём девиация частот составляет от 20 до 110 Гц.
В качестве экспериментального материала использовались кролики в связи с имеющимися данными о схожести течения посттравматических процессов при повреждении нервно-мышечного аппарата человека и кролика. Исследование проведено на 30 кроликах.
Все животные подверглись операции, при которой каждому из них в правый седалищный нерв вживлялись биполярные раздражающие электроды специальной конструкции и подшивались контакты биполярного отводящего электрода к икроножным и перонеальным группам мышц; провода от электродов фиксировались к фасциям и выводились в область шеи у затылочной кости животного. Раны зашивались и животное в течение недели находилось на восстановлении [36].
88
Контроль за состоянием животных осуществляли с помощью стимуляционной электронейромиографии (ЭНМ). При этом на миографической системе МГ-42 ("Медикор", Венгрия) регистрировали латентные периоды вызванных супермаксимальной стимуляцией потенциалов действия мышц (М-ответов), амплитуды максимального (Амакс) и минимального (Амин) М-ответов и число функционирующих единиц (ДЕ) в изучаемых мышцах при раздражении седалищных нервов. Кроме того, определяли пороги генерации М-ответов при стимуляции седалищных нервов.
Фоновые параметры после операции определяли до 5 раз, и при получении идентичных результатов при последних трёх измерениях приступали к электростимуляции животного, Проведено 2 серии опытов.
В 1-ой серии опытов преследовалась цель установить оптимальную длительность сеанса электростимуляции. Для этого у 5 кроликов с помощью аппарата "Миоритм" стимулировали мышцы голени в течение 10 минут, и также у 5 кроликов раздражали аналогичные мышцы в течение 15 минут. Режим стимуляции : 1с – посылка раздражающих импульсов, 1с – пауза. Интенсивность стимуляции устанавливали таким образом, чтобы амплитуда возникающего мышечного сокращения составляла 90% от максимальной. Число сеансов ЭС – 5, по 1 сеансу в день. После окончания курса миостимуляции на 6-ой день контролировали показатели электронейромиограммы, проводили сопоставления с фоновыми величинами соответствующих параметров.
Опыты 2-ой серии имели своей целью установить, применение каких приборов дает наибольший физиологический эффект. Эксперимент проведён на 20 кроликах. После регистрации достоверных фоновых параметров у 10 из них проводили стимуляцию мышц голени с помощью одного канала прибора "Миоритм-07" (длительность пачки импульсов – 1с, длительность паузы – 1с). Сила раздражения обеспечивала получение мышечного сокращения до 90% от максимального.
У других 10 животных стимулировали аналогичные мышцы прибором "Саноматик" (контрольная группа). Ритм – 1с сокращение – 1с расслабление. Амплитуда вызванных мышечных сокращений достигала 90% от максимально возможной.
Длительность сеанса ЭС в опытах второй серии установлена на основании результатов эксперементов 1-ой серии и составляла 10 минут, число сеансов – 10,
Таким образом, обе группы животных во 2-ой серии опытов имели одинаковый режим ЭС, что позволяло получить сопоставимые результаты, До начала курса ЭС и после её окончания проводили ЭНМГ контроль.
Через 3 недели после окончания курса ЭС у животных экспериментальной группы повторили курс миостимуляции с помощью 2-х каналов прибора "Миоритм", раздражая попеременно икроножные мышцы и мышцы перинеальной группы. До начала курса стимуляции и после её окончания также определены ЭНМГ показатели.
89