

иммунитет, агглютинация
.pdf1. Понятие «иммунитет».
Иммунитет – это способность многоклеточных организмов поддерживать постоянство своего макромолекулярного состава путем удаления чужеродных молекул, что обеспечивает устойчивость к инфекционным агентам и резистентность к опухолям.
2. Сравнительная х-ка врожденного и адаптивного иммунитета (цели, осн. этапы).
3. «Образы патогенности», их характеристика, примеры; паттерн-распознающие
рецепторы, их локализация, примеры.
Образы патогенности, или патогенассоциированные молекулярные паттерны — группы молекул, как правило, отсутствующие в организме-хозяине, но характерные для патогенов (вирусов, бактерий, грибов, простейших, паразитов). При этом PAMP связаны с патогенностью и могут рассматриваться как знаки опасности, наиболее
универсальный сигнал о проникновении в организм не просто чужеродного, но биологически агрессивного агента.
Рецепторы для PAMP ( паттернраспознающие рецепторы) позволяют распознавать все возможные типы патогенов и представлены у всех многоклеточных, включая не только животных, но и растения. Узнавание PAMP — основа распознавания во врожденном иммунитете; в определенной степени PAMP способны узнавать и клетки адаптивного иммунитета.
Примерами распознаваемых образов могут служить пептидогликан и липополисахари-ды клеточной стенки, структуры на основе тейхоевых кислот, двуцепочечные РНК (харак-терны только для вирусов), N-формил-метионин (структура, с которой начинается боль-шинство бактериальных белков) и др. На фагоцитах имеются рецепторы, распознающие такие образы.
4. Строение иммунной системы: первичные и вторичные органы, их роль в формировании и созревании иммунных клеток, в развитии иммунных реакций.
Иммунная система состоит из центрального и периферического отделов. Центральный отдел содержит органы, в которых происходят дифференцировка и созревание лимфоцитов: костный мозг и тимус. Костный мозг служит местом сосредоточения эффекторных клеток адаптивного иммунитета (например, плазмоцитов). Таким образом, только тимус является чисто центральным лимфоидным органом. Основная его функция — обеспечение развития Т-лимфоцитов. Название центральных лимфоидных органов определило обозначение основных типов лимфоцитов: Т (тимусзависимые) и В (бурсазависимые). Название третьего типа лимфоидных клеток
— NK-клеток — происходит от выполняемой ими функции (естественные киллеры — natural killers). Периферический отдел иммунной системы образован периферическими лимфоидными органами: неинкапсулироваными лимфоидными структурами, связанными со слизистыми оболочками, диффузно распределенными лимфоидными и миелоидными клетками и инкапсулированными лимфоидными органами. Все лимфоидные органы организованы сходным образом. В них выделяют зоны сосредоточения Т-лимфоцитов, В-лимфоцитов, а также сегменты, в которых В- и Т-клетки соседствуют друг с другом. Комплекс этих образований вместе с диффузно распределенными лимфоцитами, пейеровыми бляшками и региональными лимфатическими узлами формирует лимфоидную ткань, связанную со слизистыми оболочками. Стромальные клетки лимфоидных органов и MALT способны привлекать клетки соответствующих типов и поддерживать их жизнеспособность. Завершив развитие в костном мозгу, миелоидные клетки поступают в кровь и некоторое время циркулируют в кровотоке. Из кровотока они мигрируют в ткани, в которых живут от нескольких суток до месяцев или лет. Кроме такого конститутивного пути миграции, существует экстренная миграция клеток из кровотока в места контакта с патогеном.
5. Клетки, реализующие врожденный иммунитет (миелоидные и лимфоидные).
Миелоидные клетки, участвующие в иммунных процессах, представлены моноцитами, нейтрофильными, эозинофильными и базофильными гранулоцитами. А также тучные клетки и 2 типа тканевых клеток, образующихся из моноцитов, — макрофаги и дендритные клетки. Последние играют роль посредника между врожденным и адаптивным иммунитетом. Миелоидные клетки экспрессируют комплекс рецепторов, распознающих PAMP.
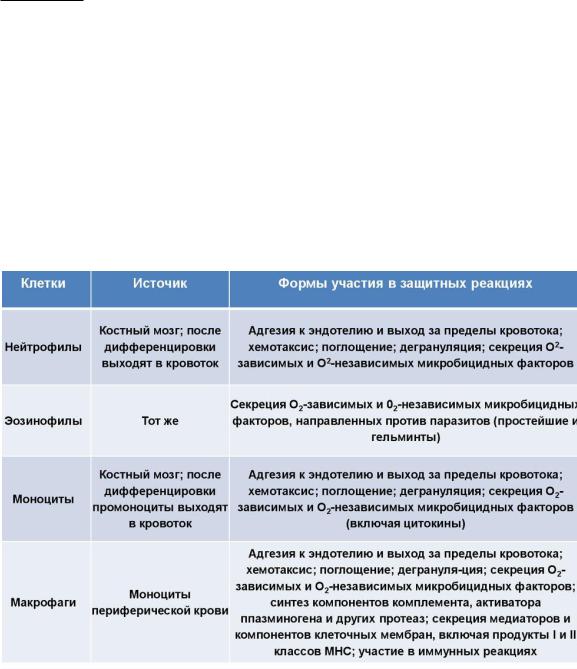
Выделяют 3 основных типа лимфоцитов — Т-, В- и естественные киллеры. NK-клетки относят к клеткам врожденного иммунитета. Они распознают стрессорные молекулы, отличные от распознаваемых миелоидными клетками.
Иммуноглобулиновый рецептор В-клеток дает им возможность распознавать антиген как в свободной, так и в связанной с мембранами формах.
Рецептор Т-клеток распознает только фрагменты антигена, связанные с молекулами
MHC.
Популяции Т- и В-лимфоцитов участвуют в иммунных реакциях клонального типа, при которых в ответ вовлекаются только клетки клонов, экспрессирующих рецепторы нужной специфичности (в отличие от естественных киллеров, не отличающихся друг от друга по специфичности).
К естественным относят 3 субпопуляции В-клеток.
В1-клетки локализованы в серозных полостях и барьерных тканях, несут рецептор с низкой специфичностью к антигену, спонтанно вырабатывают низкоаффинные антитела преимущественно IgM-изотипа, в том числе к аутоантигенам.
В- клетки маргинальной зоны — клетки, сходные с В1, но локализующиеся в маргинальной зоне селезенки.
В2-клетки локализованы в селезенке и лимфатических узлах (в том числе в фолликулах), костном мозгу, лимфоидных тканях кишечника; эти клетки отвечают за образование высокоспецифичных и высокоаффинных антител разных изотипов.
Число естественных субпопуляций Т-лимфоцитов значительно больше. Среди «классических» αβТ-клеток выделяют субпопуляции CD4+ и CD8+ Т-клеток, отличающиеся двумя основными особенностями — распознаванием антигенных пептидов в составе разных молекул MHC и функцией: после стимуляции антигеном CD4+ выступают в качестве хелперов, а CD8+ — цитотоксических Т-лимфоцитов. CD8+ Т-клетки в процессе иммунного ответа функционируют как единая субпопуляция. CD4+ клетки образуют несколько «адаптивных» субпопуляций. Существует естественная субпопуляция CD4+ Т-клеток — естественные регуляторные Т-клетки . Их функция состоит в контроле активности аутоспецифических клонов Т-лимфоцитов, не удаленных в процессе отрицательной селекции, мигрировавших в периферический отдел иммунной системы и создающих опасность аутоагрессии.
6. Фагоцитирующие клетки, их характеристика.

7. Характеристика макрофагов и дендритных клеток как фагоцитирующих и антигенпрезентирующих клеток.
8. Фагоцитоз: характеристика стадий, механизмы переваривания. Завершенный и незавершенный фагоцитоз, примеры.
Фагоцитоз - сложноорганизованный в пространстве и времени процесс поглощения крупных комплексов, или конгламератов клетки.
Этапы:
1.Хемотаксис. Лейкоциты двигаются к источнику хемоатракантов. Источник хемоатрактантов микроорганизм.
2.Адгезия лейкоцитов к микроорганизмам. Этот процесс сопровождается присоединением к рецепторам лейкоцитов различных антигенов. Наблюдается «кэппинг эффект»: перераспределение рецепторов к тому полюсу клетки, где наблюдается большая концентрация антигена.
3.Инвагинация плазматических мембран с образованием фагосомы. В этом процессе участвуют белки цитоскелета.
4.Слияние фагосомы с лизосомой => фаголизосома. У нейтрофилов наблюдается слияние фагосомы с гранулами.
5.Деструкция поглощенного антигена, до определенных продуктов.
6.Элиминациявыделение продуктов деструкции из клетки.
Завершенный фагоцитоз – задачей данного вида фагоцитоза является деструкция первично обработанного чужеродного материала. Характерен для всех лейкоцитов, но чаще характерен для нейтрофилов и макрофагов Незавершенный фагоцитоз - микробные клетки остаются жизнеспособными.
Механизмы: микобактерии туберкулеза и токсоплазмы препятствуют слиянию фагосом с лизосомами; гонококки могут быть к действию лизосомальным ферментам, риккетсии и хламидии могут долго персистировать в цитоплазме вне фаголизосомы.
9. Гуморальные факторы врожденного иммунитета (система комплемента, цитокины, антимикробны пептиды, белки острой фазы воспаления), их характеристика

Гуморальные факторы врожденного иммунитета – это белки, присутствующие в сыворотке крови, секретах слизистых оболочек и некоторых других жидкостях организма.
Система комплемента: система сывороточных и мембраносвязывающих белков с каскадным ферментативным действием. Биологическая роль: Лизис атакуемой клетки при помощи мембранатакующего комплекса, Опсонизация, обусловленная фиксацией C3b-компонента комплемента, Вовлечение в воспалительную реакцию.
Белки острой фазы продуцируются гепатоцитами, моноцитами, макрофагами и фибробластами. Синтез повышается при инфекционном процессе. Так, уровень маннозосвязывающего лектина возрастает при инфекции 2-3 раза, а концентрация С-реактивного белка может в 100 раз.
Бактерицидные пептиды – группа катионных белков, способных поражать многие вирусы, бактерии, грибы и простейшие. Осуществляют «мгновенный иммунитет». Синтезируются нейтрофилами и эпителиальными клетками слизистых оболочек и выделяются в межклеточное пространство.
Цитокины – класс полипептидных медиаторов межклеточного взаимодействия, участвующих в развитии иммунных реакций при внедрении патогенов, а также в регуляции физиологических функций (воспаление, гемопоэз, ангиогенез, и др.).
10. Комплемент, пути его активации.
Существует три пути активации системы комплемента: классический, альтернативный и лектиновый.
Классический путь активации системы комплемента предусматривает обязательное участие антител - IgG1 или IgM. Антиген предварительно связывается со специфическим ему антителом, формируется иммунный комплекс, который связывается с С1-компонентом комплемента, запуская дальнейший каскад реакции.
Альтернативный путь запускается при спонтанном расщеплении С3 или под влиянием С3-конвертаз, а также сывороточных либо микробных протеаз.
Лектиновый путь активирует систему комплемента через лектин, связывающий маннозу. К системе врожденного иммунитета можно отнести только альтернативный и лектиновый пути активации системы комплемента, так как не требуется участия АТ.
11. Группы цитокинов, их функции
На основе биологических эффектов выделяют несколько групп цитокинов: Интерлейкины (IL1 - IL35) – регуляторные молекулы иммунной системы, обеспечивают внутриклеточные медиаторные взаимодействия, связь иммунной системы с другими системами организма; Интерфероны – противовирусные агенты с иммунорегуляторным действием;
Факторы некроза опухоли – цитокины с цитотоксическим и регуляторным действием; Хемокины (IL-8, СХ3С) – регуляторы хемотаксиса различных типов клеток;
Факторы роста – регуляторы роста, дифференцировки и функциональной активности клеток различной тканевой принадлежности (напр.,фактор роста фибробластов) и трансформирующий фактор роста (ТФР).
12. Классификация интерферонов, их характеристика, клетки-продуценты интерферонов. Роль интерферонов во врожденном и адаптивном иммунитете.
Интерфероны (IFN, ИФН) -ряд биологически активных белков или гликопротеидов со сходными свойствами, синтезируемых клетками организма в процессе защитной реакции в ответ на вторжение чужеродных агентов. Благодаря интерферонам клетки становятся невосприимчивыми по отношению к вирусу.
IFN подразделяют на три типа:
Тип I - известный как вирусный интерферон, включает IFN-a (синтезируется моноцитами и В-лимфоцитами), IFN-b (синтезируется фибробластами и эпителиальными клетками, макрофагами)
Тип II - иммунный, включает IFN-y (синтезируется Т-лимфоцитами и NK-клетками); Тип III - важен в некоторых видах вирусных инфекций.
Первому типу, главным образом, присущи антивирусный и антипролиферативный эффекты, в меньшей степени - иммуномодулирующий. Они вырабатываются непосредственно после встречи с патогеном, действие направлено на локализацию возбудителя и предотвращение его распространения в организме. Эти интерфероны осуществляют раннюю и неспецифическую защиту организма от инфекционного агента.
Главное действие интерферонов второго типа - участие в реакциях иммунитета. Начинает вырабатываться на следующих этапах инф. процесса сенсибилизированными Т-лимфоцитами, участвует в каскаде специфического иммунного ответа.
Синтез интерферонов II типа индуцируется митогенными или антигенными стимулами. Большинство типов вирусоинфицированных клеток способно синтезировать IFN-a/b в клеточной культуре. В противоположность этому IFN-y синтезируется только некоторыми клетками иммунной системы, включая естественные киллерные (NK) клетки, CD4 Т-клетки и CDS цитотоксические супрессорные клетки.
13. Антигены бактерий. Структурные (клеточные) антигены: О, Н, К, Vi. Экстрацеллюлярные антигены: экзотоксины, ферменты агрессии.
В структуре бактериальной клетки разли чают жгутиковые, соматические, капсульные и некоторые другие антигены. Жгутиковые, или Н-антигены, локализуются в локомоторном аппарате бактерий — их жгутиках. Они представляют собой эпитопы сократительного белка флагеллина. При нагревании флагеллин денатурирует, и Н-антиген теряет свою специфичность.
Соматический, или О-антиген, связан с клеточной стенкой бактерий. Его основу составляют ЛПС. О-антиген проявляет термостабильные свойства — он не разрушается при длительном кипячении. Однако соматический антиген подвержен действию альдегидов и спиртов, которые нарушают его структуру.
Капсульные, или К-антигены, располагаются на поверхности клеточной стенки. Встречаются у бактерий, образующих капсулу. К-антигены состоят из кислых полисахаридов (уроновые кислоты).
На поверхности возбудителя брюшного тифа и других энтеробактерий, которые облада ют высокой вирулентностью, можно обнаружить антигена вирулентности, или Vi-антигена. Обнаружение этого антигена или специфичных к нему антител име ет большое диагностическое значение.
14. Антитела: структура молекулы иммуноглобулина (константные и вариабельные участки, домены, активные центры, их функции).
Структура молекулы антитела (иммуноглобулина) включает:
1.Вариабельную область (V-область), которая у разных антител может варьироваться. V-области L- и Н-цепей образуют Аг-связывающий центр (активный центр AT, паратоп), или Fab-фрагмент.
2.Константную область (С-область), которая называется Fc-фрагмент.

3.Шарнирную область в месте соединения Fab- и Fc-фрагментов, которая позволяет Аг-связывающим фрагментам разворачиваться для более тесного контакта с Аг.
15. Классы иммуноглобулинов, их характеристика. Изо-, алло-, идиотипы иммуноглобулинов. Механизм взаимодействия антигена с антителом. Полные, неполные антитела. Нормальные антитела. Аутоантитела.
IgA - основной изотип слизистых секретов.
IgM - первыми появляются на самых ранних стадиях гуморального иммунного ответа на инфекцию (до 7-14 дней).
IgG - вторично вырабатываются на инфекцию (через 7- 14 дней после инфицирования). Более стойкие. Способны преодолевать плацентарный барьер. Доминируют по своему количеству в сосудистом русле.
IgE - прикреплены к поверхности тучных клеток.
IgD - экспрессируется на поверхностях Влимфоцитов,но не секретируется им.
Взаимодействие антиген-антитело основано на принципе взаимного узнавания, обусловленного их конформационной комплементарностью. На основе знаний полученных на предыдущих занятиях – активный центр антитела строго комплементарен антигенной детерминанте, что обеспечивает их связывание.
Полные антитела - это антитела, которые имеют 2 и более активных центра. После их соединения с антигеном образуется видимый осадок (агглютинат, преципитат). Неполные антитела - это антитела, которые имеют один активный центр. Они способны связываться с антигенами, но это не сопровождается видимыми изменениями.
Нормальные антитела - это антитела, которые постоянно имеются у человека и животных без попадания в организм антигена (без иммунизации). К ним относятся, например, антитела плазмы крови (агглютинины), которые определяют деление крови человека на 4 группы.
16. Адаптивный иммунитет: гуморальный иммунный ответ (клеткиучастники, их взаимодействие), динамика образования антител при первичном и вторичном иммунном ответе; клеточный иммунный ответ (варианты развития, механизм).
Гуморальный иммунный ответ защищает преимущественно против внеклеточно паразитирующих микроорганизмов, которые доступны действию специфических антител. Продуцентами антител являются плазматические клетки. В-лимфоцит получает сигнал активации при соединении его антиген-распознающего рецептора с антигеном. Для активной
пролиферации и дифференцировки в плазматические клетки потомки В-лимфоцита получают дополнительные сигналы активации — продуцируемые Th2 цитокины: интерлейкины-4, -5, -6, -10, -13. Те же цитокины усиливают продукцию и секрецию специфических для данного антигена антител зрелыми плазматическими клетками.
На скорость образования АТ влияет: доза Аг, частота Аг-стимуляции и состояние иммунной системы индивида. Если организм впервые встречается с Аг, то развивается первичный иммунный ответ, а при повторном контакте — вторичный ответ.
Первичный иммунный ответ. Появлению АТ предшествует латентный период продолжительностью 3~5 сут. В это время происходит распознавание Аг и образование клонов плазматических клеток. Затем наступает логарифмическая фаза, соответствующая поступлению АТ в кровь; её продолжительность — 7-15 сут. Постепенно титры АТ достигают пика и наступает стационарная фаза, около 15-30 сут. Её сменяет фаза снижения титров AT, длящаяся 1-6 мес. В динамике антителообразования титры высокоаффинных AT постепенно нарастают: после иммунизации аффинность AT к Аг постоянно увеличивается. Первоначально образуются IgM, постепенно начинает преобладать синтез IgG. Особенности первичного ответа — низкая скорость антитело -образования и появление сравнительно невысоких титров
AT.
Вторичный иммунный ответ После антигенной стимуляции часть В- и Т-лимфоцитов циркулирует в виде клеток памяти. Особенности вторичного иммунного ответа — высокая скорость антителообразования, появление максимальных титров АТ и длительное их циркулирование. Основные характеристики вторичного имунного ответа: • образование АТ индуцируется значительно меньшими дозами Аг; • индуктивная фаза сокращается до 5-6 ч; • среди АТ доминируют IgG с большой аффинностью, пик их образования наступает раньше (3-5 сут); • АТ образуются в более высоких титрах и циркулируют в организме длительное время. Клеточный иммунный ответ направлен против внутриклеточно паразитирующих микроорганизмов, основная защитная роль в нем принадлежит активированным макрофагам и цитотоксическим лимфоцитам (CD8+ CTL). Макрофаги, инфицированные микроорганизмами, получают от Thl в качестве сигналов активации цитокины: гамма-интерферон и туморнекротизирующий фактор (ТНФ), которые действуют через свои рецепторы, вызывая усиленную продукцию макрофагами супероксидных и нитроксидных радикалов, убивающих внутриклеточные паразиты. Цитотоксические CD8+ CTL способны убивать зараженные вирусами клетки при непосредственном контакте с ними. В месте контакта из CTL в мембрану клетки-мишени проникают порообразующие белки — перфорины, формирующие в мембране микроканалы, через которые в клетку-мишень проникают ферменты — фрагментины, вызывающие разрушение ядра клетки и ее гибель.
17. Диагностические реакции с участием антигенов и антител, их практическое использование.
Реакции с использованием меченых AT и Аг составляют основу методов экспресс-диагностики инф. заболеваний, т.к. выявляют минимальное содержание Аг и AT в исследуемых образцах. В качестве меток могут быть использованы различные ферменты, красители и изотопы.
В основе метода радиоиммунного анализа — маркирование радионуклидом Аг или AT, вступающих в реакцию. Образующиеся иммунные комплексы выделяют из системы и определяют их радиоактивность на счётчиках импульсов. Наибольшее распространение получил радиоиммунный анализ на твёрдой фазе (твердофазный РИА) с использованием меченых Аг или AT, сорбированных в лунках полистироловых панелей. РИА применяют для выявления микробных Аг, различных гормонов, ферментов и т.д.
18. Реакции агглютинации (прямая, реакция непрямой гемагглютинации, реакция агглютинации латекса, реакция коагглютинации, реакция Кумбса), механизм.
Реакция агглютинации позволяет выявить Аг, локализованные на поверхности сравнительно крупных частиц.
Прямая реакция: к отмытым эритроцитам крови больного добавляют антииммуноглобулиновую сыворотку. Если на эритроцитах есть неполные антитела (иммуноглобулины), что наблюдается при гемолитической анемии, резус-конфликте (эритроциты плода), то они агглютинируются.
Реакция непрямой гемагглютинации основана на использовании эритроцитов с адсорбированными на их поверхности антигенами или антителами, взаимодействие которых с соответствующими антителами или антигенами больных вызывает склеивание и выпадение эритроцитов на дно пробирки или ячейки в виде фестончатого осадка (зонтика).
Применяют для диагностики инфекционных болезней, определения гонадотропного гормона в моче при установлении беременности, для выявления повышенной чувствительности к лекарственным препаратам, гормонам и в некоторых других случаях.
Механизм. Реакция непрямой гемагглютинации (РНГА) отличается значительно более высокой чувствительностью и специфичностью , чем реакция агглютинации. В лунках пластмассовых пластин готовят последовательные разведения исследуемого материала. Затем в каждую лунку вносят одинаковый объем 3 % суспензии нагруженных антителами эритроцитов. При необходимости реакцию ставят параллельно в нескольких рядах лунок с эритроцитами, нагруженными антителами разной групповой специфичности.
Через 2 ч инкубации при 37 °С учитывают результаты, оценивая внешний вид осадка эритроцитов (без встряхивания): при отрицательной реакции появляется осадок в виде компактного.диска или кольца (пуговки) на дне лунки, при положительной реакции — характерный кружевной осадок эритроцитов, тонкая пленка с неровными краями.
Метод латекс-агглютинации. В последние годы ассортимент диагностикумов из убитых микробов дополнили так называемые суспензионные Аг и AT, сорбированные на различных инертных корпускулярных носителях. Метод латекс-агглютинации, основанный на взаимодействии Аг (или AT) в образце с частицами латекса, нагруженными специфическими моноклональными AT (или Аг), что ведёт к образованию видимых агрегатов.
Реакция коагглютинации основана на способности белка А золотистого стафилококка неспецифически связывать Fc-фрагменты Ig. В подобных ситуациях стафилококки играют роль частиц с сорбированными на их поверхности AT. При внесении в систему соответствующих Аг с ними взаимодействуют Fab-фрагменты AT и образуются агрегаты из стафилококков, диагностических AT и исследуемых микробных Аг
Непрямая реакция Кумбса применяют у больных при внутрисосудистом гемолизе. У некоторых таких больных обнаруживают антирезусные антитела, которые являются неполными, одновалентными . Наличие таких неполных антител определяют в непрямой реакции Кумбса.
Механизм. Сложность выявления неполных антител связана с тем, что эти антитела, связываюсь с АГ, не образуют структуру решетки и реакция не выявляется. Для выявления неполных антител, например к резус-антигену эритроцитов в сыворотке крови беременной 1) к двукратным разведениям испытуемой сыворотки добавляют эритроциты, содержащие резус-АГ и выдерживают при 37 °С в течение часа; 2) к отмытым после первого этапа эритроцитам добавляют кроличью античеловеческую анти-глобулиновую сыворотку. После инкубации в течение 30 мин при 37 °С результаты оценивают по наличию гемагглютинации.
19. Реакция преципитации, механизм реакции, отличия от реакции агглютинации. Постановка реакции термокольцепреципитации и реакции преципитации в геле, практическое значение.
Реакция преципитации – это формирование и осаждение комплекса растворимого молекулярного антигена с антителами в виде помутнения, называемого преципитатом. Он образуется при смешивании антигенов и антител в эквивалентных количествах; избыток одного из них снижает уровень образования иммунного комплекса.
Реакции преципитации, которые ставят в жидких средах, проявляются в виде мути, в плотных средах (в гелях, питательных средах) - полосы преципитации.
20. Диагностикумы и их получение. Постановка ориентировочной и развернутой реакций агглютинации, учет, оценка результатов.
21. Получение, титрование и использование диагностических агглютинирующих и преципитирующих сывороток.