

Глава 9
МЕХАНИКА СЕРДЕЧНОЙ МЫЩЦЫ
9.1. Модель хилла и активное состояние миокардиальных волокон
В. С. Салъманович
Основополагающие идеи мышечной механики наиболее четко сформулировал Хилл [649 ] применительно к скелетной мышце позвоночных. Согласно концепции Хилла, мышца функционирует как активный сократительный элемент, последовательно соединенный с недемпфированным пассивным упругим элементом. Постулировано, что в покое сократительный элемент свободно растяжим и поэтому вместе с последовательным упругим элементом не участвует в создании напряжения покоя. Под действием стимула сократительный элемент переходит в активное состояние. В зависимости от того, свободны или фиксированы концы исследуемого препарата, активное состояние мышцы проявляется соответственно либо как ее укорочение, либо в виде развиваемого напряжения [268].
Соотношение между скоростью укорочения v сократительного элемента мышцы и нагрузкой р в условиях тетануса Хилл выразил уравнением гиперболы
(9.1)
υ = b(p0 — p)/ p + a
где р0 — максимальное изометрическое напряжение, которое при данной длине может развивать мышца, а и b — эмпирические коэффициенты, которые не зависят от p и имеют размерность соответственно силы и скорости. Если уравнение справедливо для всего интервала скоростей, то максимальная скорость укорочения υ0 достигается при нулевой нагрузке и равна
υ0 = bp0/a (9.2)
170
Максимальное изометрическое напряжение р0 и коэффициенты a, b находятся из уравнения Хилла по данным трех независимых измерений при различных значениях р.
Попытки применить концепцию Хилла к исследованию динамики: сердечного сокращения встретили ряд затруднений, порожденных; сложностью архитектоники сердечной мышцы и некоторыми ее функциональными особенностями. К ним прежде всего следует отнести неспособность миокарда развивать тетаническое сокращение, что препятствует проведению исследования в стабильном режимеактивности, высокое напряжение покоя и сильную зависимость механических показателей от степени исходного растяжения сократительной системы. Однако разработка метода предварительной и последующей нагрузки и метода быстрого освобождения применительно к миокарду (см. с. 172 и 183) позволили отчасти обойти эти трудности и получить данные, описывающие свойства сердца
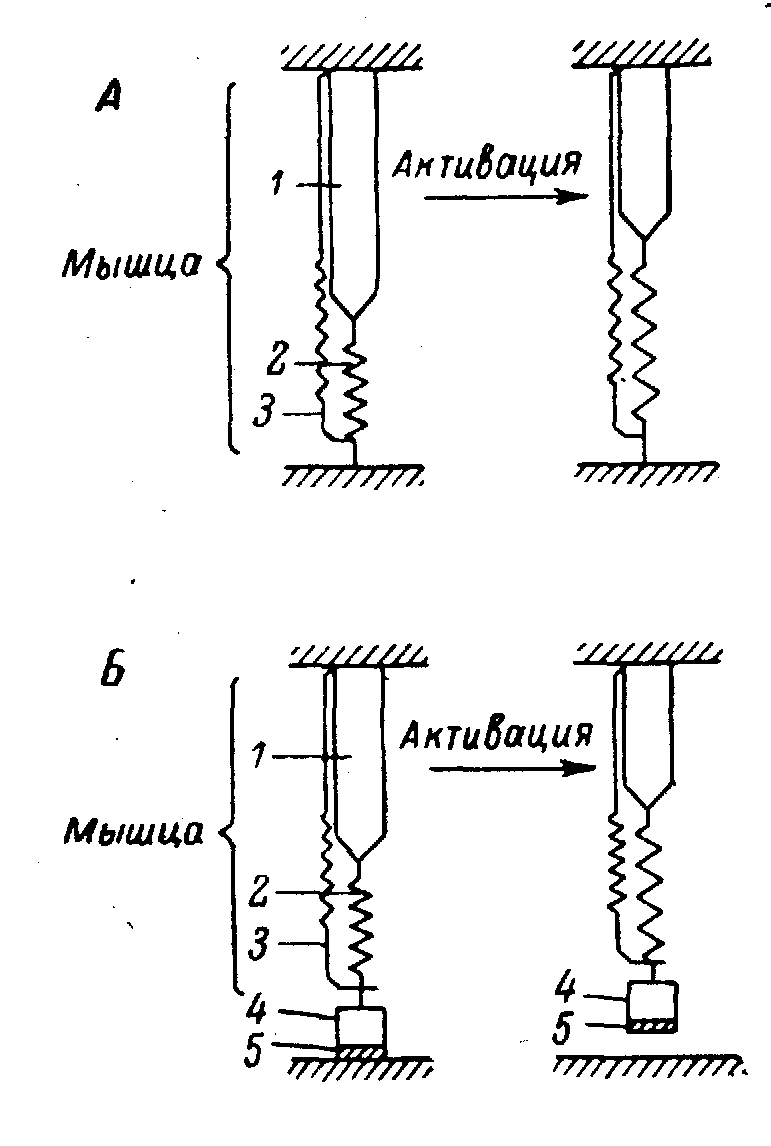
Рис. 9.1. Трехкомпонентная модель мышцы по Хиллу.
А — изометрическое, Б — изотоническое сокращение, зарегистрированное методом последующей нагрузки. Слева — положение в покое, справа — при сокращении. 1 — сократительный элемент, 2 — эластический элемент, последовательно соединенный с сократительным элементом, 3 — эластический элемент, параллельно соединенный с сократительным элементом, 4 — последовательная нагрузка, 5 — предельная нагрузка.
в тех же терминах, которые используются для скелетной мышцы.
На основе этих данных для сердечной мышцы предложена механическая модель, обычно называемая трехкомпонентной моделью Хилла (рис. 9.1). Первые два компонента ее такие же, как в двукомпонентной модели: сократительный элемент и последовательный упругий элемент. Третий же компонент представляет собой параллельный упругий элемент, который определяет напряжение покоя, или диастолическое напряжение сердечной мышцы и косвенно влияет на деятельность ее сократительного элемента и мышцы в целом [571 ]. Ниже описываются функциональные характеристики элементов механической модели миокардиального волокна и обсуждается применимость концепции Хилла к деятельности интактного сердца [364 ].
Активное состояние сократительного элемента волокон миокарда характеризуется механической энергией, которая производится в результате химических процессов в этом способном генерировать силу и укорачиваться элементе [951, 952]. Химические или
171
энергетические характеристики активного состояния нельзя измерить прямо, но можно определить их механические эквиваленты. Поэтому способ реального измерения интенсивности активного состояния и единицы, в которых она выражается, зависят от условий проведения опытов.
Для изучения изменений во времени активного состояния сердечной, как и скелетной, мышцы используется метод быстрого освобождения. Он состоит в следующем. Мышца сокращается в изо-
0 200 400 600 800 1000 1200мс
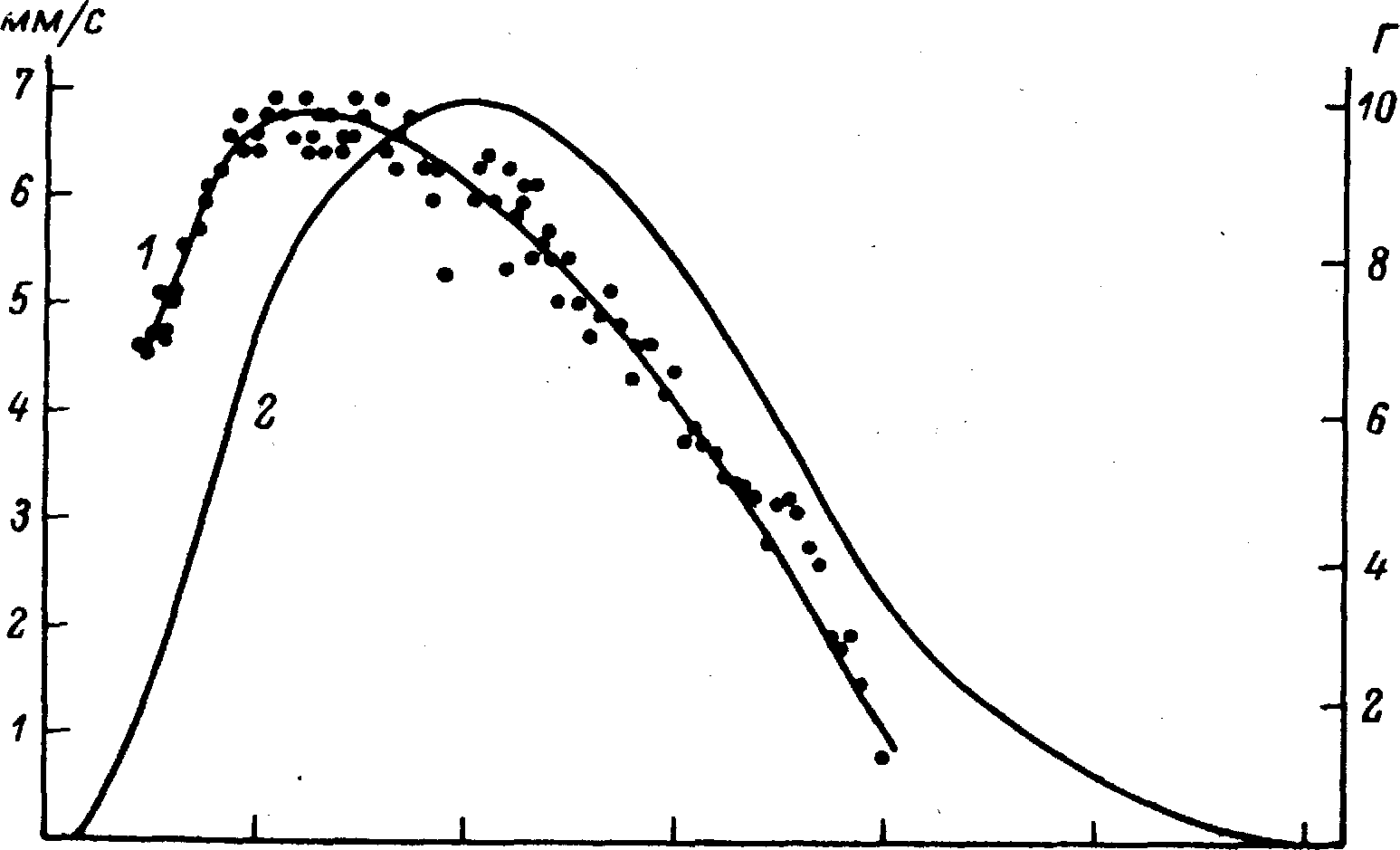
Рис. 9.2. Течение активного состояния и изометрического напряжения как функция времени после стимуляции мышцы. (По [1070]).
По оси абсцисс — время после стимуляции; по осям ординат: слева — скорость укорочения (i), справа — напряжение (2). Латентный период от момента стимуляции до первого напряжения равен 35 мс. Максимальное напряжение развивается приблизительно через 400 мс. Активное состояние определяли в каждой точке по мгновенной скорости укорочения после быстрого освобождения мышцы. 95% максимальной интенсивности активного состояния достигается после 180 мс и поддерживается около 170 мс. После 350 мс активное состояние падает ниже 95%-ного уровня.
метрическом режиме. При этом возникает укорочение сократительного элемента, приводящее к растягиванию последовательного элемента, хотя длина мышцы в целом остается неизменной. Затем один конец мышцы быстро освобождается, и она начинает сокращаться в изотоническом режиме под нагрузкой, которая меньше усилия, развитого к концу изометрической фазы. Переход от одного режима к другому вызывает очень быстрое небольшое укорочение мышцы, воспроизводящее укорочение ранее растянутого последовательного упругого элемента. Далее мышца продолжает укорачиваться за счет сократительного элемента, причем скорость этого укорочения зависит уже только от интенсивности активного состояния в момент быстрого освобождения мышцы и является, таким образом, его мерой.
Этим методом были установлены характерные черты изменения активного состояния миокарда во времени, оцениваемого по ско-
рости укорочения сократительного элемента [370, 490, 1070]. Возникающее в результате действия стимула активное состояние миокарда начинает развиваться относительно медленно и достигает максимума только через 0.3—0.5 периода его общей длительности (рис. 9.2). Пик интенсивности активного состояния не совпадает с пиком изометрического напряжения, опережая его на 150 мс. Активное состояние миокарда не имеет периода стабилизации (плато). Практически сразу по достижении максимума кривая активного состояния миокарда переходит от подъема к спаду, хотя достаточно высокая интенсивность активного состояния (в пределах 95% максимальной интенсивности) поддерживается относительно долго от 90 до 220 мс. Этот период высокой интенсивности
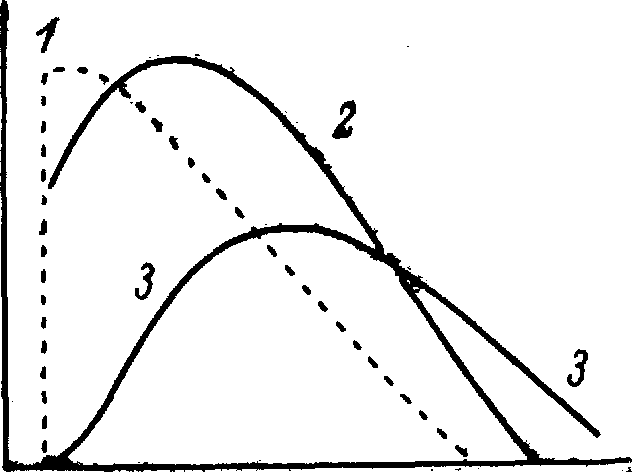
Рис. 9.3. Течение активного состояния во времени в сердечной и скелетной мышцах. (По [1070]).
По оси абсцисс — время; по оси ординат — напряжение. В скелетной мышце (1) начало актив-% ного состояния развивается очень быстро по сравнению с началом максимального активного состояния в сердечной мышце (г); з - напряжение в сердечной мышце.
приходится на вторую половину восходящей фазы изометрического сокращения (рис. 9.2).
Такое распределение во времени активного состояния миокарда значительно отличается от временного распределения его в скелетной мышце (рис. 9.3). Из рис. 9.3 следует, что активное состояние скелетной мышцы развивается очень быстро и за весьма короткий период достигает максимальной интенсивности. В отличие от миокарда в скелетной мышце максимальная интенсивность сохраняется какое-то время постоянной, формируя отчетливое плато. Другое различие в характере активности сердечной и скелетной мышц (не показанное на графике) состоит в том, что максимальная интенсивность активного состояния миокарда, определенная при данной длине его волокон, вариабельна и меняется в широких пределах, например под влиянием изменения частоты сердечных сокращений [488]. Напротив, максимальная интенсивность активного состояния скелетной мышцы относительно постоянна.
Разная форма кривых изменения активного состояния в сердечной и скелетной мышцах указывает, что либо в сердечной мышце ассоциация актина с миозином происходит более медленно, либо скорость освобождения и связывания активатора сокращения (кальция) в обоих типах мышц различна [370, 490, 724, 1065 ]. В последнем случае медленное по сравнению со скелетной мышцей достижение максимальной интенсивности активного состояния в сердечной мышце и отсутствие у него плато должны рассматриваться как показатель того, что концентрация активатора сокращения —
172
173
кальция — в миокардиальных клетках создается относительно медленно, и за период активности не достигается насыщение им сократительной системы [490].
Характер изменения во времени и интенсивность активного состояния миокарда существенно зависят от исходной длины волокна (см. гл. 18.1). Эдман и Нильсон [490] установили, что влияние оказывают также изменения длины мышцы, происходящие во время самого сокращения, хотя в начале сокращения это влияние мало выражено. В опытах авторов укорочение мышцы на 5—8% приводило к падению почти в 2 раза максимальной интенсивности активного состояния и укорочению его длительности. В связи с этим было высказано предположение, что скорость освобождения и связывания кальция в динамике сокращения миокардиальной клетки контролируется ее длиной.