

24 Influenza 2006
tients with underlying pulmonary or cardiopulmonary diseases. The very young and the elderly usually have the highest risk of developing serious complications; however, during pandemics, there is a mortality shift towards younger age groups (Simonson 1998).
In humans, replication of influenza subtypes seems to be limited to the respiratory epithelial cells. Once the virus enters a cell, it causes complex cytopathic effects, predominantly in the columnar epithelial cells, by shutting down the synthesis of host proteins. The loss of critical host cell proteins leads to cell death by necrosis (Yuen 2005). There are numerous individual factors associated with protection against or increasing the risk of a fatal outcome caused by a given influenza strain (Behrens and Stoll 2006), and genetic factors that affect host susceptibility are likely to play a role. Specific immunity against certain viral epitopes or some degree of cross-immunity may explain why people > 65 years were less affected by the 1918 pandemic. It is unknown whether similar mechanisms play a role in the curious age distribution of cases in the current outbreak of avian H5N1 influenza (ProMED 20060211.0463).
The unusual severity of H5N1 infection in humans was initially ascribed to multiple basic amino acids adjacent to the cleavage site, a feature characteristic of highly pathogenic avian influenza A viruses (Subbarao 1998). The presence of these basic amino acids renders the protein susceptible to proteases from many different types of tissues and allows extrapulmonary dissemination due to broadened tissue tropism (Yuen 2005). Another explanation may be that interferons are pivotal in preventing viral spread outside the respiratory tract and that H5N1 interferes with this innate defence against viral infection. It has been shown that the non-structural (NS) gene of highly pathogenic H5N1 viruses confers resistance to the antiviral effects of interferons and tumour necrosis factor alpha (Seo 2002). H5N1 viruses seem to induce higher gene transcription of pro-inflammatory cytokines than do H3N2 or H1N1 viruses, and are potent inducers of pro-inflammatory cytokines in macrophages, the most notable being TNF alpha (Cheung 2002). These mechanisms might ultimately lead to a cytokine storm and death (Peiris 2004).
In interpandemic influenza epidemics, recovery from interpandemic influenza is usually uneventful. In severe cases of human H5N1 influenza, however, mortality has so far been considerable (WHO 20060213). Dyspnoea, ARDS and multi-organ failure has been a dominant clinical feature in fatal cases (Hoffmann 2006a), with a median time from onset of symptoms to death of 9 days (n=76) (http://www.influenzareport.com/links.php?id=16).
The Virus
Infectious diseases are the result of a conflict of interest between macroorganisms and microorganisms. We are not alone on earth.
Requirements for Success
To become a pandemic strain, an influenza virus must comply with a series of requirements. It has to
The Virus 25
•enter the human body and replicate there,
•cause illness in humans, and
•be easily transmittable between humans.
Ideally, it has to be more pathogenic than other competing influenza strains. In the current situation, the potential pandemic virus would have to compete with the already circulating H3N2 and H1N1 strains.
The prerequisite for success is good adaptation: adaptation to human cells; the capability to take over the production machinery of the host cell to produce new offspring; as well as making the individual cough and sneeze to spread the offspring viruses. The clue to success is virulence (Noah 2005, Obenauer 2006, Salomon 2006) – and novelty: if the virus is a true newcomer, most living human beings will have little or no protection at all. The new virus will have unlimited access to virtually every human being and will find a feeding ground of > 6.5 billion human beings. This is one of the biggest biomasses in the world.
The passing of powers from one reigning influenza subtype to a new one is called “antigenic shift” because the antigenic characteristics of the new virus need to shift dramatically to elude the immune system of virtually the entire mankind. Antigenic shift is a major change in the influenza A viruses resulting in new haemagglutinin and/or new neuraminidase proteins. This change may occur by: 1) reassortment of the segmented genome of two parent viruses, or 2) gradual mutation of an animal virus. For reassortment to take place, both the new pandemic candidate virus, normally of avian origin, and an already circulating human virus, i.e., H3N2 or H1N1, need to infect the same human host cell. Inside the cell, genes from both viruses are reassembled in an entirely new virus (they don’t actually have sex, but for didactic purposes, this image might work quite nicely). That’s what happened in 1957 and 1968 (Figure 2).
Reassortment may not be the best route for a candidate pandemic virus. Recent evidence with recombinant viruses containing genes from the 1918 pandemic virus shows that viruses expressing one or more 1918 virus genes were less virulent than the constellation of all eight genes together (Tumpey 2005). The 1918 virus was particular indeed: it appears that it was not the result of a reassortment of two existing viruses, but an entirely avian-like virus that gradually adapted to humans in stepwise mutations (Taubenberger 2005). It is obviously tempting to speculate that the emergence of a completely new human-adapted avian influenza virus in 1918 (n=1) could be deadlier than the introduction of reassortant viruses in 1957 and 1968 (n=2), but such speculation is not scientific. Interestingly – and worryingly –, some amino acid changes in the 1918 virus that distinguish it from standard avian sequences are also seen in the highly pathogenic avian influenza virus strains of H5N1, suggesting that these changes may facilitate virus replication in human cells and increase pathogenicity (Taubenberger 2005).
Virology
Influenza A and B viruses are enveloped viruses with a segmented genome made of eight single-stranded negative RNA segments of 890 to 2,341 nucleotides each (Gürtler 2006). They are spherical or filamentous in structure, ranging from 80 to 120 nm in diameter (Figure 4 and 5). When sliced transversely, influenza virions resemble a symmetrical pepperoni pizza, with a circular slice of pepperoni in the
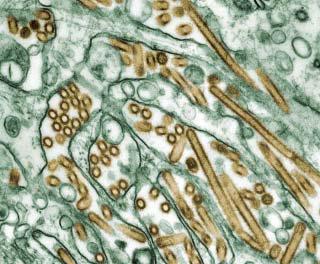
26 Influenza 2006
middle and seven other slices evenly distributed around it (Noda 2006). On the basis of the antigenicity of the surface glycoproteins, haemagglutinin (HA) and neuraminidase (NA), inßuenza A viruses are further divided into sixteen H (H1– H16[Fouchier 2005]) and nine N (N1–N9) subtypes. HA is the major antigen for neutralising antibodies, and is involved in the binding of the virus to host cell receptors. NA is concerned with the release of progeny virions from the cell surface. Currently, only viruses of the H1N1 and H3N2 subtypes are circulating among humans.
Figure 4. Colourised transmission electron micrograph of Avian influenza A H5N1 viruses (seen in gold) grown in MDCK cells (seen in green). Courtesy of CDC/ Cynthia Goldsmith, Jacqueline Katz, and Sharif R. Zaki, Public Health Image Library, http://phil.cdc.gov/Phil/home.asp
Natural Reservoir + Survival
Influenza A viruses occur in a large variety of species, mainly birds, notably aquatic ones, in which infection is largely intestinal, waterborne, and asymptomatic. The domestic duck in Southeast Asia is the principal host of influenza A viruses and also has a central role in the generation and maintenance of the H5N1 virus (Li 2004). In Thailand, there was a strong association between the H5N1 virus and the abundance of free-grazing ducks and, to a lesser extent, native chickens and cocks, as well as wetlands, and humans. Wetlands that are used for double-crop rice production, where free-grazing ducks feed year round in rice paddies, appear to be a critical factor in HPAI persistence and spread (Gilbert 2006).
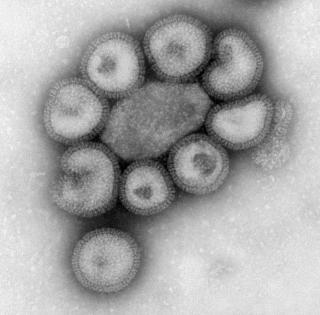
The Virus 27
Figure 5. This negative-stained transmission electron micrograph (TEM) depicts the ultrastructural details of a number virions. Courtesy of CDC/ Dr. F. A. Murphy, Public Health Image Library, http://phil.cdc.gov/Phil/home.asp
Highly pathogenic avian viruses can survive in the environment for long periods, especially in low temperatures (i.e., in manure-contaminated water). In water, the virus can survive for up to four days at 22°C, and more than 30 days at 0°C. In frozen material, the virus probably survives indefinitely. Recent studies indicate that the H5N1 viruses isolated in 2004 have become more stable, surviving at 37°C for 6 days – isolates from the 1997 outbreak survived just 2 days (WHO 20041029). The virus is killed by heat (56°C for 3 hours or 60°C for 30 minutes) and common disinfectants, such as formalin and iodine compounds.
Transmission
Influenza is primarily transmitted from person to person via droplets (> 5 µm in diameter) from the nose and throat of an infected person who is coughing and sneezing (Figure 6). Particles do not remain suspended in the air, and close contact (up to 3–6 feet) is required for transmission. Transmission may also occur through direct skin-to-skin contact or indirect contact with respiratory secretions (touching contaminated surfaces then touching the eyes, nose or mouth). Individuals may spread influenza virus from up to two days before to approximately 5 days after onset of symptoms. Children can spread the virus for 10 days or longer.