

сохраняется в живых лишь 1/4 синиц каждого поколения. По истечении 2-го года от поколения остаются единицы, если это вообще происходит. В лабораторных условиях птицы достигают 9-летнего возраста. В таком случае практически невозможно объяснить, в силу каких обстоятельств естественный отбор мог формировать генетическую программу саморазрушения организма в процессе старения, рассчитанную на 7—8 лет жизни, которые синицами не проживаются.
Изложенное выше не исключает зависимости скорости старения и времени наступления старческих изменений от генетических факторов, однако этими факторами не являются специальные гены или программа. Рассмотрим еще один пример. Для хореи Гентингтона типичным признаком служит сильный тремор (дрожание) головы, конечностей (пляска святого Витта). Симптомы этого наследственного заболевания обычно появляются в возрасте 35—39 лет, причем у мужчин позднее, чем у женщин. Различие в сроках появления болезни объясняется особенностями эволюции мужского и женского генотипов. У мужчин, имеющих по сравнению с женщинами большую продолжительность репродуктивного периода, давление отбора против соответствующего признака угасает с возрастом более медленно. Неблагоприятное фенотипическое действие гена, лежащего в основе хореи Геттингтона, в юношеском и зрелом возрасте подавлялось благодаря присутствию в геноме генов-модификаторов (см. разд. 8.6.1).
Таким образом, из двух принципиально различных направлений в объяснении старения как закономерной стадии онтогенеза в настоящее время более обоснованным является представление, рассматривающее этот процесс как износ биологических структур, а не генетически предопределенное саморазрушение.
8.8. ВВЕДЕНИЕ В БИОЛОГИЮ ПРОДОЛЖИТЕЛЬНОСТИ ЖИЗНИ ЛЮДЕЙ
Продолжительность жизни как житейская проблема связывается в нашем сознании обычно с геронтологией, т.е. с возможностью пережить период зрелости и дожить до преклонного возраста. На самом деле определенная вероятность умереть существует в любом возрасте. Отражением этого является, в частности, широкое использование наряду с видовой или максимальной зарегистрированной продолжительностью жизни такого показателя, как средняя продолжительность жизни. На рис. 8.33 приведена кривая дожития (интенсивности смертности) по возрастам для популяции людей.
Исключительный рост средней продолжительности жизни в экономически развитых странах в XX столетии связан с повышением жизненного уровня, качества питания, жилья, медицинской помощи, улучшением санитарно-гигиенических и эпидемиологических условий. Можно сказать, что в основе демографической революции XX в. лежат социальные по своей природе факторы. Вместе с тем, несмотря на отмеченный рост средней продолжительности жизни людей, форма кривой дожития не меняется. Более того такая же форма кривой воспроизводится в популяциях лабораторных животных, причем относящихся к разным типам
439
животного царства (мышь, крыса, лошадь, головная вошь, плодовая муха).
Это означает, что продолжительность жизни кроме условий существования в немалой степени определяется биологическими факторами. Они отражают особенности структурно-функциональной организации, индивидуального развития, объема приспособительных возможностей и в конечном счете выживаемость отдельных особей. Таким образом, продолжительность жизни отличается исключительной индивидуальной изменчивостью. Указанное свойство проявляется, в частности, в наличии иногда значительного разрыва в значениях средней и максимальной зарегистрированной продолжительности жизни. Так, рекордсмен среди долгожителей японец Шигешио Изуми превысил 120-летний рубеж, тогда как средние показатели длительности жизни в Японии составили 74,1 года для мужчин
и79,6 года для женщин.
8.8.1.Статистический метод изучения закономерностей продолжительности
жизни
Закономерности изменений длительности жизни в историческом времени или от популяции к популяции по понятной причине невозможно изучать на одном отдельно взятом организме. Такие исследования проводят в популяциях с применением статистических методов. Еще в 1825 г. Б. Гомперц описал зависимость увеличения смертности от возраста людей или животных. Позже оказалось, что в отличие от популяций лабораторных животных, вымирающих в силу достаточно комфортных условий существования в точном соответствии с уравнением Гомперца, часть людей умирает вне зависимости от возраста по причине производственных, транспортных и бытовых травм, природных катастроф, неблагоприятных изменений экологии. Указанная особенность была учтена У. Мейкемом, дополнившим уравнение Гомперца слагаемым, не зависящим от возраста. Обобщенный закон Гомперца — Мейкема выглядит следующим образом:
Мt = А + R0ехр(αt)
где Мt — интенсивность смертности (вероятность смерти) людей возраста t (доля ежегодно умирающих в заданном возрасте), А — фоновая компонента смертности, одинаковая для всех возрастных групп (слагаемое Мейкема), R0ехр(αt)—возрастная компонента смертности, отражающая экспоненциальный рост смертности с возрастом (слагаемое Гомперца).
Закон Гомперца — Мейкема описывает изменение вероятности смерти в интервале возрастов от 20 до 80 лет.
Одна из особенностей уравнения Гомперца — Мейкема заключается в том, что оно выделяет в общей смертности две части, которые различаются по природе факторов, их определяющих. Вклад в конечное значение смертности первого слагаемого А определяется условиями жизни, тогда как вклад второго слагаемого R0ехр(αt) зависит от сопротивляемости (жизнеспособности) организма. Фоновая
440
компонента смертности А является социально-контролируемой, а возрастная компонента R0ехр(αt) — социально-независимой. Соответственно первую можно назвать социальной, вторую—биологической.
8.8.2. Вклад социальной и биологической компонент в общую смертность в историческом времени и в разных популяциях
Практическое применение закона Гомперца — Мейкема дает точное представление о природе факторов, обусловливающих различие в интенсивности смертности людей из разных популяций или же из одной популяции, но в разное историческое время. На рис. 8.38 показано, как изменялась общая смертность (7), а также социальная (2) и биологическая (3) ее компоненты в популяции 40-летних женщин Финляндии в интервале с 1890 по 1970 г., т.е. в период исключительного роста средней продолжительности жизни. Нетрудно видеть, что примерно 5-кратное снижение интенсивности смертности к 1970 г. полностью обусловлено уменьшением вклада социально-контролируемой компоненты. Ход кривых 1 и 2 практически совпадает. С другой стороны, доля возрастной компоненты, отражающая состояние биологических механизмов выживания, за описанный исторический период не менялась.
Из представленных материалов вытекают два важных практических следствия. Во-первых, увеличение продолжительности жизни финок целиком связано с повышением жизненного уровня, улучшением социально-гигиенических условий, ростом эффективности профилактической и лечебной медицины. Вовторых, дальнейшее увеличение длительности жизни в описываемой популяции не может быть достигнуто путем изменения социально-контролируемых факторов. Справедливость второго следствия подтверждается незначительным приростом средней продолжительности жизни среди населения Финляндии в последующее десятилетие. Так, если за 1969—1977 гг. названный показатель составлял для женщин Финляндии примерно 76,1 года, то в 1980г.—77,6 года.
Значение биологической компоненты в определении длительности жизни видно из сопоставления ее вклада в интенсивность смертности мужчин и женщин из итальянской популяции (рис. 8.39). Как и в предыдущем примере, за выбранный исторический период отмечается существенное снижение интенсивности общей смертности, причем и у мужчин, и у женщин. Достигнутый к середине 70-х гг. итог для женщин оказался выше, чем у мужчин, за счет биологических особенностей женского организма, что и нашло отражение в уровне возрастной компоненты смертности. Действительно, в 1978 г. средняя продолжительность жизни итальянок составляла 77,4 года, тогда как итальянцев — 70,7. Из материалов по итальянской популяции следует также, что дальнейшее приращение длительности жизни как мужчин, так и женщин требует воздействия на биологические механизмы выживания.
Различия в средней продолжительности жизни мужчин и женщин колеблются от популяции к популяции. Так, в США (1979), Финляндии и Франции (1980) они
441

превышали 8 лет, тогда как в Греции (1981) составляли 4,5 года, а в Болгарии и Японии (1981)—5,5 года. О зависимости возрастной компоненты общей смертности от биологических механизмов свидетельствуют межпопуляционные колебания ее значений в различных европейских странах, в которых достигнут примерно одинаковый уровень жизни (рис. 8.40).
Рис. 8.38. Историческая динамика общей смертности и обеих ее компонент в популяции 40-летних женщин Финляндии (пояснение см. в тексте)
Рис. 8.39. Историческая динамика общей смертности и ее возрастной компоненты среди 40-летних мужчин и женщин (Италия, 1910—1970 гг.):
1—общая стертость среди мужчин, 2—возрастная компонента смертности среди мужчин, 3—общая смертность среди женщин, 4—возрастная компонента смертности среди женщин
442
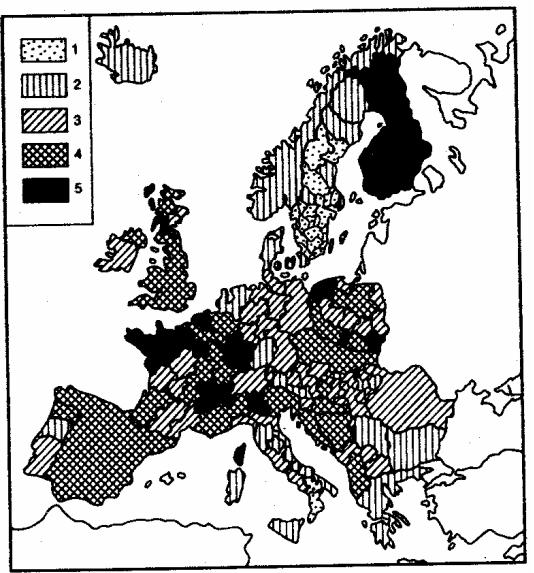
Рис. 8.40. Картограмма значений возрастной компоненты смертности 40-летних мужчин на территории Западной и Центральной Европы:
1—низкий уровень (0,00160 год-1), 2—пониженный уровень (0,00161—0,002220 год- 1), 3—средний уровень (0,00221—0,00280 год-1), 4—повышенный уровень (0,00281— 0.00340 год-1), 5—высокий уровень (свыше 0,00341 год-1)
443