

Биология Ярыгин 2003
.pdfнегомологичными хромосомами приводит к образованию при конъюгации не бивалента, а квадривалента, в котором хромосомы образуют фигуру креста благодаря притягиванию гомологичных участков, расположенных в разных хромосомах (рис. 3.62). Участие в реципрокных транслокациях большего числа хромосом с образованием поливалента сопровождается формированием еще более сложных структур при конъюгации (рис. 3.63).
Рис. 3.62. Образование при конъюгации квадривалента из двух пар хромосом, несущих реципрокную транслокацию
Рис. 3.63. Образование при конъюгации поливалента шестью парами хромосом, участвующих
вреципрокных транслокациях: I — конъюгация между парой
хромосом, не несущих транслокацию; II — поливалент, образуемый шестью парами хромосом, участвующих
втранслокации
Вслучае инверсии бивалент, возникающий в профазе I мейоза, образует петлю, включающую взаимно инвертированный участок (рис. 3.64).
Конъюгация и последующее расхождение структур, образованных измененными хромосомами, приводит к появлению новых хромосомных перестроек. В результате гаметы, получая неполноценный наследственный материал, не способны обеспечить формирование нормального организма нового поколения. Причиной этой является нарушение соотношения генов, входящих в состав отдельных хромосом, и их взаимного расположения.
Однако, несмотря на неблагоприятные, как правило, последствия хромосомных мутаций, иногда они оказываются совместимыми с жизнью клетки и организма и обеспечивают возможность эволюции структуры хромосом, лежащей в основе биологической эволюции. Так, небольшие по размеру делении могут сохраняться в гетерозиготном состоянии в ряду поколений. Менее вредными, чем
141
medwedi.ru

делении, являются дупликации, хотя большой объем материала в увеличенной дозе (более 10% генома) приводит к гибели организма.
Рис. 3.64. Конъюгация хромосом при инверсиях:
I — парацентрическая инверсия в одном из гомологов, II — перидентрическая инверсия в одном из гомологов
Нередко жизнеспособными оказываются робертсоновские транслокации, часто не связанные с изменением объема наследственного материала. Этим можно объяснить варьирование числа хромосом в клетках организмов близкородственных видов. Например, у разных видов дрозофилы количество хромосом в гаплоидном наборе колеблется от 3 до 6, что объясняется процессами слияния и разделения хромосом. Возможно, существенным моментом в появлении вида Homo sapiens были структурные изменения хромосом у его обезьяноподобного предка. Установлено, что два плеча крупной второй хромосомы человека соответствуют двум разным хромосомам современных человекообразных обезьян (12-й и 13-й — шимпанзе, 13-й и-14-й —гориллы и орангутана). Вероятно, эта человеческая хромосома образовалась в результате центрического слияния по типу робертсоновской транслокации двух обезьяньих хромосом.
К существенному варьированию морфологии хромосом, лежащему в основе их эволюции, приводят транслокации, транспозиции и инверсии. Анализ хромосом человека показал, что его 4, 5, 12 и 17-я хромосомы отличаются от соответствующих хромосом шимпанзе перицентрическими инверсиями.
Таким образом, изменения хромосомной организации, чаще всего оказывающие неблагоприятное воздействие на жизнеспособность клетки и организма, с определенной вероятностью могут быть перспективными, наследоваться в ряду поколений клеток и организмов и создавать предпосылки для эволюции хромосомной организации наследственного материала.
142
3.5.4.Значение хромосомной организации в функционировании
инаследовании генетического аппарата
Хромосомный уровень организации наследственного материала обеспечивает
вэукариотической клетке не только определенный характер функционирования отдельных генов, тип их наследования, но и регуляцию их активности.
Хромосома как комплекс генов представляет собой эволюционно сложившуюся структуру, свойственную всем особям данного вида. Взаимное расположение генов в составе хромосомы играет немаловажную роль в характере их функционирования. Расположение гена в той или иной хромосоме определяет тип наследования соответствующего признака (см. разд. 6.3.1).
Принадлежность генов к одной хромосоме обусловливает сцепленный характер наследования детерминируемых ими признаков, а расстояние между генами влияет на частоту рекомбинации этих признаков в потомстве (правило Т. Моргана). Расположение генов в разных хромосомах служит основой независимого наследования признаков (закон независимого наследования признаков Г. Менделя).
Образуя в хромосоме устойчивый комплекс с гистонами, ДНК эукариотической клетки оказывается недоступной для других белков, осуществляющих транскрипцию (РНК-полимераза) и выполняющих регуляторные функции (см. разд. 3.6.6.4). У прокариот кольцевая молекула ДНК, формирующая неустойчивый комплекс с негистоновыми белками, остается доступной для белков, активирующих отдельные гены путем прямого взаимодействия с определенными нуклеотидными последовательностями ДНК. Таким образом, гистоны, участвующие
впространственной организации ДНК в хромосоме, регулируют генную активность, угнетая ее.
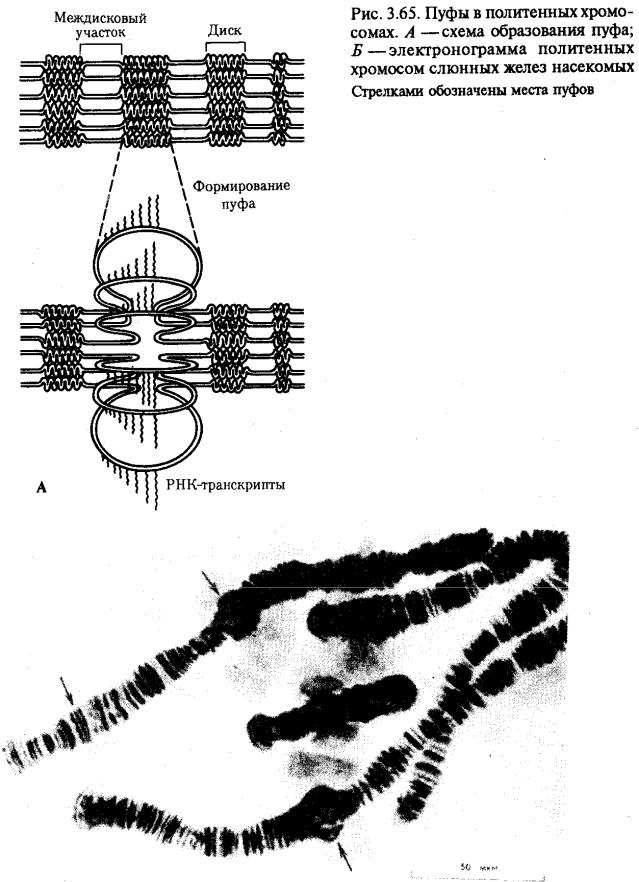
Процесс транскрипции с участка ДНК эукариотической клетки предполагает необходимость предварительной декомпактизации хроматина путем временного ослабления связи ДНК с этими белками. Примером такой декомпактизации хроматина являются пуфы (вздутия), наблюдаемые в участках политенных хромосом и соответствующие активно функционирующим генам (рис. 3.65) или обнаруженные в овоцитах амфибий хромосомы типа ламповых щеток, в которых
активные участки ДНК формируют петлеобразные структуры с меньшей степенью компактизации (рис. 3.66).
143
medwedi.ru
144
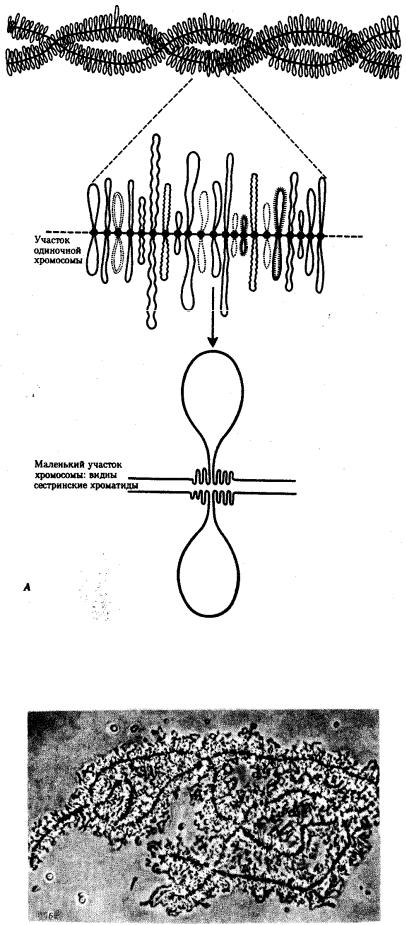
Рис. 3.66. Хромосомы типа ламповых щеток.
А — схема строения; Б — хромосомы из овоцита амфибий
145
medwedi.ru
Рис. 3.66. Продолжение
3.5.5. Биологическое значение хромосомного уровня организации наследственного материала
Возникновение хромосомной организации наследственного материала в эукариотической клетке тесно связано с большим его объемом по сравнению с прокариотической клеткой. Распределение основной массы генетического материала в ограниченном количестве ядерных структур — хромосом — обеспечивает упорядоченность его пространственной организации по группам сцепления. Относительная самостоятельность хромосом в процессах репликации ДНК и распределения ее молекул между дочерними клетками позволяет закономерно воспроизводить и передавать обширную информацию в ряду клеточных поколений, сохраняя постоянство ее организации. Наконец, распределение генов по группам сцепления допускает возможность рекомбинации генетического материала гомологичных и негомологичных хромосом при мейозе и оплодотворении (см. разд. 3.6.2.3).
Эффективность рекомбинации наследственного материала, возрастающая у эукариот благодаря его хромосомной организации, существенно увеличивает степень комбинативной изменчивости у данных организмов. Это является важным эволюционным фактором, обеспечивающим разнообразный исходный материал для естественного отбора.
3.6. ГЕНОМНЫЙ УРОВЕНЬ ОРГАНИЗАЦИИ НАСЛЕДСТВЕННОГО МАТЕРИАЛА
3.6.1. Геном. Генотип. Кариотип
Геномом называют всю совокупность наследственного материала, заключенного в гаплоидном наборе хромосом клеток данного вида организмов. Геном видоспецифичен, так как представляет собой тот необходимый набор генов, который обеспечивает формирование видовых характеристик организмов в ходе их нормального онтогенеза. Например, у некоторых видов появляются гаплоидные организмы, которые развиваются на основе одинарного набора генов, заключенного в геноме. Так, у ряда видов членистоногих гап-лоидными являются самцы, развивающиеся из неоплодотворенных яйцеклеток.
При половом размножении в процессе оплодотворения объединяются геномы двух родительских половых клеток, образуя генотип нового организма. Все соматические клетки такого организма обладают двойным набором генов, полученных от обоих родителей в виде определенных аллелей. Таким образом, генотип — это генетическая конституция организма, представляющая собой совокупность всех наследственных задатков его клеток, заключенных в их хромосомном наборе — кариотипе.
146
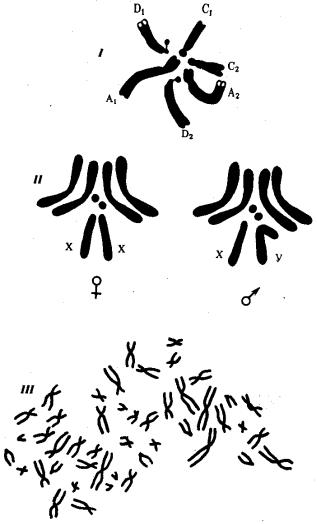
Рис. 3.67. Кариотипы организмов различных видов: I — скерды, II — дрозофилы. III — человека
Кариотип — диплоидный набор хромосом, свойственный соматическим клеткам организмов данного вида, являющийся видоспецифическим признаком и характеризующийся определенным числом, строением и генетическим составом хромосом (рис. 3.67). Ниже приведены количества хромосом соматических клеток некоторых видов организмов.
Если число хромосом в гаплоидном наборе половых клеток обозначить п, то общая формула кариотипа будет выглядеть как 2п, где значение п различно у разных видов. Являясь видовой характеристикой организмов, кариотип может отличаться у отдельных особей некоторыми частными особенностями. Например, у представителей разного пола, имеются в основном одинаковые пары хромосом (аутосомы), но их кариотипы отличаются по одной паре хромосом (гетерохромосомы, или половые хромосомы). Иногда эти различия состоят в разном количестве гетерохромосом у самок и самцов (XX или ХО). Чаще различия
147
medwedi.ru
касаются строения половых хромосом, обозначаемых разными буквами —X и Y (XX или XY).
Каждый вид хромосом в кариотипе, содержащий определенный комплекс генов, представлен двумя гомологами, унаследованными от родителей с их половыми клетками. Двойной набор генов, заключенный в кариотипе,— генотип — это уникальное сочетание парных аллелей генома. В генотипе содержится программа развития конкретной особи.
Животные |
Число хромосом |
Малярийный плазмодий |
2 |
Гидра |
32 |
Таракан |
48 |
Комнатная муха |
12 |
Сазан |
104 |
Окунь |
28 |
Зеленая лягушка |
26 |
Голубь |
80 |
Кролик |
44 |
Шимпанзе |
48 |
Человек |
46 |
3.6.2. Проявление свойств наследственного материала на геномном уровне его организации
3.6.2.1. Самовоспроизведение и поддержание постоянства кариотипа в ряду поколений клеток
В основе самовоспроизведения кариотипа в ряду клеточных поколений лежат процессы самоудвоения отдельных хромосом, описанные в разд. 3.5.3.1. Жизнедеятельность клеток как структурно-функциональных единиц живого обеспечивается всей совокупностью получаемого ими генетического материала, поэтому особое значение приобретает поддержание постоянства кариотипа в ряду их поколений. На геномном уровне организации наследственного материала это обеспечивается всей совокупностью процессов, происходящих в митотическом цикле (см. гл. 2).
148
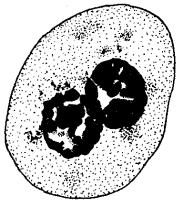
Рис. 3.68. Образование двуядерных клеток в результате торможения цитотомии при их делении
Если содержание ДНК в гаплоидном наборе хромосом, т.е. в геноме, является видоспецифическим и соответствует с, то в диплоидном кариотипе, равном 2n, оно составляет 2с. В ходе митотического цикла количество ДНК в клетке изменяется от 2с (сразу после ее образования, когда хромосомы состоят из одной нити) до 4с (после репликации ДНК в S-периоде интерфазы, когда хромосомы становятся двунитчатыми). Число хромосом при этом остается неизменным и равным 2n на протяжении всего митотического цикла. Благодаря описанным ранее процессам, происходящим в ходе интерфазы и последующего митоза (см. гл. 2), дочерние клетки получают от материнской полноценную наследственную программу — генотип, заключенный в кариотипе.
Однако не всегда клетки, удвоившие свой наследственный материал, вступают в митоз. Этим обусловлено существование в тканях некоторого количества клеток с формулой наследственного материала 2я4с, представляющих определенный резерв, готовый немедленно приступить к делению. Наконец, не всегда начавшееся деление заканчивается образованием двух дочерних клеток. Нарушение деления цитоплазмы при нормальном формировании дочерних ядер приводит к образованию двуядерных клеток (рис. 3.68). Разрушение ахроматанового веретена, делающее невозможным распределение дочерних хромосом между его полюсами, сопровождается появлением полиплоидных клеток (рис. 3.69), в которых 4п дочерних хромосом остаются в одном неразделившемся ядре.
Указанные отклонения от обычного течения митотического цикла являются причиной появления в тканях клеток, несущих больший по объему наследственный материал.
149
medwedi.ru
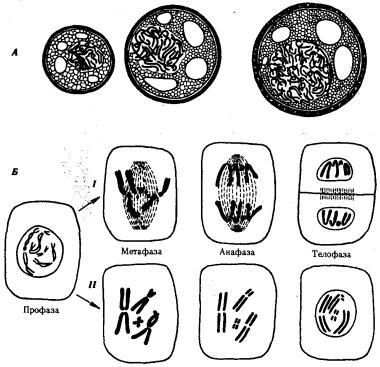
Рис. 3.69. Образование полиплоидньк клеток в результате разрушения веретена деления. А — различные наборы хромосом (12, 24, 48) в пыльцевых зернах одного из сортов гиацинта; Б — образование полиплоидных клеток
/—в норме, II—при разрушении веретена деления колхицином
3.6.2.2.Механизмы поддержания постоянства кариотипа
вряду поколений организмов
Уорганизмов, размножающихся бесполым путем, новое поколение появляется из неспециализированных в отношении генеративной функции клеток тела. В основе их самовоспроизведения лежит митоз, обеспечивающий таким образом сохранение постоянной структуры наследственного материала в ряду поколений не только клеток, но и организмов.
150