

Конспект лекций
.pdfПри замыкании цепи постоянного тока под катодом (действуют допороговым, но продолжительным раздражителем) на мембране возникает стойкая длительная деполяризация, которая не связана с изменением ионной проницаемости мембраны, а обусловлена перераспределением ионов снаружи (привносятся на электроде) и внутри – катион перемещается к катоду.
Вместе со смещением мембранного потенциала смещается и КУД – к нулю. При размыкании цепи постоянного тока под катодом мембранный потенциал быстро возвращается к исходному уровню, а КУД медленно, следовательно, порог увеличивается, возбудимость снижается – катодическая депрессия Вериго.
Изменение возбудимости под анодом.
При замыкании цепи постоянного тока под анодом (допороговый, продолжительный раздражитель) на мембране развивается гиперполяризация за счет перераспределения ионов по обе стороны мембраны (без изменения ионной проницаемости мембраны) и возникающее за ней смещение КУД в сторону мембранного потенциала. Следовательно, порог уменьшается, возбудимость повышается –
анодическая экзальтация.
При размыкании цепи мембранный потенциал быстро восстанавливается к исходному уровню и достигает сниженного уровня критической деполяризации, генерируется потенциал действия. Таким образом, возбуждение возникает только при размыкании цепи постоянного тока под анодом.
Сдвиги мембранного потенциала вблизи полюсов постоянного тока получили название электротонических.
Сдвиги мембранного потенциала не связанные с изменением ионной проницаемости мембраны клетки называют пассивными.
1.7.6. Лабильность (функциональная подвижность). Парабиоз.
Понятие лабильности, или функциональной подвижности, возбудимых тканей ввел Н.Е. Введенский. Мерой лабильности является количество ПД, которое способна генерировать возбудимая ткань в единицу времени. Можно говорить о максимальной частоте раздражения, которую возбудимая ткань способна воспроизводить без трансформации ритма. Лабильность определяется скоростью протекания физиологических процессов в возбудимой ткани и, прежде всего, продолжительностью периода рефрактерности. Например, при продолжительности периода абсолютной рефрактерности 4 мс максимальная частота генерации ПД – 250 Гц.
Уровень лабильности характеризует скорость возникновения и компенсации возбуждения, уровень функционального состояния любых возбудимых клеток и тканей.
Переход от оптимальной к чрезмерной (пессимальной) частоте или силе раздражения вызывает смену возбуждения торможением. Подобные же явления возникают при действии раздражителя с оптимальными характеристиками в случае снижения лабильности возбудимой ткани. При этом развиваются явления, которым Н.Е. Введенский дал название – парабиоз. Парабиоз – это состояние ткани, лабильность которой не удовлетворяет требованиям раздражителя, что затрудняет возникновение и распространение возбуждения.
По мере углубления парабиоза раздражимость ткани проходит несколько стадий: Начальная, уравнительная стадия, когда более сильные раздражения оказываются
для парабиотического участка пессимальными и их эффекты уравниваются с эффектами от слабых раздражений.
Следующая, парадоксальная стадия наступает, когда парабиоз достигает такой степени, при которой сильные раздражения становятся настолько пессимальными, что становятся менее эффективными, чем слабые раздражения.
Наконец, тормозная стадия характеризуется таким низким уровнем лабильности парабиотического участка, при котором любое раздражение вызывает только торможение.
21
1.8. МЕЖКЛЕТОЧНАЯ ПЕРЕДАЧА ВОЗБУЖДЕНИЯ. СИНАПС
Синапсами называют контакты, которые устанавливают нейроны.
Синапс – это специализированная структура, обеспечивающая передачу нервного импульса с аксона на другую клетку.
Существуют классификации синапсов по местоположению, по характеру действия, по способу передачи сигнала.
По местоположению: нервномышечные (мионейрональные), нейросекреторные, нейронейрональные (межнейронные).
Нейрональные: аксосоматические, аксодендритические, аксоаксональные. По характеру действия: возбуждающие и тормозящие.
По способу передачи сигнала: электрические, химические, смешанные.
1.8.1.Химические синапсы.
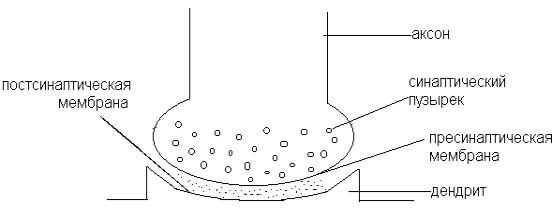
Умлекопитающих и человека в НС чаще встречаются химические синапсы (рис. 11), которые состоят из пресинаптической части (окончание нейрона, передающего сигнал), синаптической щели (непосредственное продолжение межклеточного пространства) и постсинаптической части (структура воспринимающей клетки). При поступлении возбуждения (ПД) к окончанию аксона, в нем высвобождается химическое вещество (медиатор), которое вызывает возбуждение или торможение на мембране иннервируемой клетки.
В синапсе возбуждение всегда передается от пресинаптического (аксонного) участка к постсинаптической области соседней клетки (рис. 11).
Рис. 11. Химический синапс.
Пресинаптическое нервное окончание.
Характерно наличие большого количества субмикроскопических структур округлой формы, которые называют синаптическими пузырьками (везикулами), имеются митохондрии.
Синаптическая щель.
Ширина 10 – 50 нм (100-500 А). При таких размерах электрическая передача возбуждения практически невозможна из-за значительной потери тока во внеклеточной среде, поэтому химическая передача возбуждения представляет собой необходимый усиливающий механизм.
Постсинаптическая мембрана (ПСМ).
Малое количество электроуправляемых натриевых каналов, а потому низкая чувствительность к электрическому току. Следовательно, невозможность генерировать ПД. Зато имеются специфические хемочувствительные рецептороуправляемые каналы.
22
Под влиянием медиатора изменяется проницаемость для ионов K+ и Na+, в результате чего изменяется мембранный потенциал. В зависимости от природы медиатора и свойств рецепторов ПСМ может происходить деполяризация мембраны, что характерно для возбуждения, или гиперполяризация, что типично для торможения. В случае деполяризации ПСМ говорят о возникновении возбуждающего постсинаптического потенциала (ВПСП), а при гиперполяризации – о тормозящем постсинаптическом потенциале (ТПСП).
В состоянии покоя некоторые везикулы с медиатором подходят к пресинаптической мембране (ПреСМ) и медиатор попадает в синаптическую щель, диффундирует, вступает во взаимодействие с рецепторами ПСМ и обусловливает возникновение миниатюрных потенциалов.
При возбуждении пресинаптического окончания взаимосвязь между деполяризацией ПреСМ и высвобождением медиатора обеспечивают ионы Са+, которые поступают снаружи в нервное окончание. Механизм секреции медиатора регулируется рядом биологически активных веществ, в том числе самими медиаторами, а также циклическими нуклеотидами и нейропептидами, которые являются в данном случае модуляторами синаптической передачи.
Основные этапы синаптической передачи.
Приход ПД к пресинаптической мембране, ее деполяризация и генерация на ней потенциала действия.
Проникновение внутрь пресинаптической мембраны ионов кальция – для транспорта везикул с медиатором.
Взаимодействие везикул (синаптических пузырьков) с активными участками пресинаптической мембраны.
Экзоцитоз и выделение квантов медиатора в синаптическую щель (квант медиатора
– это содержимое одной везикулы; квант ацетилхолина примерно 400 – 40000 молекул). Диффузия медиатора к постсинаптической мембране.
Взаимодействие медиатора с клеточными рецепторами ПСМ. Изменение проницаемости для ионов.
Образование постсинаптических потенциалов.
Синапсы и рецепторы постсинаптических мембран этих синапсов могут подразделяться в зависимости от того, какой медиатор выделяется в синаптическую щель. Например:
1.Холинергические (ацетилхолин – АЦХ)
5 – 10% всех синапсов.
Н–холинергические рецепторы (никотинзависимые) в одних случаях (нервномышечные синапсы) блокируются курареподобными веществами, в других (синапсы головного мозга, синапсы вегетативной нервной системы) не реагируют на курареподобные вещества, но блокируются ганглиоблокаторами (бензогексоний и ему подобные вещества).
М–холинергические рецепторы (мускаринзависимые) имеются в центральных холинергических синапсах, а также в окончаниях постганглионарных парасимпатических волокон. Все М-ХР блокируются атропином. В отличие от Н-ХР синапсов, в М-ХР синапсах может иметь место не только возбуждение, но и торможение. Часть М-ХР синапсов являются возбуждающими (гладкомышечные клетки (ГМК) желудочнокишечного тракта и бронхов), а часть – тормозными (в сердечной мышце).
2. Адренергические.
0,5% всех синапсов. Медиатор – норадреналин (НА). Эффект выделения НА зависит от того, какой вид рецептора находится на ПСМ. Различают 4 вида адренорецепторов (АР). Их делят на класс альфа-АР (внутри класса 2 популяции: альфа-1 и альфа-2) и бета-АР (соответственно бета-1 и бета-2). В каждом синапсе есть, вероятно, все 4 вида рецепторов, но доминирует всегда какой-то один.
23
При взаимодействии НА с альфа-1-АР происходит деполяризация ПСМ (образуется ВПСМ). Альфа-1-АР много в ГМК сосудов кожи и желудочно-кишечного тракта, в нейронах головного мозга.
Альфа-2-АР находятся на ПреСМ адренергического синапса, благодаря чему НА оказывает тормозной антидромный эффект (тормозит выделение очередной порции НА).
Бета-1-АР в основном представлены в миокарде, благодаря чему НА вызывает здесь активацию.
Бета-2-АР в основном расположены в ГМК сосудов скелетных мышц, коронаров, в ГМК бронхов, матки. При их активации возникает торможение активности соответствующих структур.
Т.о. возбуждающий или тормозной характер действия медиатора определяется не столько медиатором, сколько свойствами постсинаптической мембраны, находящимися на ней рецепторами.
В ЦНС есть синапсы, медиаторами которых могут быть: серотонин (0,5% всех синапсов); дофамин; гистамин; АТФ; глицин; ГАМК (25-40% синапсов). Глицин, ГАМК – в тормозных синапсах ЦНС.
Инактивация медиатора необходима для реполяризации ПСМ, восстановления исходного потенциала. Например, АЦХ действует на рецепторы ПСМ нервно-мышечного синапса 1-2 мс. Потом часть АЦХ диффундирует в лимфу и кровь, а часть гидролизуется ферментом – ацетилхолинэстеразой на холин и уксусную кислоту, которые снова поступают в пресинаптическое окончание и являются продуктами для синтеза новых порций медиатора.
Для норадреналина ингибитором является моноаминооксидаза и катехолоксиметилтрансфераза.
Но для норадреналина наиболее характерен «обратный захват» пресинаптическими структурами. Несмотря на наличие ингибиторов НА разрушается ими в незначительном количестве и снова депонируется синаптическими пузырьками.
Свойства химических синапсов.
1.Возбуждение проводится в одном направлении.
2.Количество выделяющегося медиатора пропорционально частоте приходящей нервной импульсации. В покое: 1 квант в 1 сек; при ПД – 200 квантов за 2-3 мс.
3.Синаптическая передача не подчиняется закону «все или ничего». Возможна суммация ПСП на ПСМ.
4.Трансформация ритма. Скорость проведения возбуждения в синапсе меньше, чем по нерву. Синаптическая задержка (около 0,5 мс).
5.Высокая чувствительность к химическим веществам, недостатку кислорода.
6.Высокая утомляемость. Развивается в результате длительного высокочастотного стимулирования. Обусловлено истощением и несвоевременным синтезом медиатора в пресинаптической части синапса или глубокой, стойкой деполяризацией ПСМ (пессимальное торможение).
1.8.2. Электрическая передача. 1.8.2.1. Электрические синапсы.
Встречаются редко. ПД вызывает возбуждение в соседней клетке или торможение без химического посредника.
Концептуально были предсказаны еще до открытия химических синапсов (назывались эфапсами). Между 1930 – 1950 г.г., когда концепция химической синаптической передачи была уже общепризнанной, выяснилось, что межклеточная передача возбуждения может осуществляться и электрическим способом.
Принцип: 2 соседние клетки прилегают тесно друг к другу (рис. 12). Сопротивление электрическому току соприкасающихся мембран должно быть сравнимо с сопротивлением остальной, внесинаптической области мембраны.
24
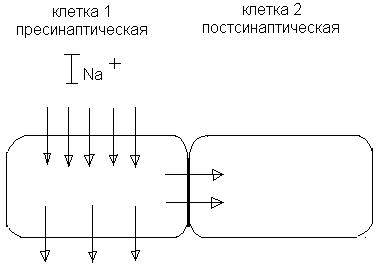
Рис. 12. Электрический синапс.
При возбуждении клетки 1 натриевый ток входит в нее через открытые натриевые каналы и выходит через пока не возбужденные участки мембраны. Часть тока входит через участок мембранного контакта в клетке 2, вызывая ее деполяризацию. Уровень деполяризации здесь примерно в 10 раз ниже, чем в клетке 1, однако выше порога генерирования ПД в клетке 2. Часто такая деполяризация подпороговая, а клетка 2 возбуждается в результате суммации синаптических потенциалов.
Ионы, переносящие электрические токи не могут проходить через липидные мембраны. Для их транспорта в «мембранных контактах» между электрически сопряженными клетками необходимы канальные белки. Такие межклеточные связи называются нексусами, или «щелевыми контактами».
Щелевые контакты соединяют группы синхронно функционирующих клеток.
В тканях, не относящихся к нервной системе, клетки также бывают соединены щелевыми контактами. В миокарде и гладкой мускулатуре эти контакты создают функциональный синцитий. Возбуждение здесь переходит от одной клетки к другой без заметной паузы или снижения амплитуды ПД на границе.
Для таких органов важна регулируемость щелевых контактов. Их каналы закрываются при снижении рН или повышении концентрации Са2+. Это происходит в случае повреждения клеток или глубокого нарушения обмена. За счет такого механизма пораженные места изолируются от остальной части функционального синцития, и распространение патологии ограничивается (например, при инфаркте миокарда).
Кроме этих возбудимых тканей существует и много других, где клетки также соединены щелевыми контактами (все эпителии, печень). В принципе такая связь присуща любой клетке на ранних стадиях эмбрионального развития, когда все клетки соединены между собой щелевыми контактами и сохраняют их до стадии дифференцировки органов.
Щелевой контакт – наиболее распространеный тип электрического синапса. Однако существуют и другие. Например, электрическим путем может передаваться и торможение. В этом случае ПД особым образом расположенных пресинаптических волокон генерируют во внеклеточном пространстве вокруг постсинаптического аксона местный положительный потенциал такой амплитуды, что его деполяризация не может достичь порогового уровня, и проведение по нему ПД блокируется.
Учитывая широкое распространение щелевых контактов, кажется удивительным, почему в нервной системе они не используются для синаптической передачи повсеместно. Видимо, сложнее организованные химические синапсы обеспечивают настолько более высокую специфичность и регулируемость межклеточной коммуникации, что в значительной степени вытеснили электрические.
25
1.8.2.2. Эфаптическая передача.
При некоторых заболеваниях аксоны теряют свою миелиновую оболочку, становясь демиелинизированными. Демиелинизированные аксоны могут вступать в аномальные взаимодействия, когда импульсы, проходящие по группам нервных волокон, индуцируют возбуждение других параллельно идущих аксонов. Это называется эфаптической передачей. Когда такие аномальные ПД генерируются в сенсорных нервных волокнах, появляются аномальные ощущения, парестезии. Если они связаны с ноцицептивными (болевыми) волокнами, то возникают такие неприятные синдромы, как невралгия, каузалгия, невромные боли. Межаксонные помехи могут быть следствием не только недостаточной изоляции (миелиновыми оболочками), но и повышенной возбудимости аксонов.
1.9. ВОЗНИКНОВЕНИЕ ПД В АФФЕРЕНТНЫХ НЕЙРОНАХ. РЕЦЕПТОРНЫЙ И ГЕНЕРАТОРНЫЙ ПОТЕНЦИАЛЫ
Афферентные (сенсорные) нейроны - биполярные нервные клетки, выполняющие функцию восприятия и проведения возбуждения от периферических рецепторов в ЦНС. Тело округлой формы находится вне ЦНС, в спинальном ганглии, имеет один отросток, который затем Т-образно делится. Один отросток идет на периферию и образует там чувствительные окончания (рецепторы). Другой отросток идет в ЦНС, где ветвится и формирует синаптические окончания на вставочных или эффекторных клетках. Тело афферентной клетки в возбуждении участия не принимает, выполняя трофическую функцию. Терминальная же часть афферентного волокна обеспечивает передачу возбуждения от одного рецептора к нескольким вставочным нейронам.
Рецептор является преобразователем внешних стимулов в информационную систему кодируемых нервных импульсов. Рецептор – это специализированная структура (клетка или окончание нейрона), которая в процессе эволюции приспособилась к восприятию соответствующего раздражителя внутренней или внешней среды путем преобразования энергии стимула (раздражителя) в изменение проницаемости своей мембраны. Рецепторы обладают наибольшей чувствительностью к адекватным для них раздражениям.
Классификации рецепторов.
1. По модальности адекватных раздражителей:
Фоторецепторы, хеморецепторы, механорецепторы, терморецепторы, осморецепторы (на изменение осмотического давления), фонорецепторы.
2. По отношению к внешней среде:
Экстерорецепторы – воспринимают информацию из внешней среды: зрение, слух, обоняние, осязание.
Интерорецепторы – воспринимают информацию от внутренних органов: органы пищеварения, сердечнососудистой системы, проприорецепторы мышц и суставов.
Вестибулорецепторы – занимают промежуточное положение, они находятся внутри организма, но возбуждаются внешними факторами.
3. По взаиморасположению раздражителя и рецептора:
Дистантные – воспринимают энергию на расстоянии (зрение, слух, обоняние); Контактные – непосредственный контакт с раздражителем (вкус).
4. По модальности раздражителя:
мономодальные (моносенсорные) – воспринимают один вид энергии (зрение, слух); полимодальные (полисенсорные) – воспринимают несколько видов энергии.
Например, рецепторы роговицы глаза реагируют на изменение температуры и прикосновение;
ноцицепторы (болевые) рецепторы.
5. Гистофизиологическая (структурно-функциональная):
26
первичночувствующие - обоняние, тактильные, проприорецепторы (восприятие стимула осуществляется непосредственно окончанием афферентного нейрона);
вторичночувствующие: вкус, слух, зрение, вестибулорецепторы (здесь между действующим стимулом и афферентным нейроном располагается специализированная клетка эпителиального происхождения, из которой при раздражении выделяется медиатор, действующий на окончание афферентного нейрона).
Процесс преобразования энергии стимула (сигнала) в изменение проницаемости мембраны с последующим формированием рецепторного потенциала мембраны получил название трансдукции и включает в себя 3 основных этапа:
1)взаимодействие стимула с рецепторной белковой молекулой, которая находится
всоставе клеточной мембраны рецепторной клетки;
2)внутриклеточные процессы усиления и передачи сенсорного стимула в пределах рецепторной клетки;
3)открывание находящихся в мембране рецептора ионных каналов, через которые начинает течь ионный ток, что, как правило, приводит к деполяризации клеточной мембраны рецепторной клетки (возникает рецепторный потенциал). В фоторецепторах, наоборот, возникает гиперполяризация.
Зависимость между величинами стимула и рецепторного потенциала логарифмическая.
Упервичночувствующих рецепторов рецепторный потенциал является одновременно и генераторным, т.к. вызывает генерацию ПД в наиболее чувствительных участках мембраны.
Увторичночувствующих рецепторов рецепторный потенциал вызывает выделение квантов медиатора из пресинаптических окончаний рецепторной клетки. Медиатор изменяет поляризацию ПСМ, т.е. здесь генераторный потенциал является постсинаптическим потенциалом первого нейрона сенсорной системы.
Свойства рецепторных и генераторных потенциалов:
- градуальны (стимулами разной интенсивности деполяризуются или гиперполяризуются неодинаково), их амплитуда отражает силу стимуляции, хотя последняя не служит для них источником энергии;
-локальны – распространяются по мембране электротонически, а не активно;
-подвергаются пространственной и временной суммации (два слабых одиночных стимула вместе могут вызвать надпороговую деполяризацию).
Трасформация генераторных потенциалов в залпы ПД обычно происходит на первом перехвате Ранвье афферентного нервного волокна. У немиелинизированных афферентов точное место трансформации неизвестно. Генераторный потенциал распространяется электротонически до места генерирования ПД, точно также как синаптический потенциал по телу мотонейрона к аксонному холмику.
Частота импульсации в афферентном нервном волокне пропорциональна величине генераторного потенциала.
Такое же перекодирование локального потенциала с переменной амплитудой в проводимый сигнал с переменной частотой происходит в синапсах ЦНС.
1.10. ВОЗНИКНОВЕНИЕ ПД В ЭФФЕРЕНТНЫХ НЕЙРОНАХ. МЕХАНИЗМЫ СУММАЦИИ ПСП
Вставочные нейроны cоставляют 90% всех нейронов. Отростки не покидают пределов ЦНС, но обеспечивают многочисленные связи по горизонтали и вертикали. Осуществляют обработку информации и связь между афферентными и эфферентными нейронами. Делятся на возбуждающие и тормозные.
27

Эфферентные (эффекторные) нейроны - это нейроны, передающие информацию от нервного центра к исполнительным органам.
Пирамидные клетки двигательной зоны коры больших полушарий, посылающие импульсы к мотонейронам передних рогов спинного мозга.
Мотонейроны – аксоны выходят за пределы ЦНС и заканчиваются синапсом на эффекторных структурах.
Терминальная часть аксона ветвится, но есть ответвления и вначале аксона – аксонные коллатерали. Место перехода тела мотонейрона в аксон – аксонный холмик – наиболее возбудимый участок. Здесь генерируется ПД, затем распространяется по аксону.
На теле нейрона огромное количество синапсов.
Если синапс образован аксоном возбуждающего интернейрона, то при действии медиатора на постсинаптической мембране возникает ВПСП.
Если синапс образован аксоном тормозной клетки, то при действии медиатора на постсинаптической мембране возникает гиперполяризация или ТПСП.
Алгебраическая сумма ВПСП и ТПСП на теле нервной клетке проявляется в возникновении ПД в аксонном холмике, т.е. в области аксонного холмика происходит интеграция событий, разыгрывающихся на отдельных участках мембраны нейрона
(рис.13).
Временная (последовательная) суммация. Если с определенным интервалом к нейрону в точку А приходят импульсы, они вызывают ВПСП. Если эти ВПСП не достигают КУД, то ПД не возникает. Если же частота следования импульсов большая, то в этом месте происходит суммация ВПСП и при достижении КУД нейрон возбуждается.
Пространственная суммация. Возбуждения приходящие одновременно в разные точки нейрона (А, В, С), даже если они сами по себе подпороговые, могут привести к возбуждению, при условии, что суммированный ВПСП достигнет КУД.
Рис. 13. Последовательная и пространственная суммация на уровне нейрона.
1 – приход импульса к нервной клетке;
2 – формирование биопотенциала.
28
1.11. СКЕЛЕТНЫЕ МЫШЦЫ
Скелетные мышцы обеспечивают перемещение составных элементов скелета. Имеют поперечную исчерченность (поперечнополосатые).
Свойства скелетной мышцы: 1) возбудимость; 2) проводимость (способность проводить ПД вдоль мышечного волокна и вглубь его по Т-системе поперечных трубочек, служащих связующим звеном между поверхностной мембраной и сократительным аппаратом); 3) сократимость (способность укорачиваться или развивать напряжение при возбуждении); 4) эластичность (способность развивать напряжение при растягивании).
Скелетные мышцы имеют 2 типа волокон: интрафузальные и экстрафузальные. Интрафузальное волокно находится внутри мышечного веретена (специализированного мышечного рецептора), располагающегося в толще скелетной мышцы. Оно необходимо для регуляции чувствительности рецептора и управляется специальными мотонейронами спинного мозга – гамма-мотонейронами. Мышечные волокна не входящие в состав мышечного веретена, называются экстрафузальными.
Особенности нервно-мышечного (мионеврального) синапса.
Наличие большого числа изгибов на пресинаптической и постсинаптической мембранах, которые увеличивают площади пресинапса и постсинапса, а, следовательно, и вероятность взаимодействия.
В пресинапсе (в основном в активных зонах) – везикулы с АЦХ (до 100010000 молекул).
Постсинаптическая мембрана в виде гребешков (с интервалом 1 мкм). На вершине гребешка концентрация Н-ХР максимальна (2000 на 1 мкм2, в устьях – 1000, а во внесинаптической зоне 50 на 1 мкм2). В синаптической щели находится гликокаликс – волокна, выполняющие опорную функцию.
Здесь расположена ацетилхолинэстераза (АЦХЭ), скорость расщепления АЦХ 1мол/мс.
Деполяризация ПСМ носит здесь название потенциала концевой пластинки (ПКП). В покое выделяется 1 квант/с – миниатюрный потенциал концевой пластинки (МПКП). При ПД в синапсе лягушки выделяется 100 квантов медиатора, а у млекопитающих 200300 квантов медиатора.
МПП мышечных волокон примерно – 90 мв. ПД – 120-130 мв. Длительность ПД 1- 3 мс. КУД – 50 мв.
Механизмы блокады нервно-мышечной передачи.
1.Блокада проведения возбуждения в пресинаптичесой части (местноанестезирующие вещества).
2.Блокада высвобождения медиатора в пресинаптической части (токсин ботулизма).
3.Нарушение синтеза медиатора.
4.Блокада холинорецепторов (бунгаротоксин), вытеснение АЦХ из рецепторов (кураре), инактивация ПСМ (сукцинилхолин).
5.Угнетение ацетилхолинэстеразы (фосфорорганические соединения). Приводит к длительному сохранению АЦХ и вызывает длительную деполяризацию и инактивацию рецепторов синапсов.
Понятие о нейромоторной единице или двигательной единице (ДЕ).
Это морфологический комплекс, состоящий из двигательного нейрона (альфамотонейрона, расположенного в спинном мозге или в стволе мозга) и иннервируемых им группы мышечных волокон. Число иннервируемых мышечных волокон может варьировать от нескольких единиц до нескольких сотен (ДЕ – 10-1000 волокон).
Виды сокращений:
1. Динамический – чередование сокращения и расслабления.
2. Статический – длительное сокращение без изменения длины мышцы.
Режимы сокращений:
29
1.Изотонический – напряжение остается постоянным, длина мышцы
уменьшается;
2.Изометрический – увеличение напряжения при постоянной длине мышечного волокна;
3.Ауксотонический – физиологический режим сокращения, при котором длина уменьшается, напряжение увеличивается.
Классификация скелетных мышечных волокон.
Подразделяются на фазические (фазные – они генерируют ПД) и тонические (не способны генерировать полноценный ПД распространяющегося типа).
Медленные фазические волокна окислительного типа.
-большое содержание миоглобина (красные мышцы)
-большое число митохондрий
-утомление наступает медленно, а восстановление быстро
-нейромоторные единицы состоят из большого числа волокон.
Быстрые фазические окислительного типа.
-быстрые сокращения без заметного утомления
-большое количество митохондрий
-число волокон нейромоторной единицы меньше, чем в предыдущей группе.
Быстрые фазические с гликолитическим типом окисления.
-миоглобин отсутствует (белые мышцы)
-АТФ образуется за счет гликолиза
-Митохондрий меньше, чем у волокон окислительного типа
Для всех фазических волокон характерно наличие одной, в крайнем случае нескольких концевых пластинок, образованных одним двигательным аксоном.
Быстрые фазические волокна имеют более развитую саркоплазматическую сеть и обширную сеть Т-системы, чем медленные.
Тонические волокна (медленные).
Двигательный аксон образует множество синаптических контактов с мембраной мышечного волокна.
Сокращения и расслабления происходят медленно, низкая активность миозиновой АТФ-азы.
Эффективно работают в изометрическом режиме.
Не генерируют ПД и не подчиняются закону «все или ничего». Одиночный пресинаптический импульс вызывает незначительное сокращение. Серия вызывает суммацию ПСП и плавно нарастающую деполяризацию мышечного волокна (входят в состав наружных мышц глаза).
Одиночное мышечное сокращение.
1. Латентный период. 2. Фаза сокращения (укорочения) мышцы. 3. Фаза расслабления.
Суммированные сокращения.
В зависимости от частоты раздражения меняется характер сокращения.
Если стимулы попадают в латентные периоды – наблюдаются одиночные сокращения.
Если очередной стимул (или его действие) попадает в фазу расслабления, мышца не успевает расслабиться, возникает дополнительное сокращение, развивается длительное напряжение - зубчатый тетанус.
При более высокой частоте (т.е. с еще меньшим интервалом между раздражителями), когда каждый очередной стимул попадает в фазу укорочения мышцы, происходит продолжительная активация сократительной системы, развивается мощное длительное сокращение, которое называется гладким тетанусом. Расслабление возникает при утомлении.
Амплитуда гладкого тетануса зависит от частоты раздражения. Если каждый последующий стимул (раздражитель) попадает в фазу экзальтации (повышенной
30