

lek05 / Лекция 5 Регуляция активности генов
.doc
Конспект лекции № 5.
Тема. Регуляция биосинтеза белка. Понятие о гомеостазе. Понятие о точковых мутациях.
Многоклеточный организм человека возникает при слиянии сперматозоида и яйца в единственную клетку – зиготу, содержащую 46 хромосом. Затем из зиготы, путем митотического деления, вырастает полноценный организм из огромного числа разнообразных клеток. В ядре каждой их этих десятков триллионов клеток есть все те же 46 хромосом (46 молекул нуклеопротеидов). При этом последовательность нуклеотидов в ДНК ядра зиготы и ДНК ядра любой клетки (печени, кожи, мозга) одинаковы. Но тогда возникает вопрос, почему клетки нашего организма не похожи друг на друга и почему они выполняют разные функции.
Серьезный шаг по решению этой проблемы был сделан Жакобом и Моно. Они выдвинули гипотезу о том, что у бактерий гены (участки молекулы ДНК, несущие элементарную функционально значимую информацию) работают в клетке не одновременно. Большую часть времени большая часть генов выключена "отдыхает", небольшая часть генов включена "работает". Главный механизм включения и выключения генов Жакоб и Моно связали с понятием о генах-операторах, белках репрессорах и индукторах.
По гипотезе Жакоба-Моно, перед обычным структурным геном находится ген-оператор, обеспечивающий фиксацию фермента РНК-полимеразы (участок для непосредственного присоединения РНК полимеразы называется промотор) и начало транскрипции. Обычно, с геном-оператором связан белок-репрессор и в таком состоянии фермент РНК-полимераза не может связаться с геном оператором и начать считывать информацию структурных генов ДНК (рис. 1).
Рисунок 1. Выключенное (исходное) состояние структурных генов.
ген-оператор структурные гены
───────────────|───────────────────────
───┬═══════════|───────────────────────
└───────────┘ - белок-репрессор
│ │ │
╚════════╩═╝ - РНК-полимераза
Чтобы начать процесс считывания информации (транскрипцию), надо освободить ген-оператор от белка-репрессора. Роль "освободителя" берет на себя молекула-индуктор (дерепрессор). Индуктор связывается с белком-репрессором и отрывает его от гена-оператора (рис. 2 а и 2 б).
Рисунок 2 а. Связывание индуктора с белком-репрессором.
ген-оператор структурные гены
───────────────|───────────────────────
───┬═══════════|───────────────────────
╞═══════════╡ - белок-репрессор
└───────────┘ - индуктор
│ │ │
╚════════╩═╝ - РНК-полимераза
Рисунок 2 б. Освобождение гена-оператора от белка-репрессора (дерепрессия).
ген-оператор структурные гены
───────────────|───────────────────────
───────────────|───────────────────────
┌───────────┐ - белок-репрессор
╞═══════════╡
└───────────┘ - индуктор
│ │ │
╚════════╩═╝ - РНК-полимераза
Теперь ген-оператор свободен и к нему прикрепляется фермент РНК-полимераза, начиная транскрипцию – синтез м-РНК. Структурные гены включаются, и начинается транскрипция и трансляция (рис. 3).
Рисунок 3. Структурные гены включены, идет биосинтез белка.
ген-оператор структурные гены
───────────────|──────────────────────
─────┬────────┬|┬─────────────────────
╚════════╩═╝~~~~~~~~~~~~~~~~~~~~ - м-РНК (транскрипция)
РНК-полимераза
~~~~~~~~~~~~~~~~ - и-РНК (процессинг)
┬┬┬┬┬┬┬┬┬┬┬┬┬┬┬┬┐ - белок-продукт
┴┴┴┴┴┴┴┴┴┴┴┴┴┴┴┴┘ (трансляция)
При накоплении достаточного количества белка-продукта происходит реакция его взаимодействия с комплексом индуктор – белок-репрессор. При этом молекулы индуктора разрушаются. Белок-репрессор освобождается (рис. 4).
Рисунок 4. Разрушение молекулы индуктора и освобождение белка-репрессора.
белок-репрессор
┌───────────┐
╞═══════════╡ ┬┬┬┬┬┬┬┬┬┬┬┬┬┬┬┬┐ - белок-продукт
└───────────┘ ┴┴┴┴┴┴┴┴┴┴┴┴┴┴┴┴┘
индуктор
белок-репрессор
┌───────────┐
└───────────┘ ┬┬┬┬┬┬┬┬┬┬┬┬┬┬┬┬┐ - белок-продукт-терминатор индуктора
┴┴┴┴┴┴┴┴┴┴┴┴┴┴┴┴┘
┌──┐┌───────┐
└──┘└───────┘ -распад индуктора
В заключении освобожденный белок-репрессор вытесняет РНК-полимеразу со своего "законного" места и закрывает подход к структурным генам. Тем самым, процесс биосинтеза белка прекращается – структурные гены выключаются – переходят в исходное состояние (рис. 5).
Рисунок 5. Выключенное (исходное) состояние структурных генов
ген-оператор структурные гены
───────────────|───────────────────────
───┬═══════════|───────────────────────
└───────────┘ - белок-репрессор
│ │ │
╚════════╩═╝ - РНК-полимераза
Гипотеза Жакоба-Моно была обоснована результатами исследования культуры кишечной палочки, жившей на питательной среде с глюкозой. При пересадке ее на питательную среду с лактозой скорость роста колонии снижалась, но затем восстанавливалась после включения гена, ответственного за синтез ферментов расщепляющих лактозу. В этом случае, индуктором была сама лактоза, а реактиватором белка-репрессора был белок-продукт (фермент для расщепления лактозы).
У более сложных эукариотов и многоклеточных организмов включение и выключение генов может опосредоваться несколькими промежуточными веществами и медиаторами нервной системы. Тем не менее, общая схема регуляции активности генов остается подобной той, что описана у Жакоба и Моно.
Дальнейшие исследования по клонированию лягушки (Гёрдон) и млекопитающих, доказали определяющее значение белков-репрессоров и индукторов в индивидуальном развитии особей.
Открытие системы белков-репрессоров, контролирующих большую часть структурных генов, и принципиальная возможность их дерепрессии открывает перед медицинской генетикой фантастические перспективы. Если научиться избирательно включать и выключать определенные гены, то можно, например, восстанавливать у больных людей любые органы и даже ампутированные конечности.
Понятие о гомеостазе. Кибернетические основы поддержания гомеостаза.
Живые организмы подчиняются общим закономерностям кибернетики (науки об управлении), открытым Норбертом Винером (1948).
Стационарное состояние любой живой системы создает оптимальные условия для выживания в изменяющихся условиях среды обитания.
Гомеостаз – это постоянство внутренней среды организма.
Адаптация (приспособление) – это система механизмов, поддерживающих гомеостаз.
Классическая схема кибернетической регуляции живой системы состоит из 4 главных элементов (рис. 6): 1-входа, 2-модулятора, 3-выхода 4-обратной связи (отрицательной или положительной).
Рисунок 6. Элементы кибернетической системы.
МОДУЛЯТОР
┌────────────────────────────────────────┐
ВХОД ──│ детектор + регулятор + эффектор │── ВЫХОД
└────────────────────────────────────────┘
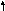 ОБРАТНАЯ
СВЯЗЬ │
ОБРАТНАЯ
СВЯЗЬ │
─────────────────────────────────────────────
Модулятор – центральное звено кибернетической системы состоит из 3 частей.
Детектор - устройство контроля состояния системы (осморецепторы раздражаются ионами Na+).
Регулятор - устройство для анализа информации и выработки ответа на сигнал от детектора (гипоталамус раздражается и выделяет антидиуретический гормон – АДГ).
Эффектор – устройство обеспечения ответной реакции (почки под воздействием АДГ увеличивают обратную реабсорбцию воды).
Вход – это внешнее воздействие – раздражение, которое приводит к отклонению параметра системы (организма) от стационарного состояния (после употребления соленой пищи растет концентрация натрия в крови).
Выход – это результат реагирования модулятора системы на отклонении ее параметра от стационарного состояния (появление чувства жажды и утоление его водой).
Обратная связь – влияние выхода на вход (увеличение воды в организме снижает влияет на концентрацию соли в крови).
Обратная связь может быть отрицательной (не значит, что плохой) и положительной (не значит, что хорошей).
Отрицательная обратная связь препятствует отклонению системы от стационарного состояния, уменьшая ("-") отклонение по модулю. Ее результат: восстановлен исходный уровень параметра.
Так, для нормальной работы организма необходимо поддерживать концентрацию глюкозы в крови на уровне около 4 мМоль/л (3,6 - 5,5) - это исходный уровень – стационарное состояние. Повышение или снижение концентрации глюкозы называют отклонением от исходного уровня параметра гомеостаза. После этого, в живом организме включаются механизмы, которые могут вернуть концентрацию глюкозы к исходному значению.
Например, если человек съел сладкое или мучное, то уровень глюкозы в крови повышается. Включается парасимпатическая нервная система и вырабатывается гормон инсулин, который переводит глюкозу из крови в клетки (снижение концентрации глюкозы в крови).
Наоборот, когда при физической нагрузке (затрата энергии и уменьшение концентрации глюкозы в крови) включается симпатическая нервная система, в ответ вырабатывается адреналин и переводит глюкозу из депо (гликоген в печени) в кровь (концентрация глюкозы в крови повышается).
В обоих случаях система возвращается в исходное стационарное состояние.
В этих примерах произошло поддержание гомеостаза по принципу отрицательной обратной связи. Уменьшается отклонение от нормы по модулю. Т.о., если ответ организма уменьшает « - » отклонение от стационарного состояния, то обратная связь отрицательная.
Если ответ организма увеличивает « + » отклонение от стационарного состояния, то обратная связь положительная.
Например, при больном сердце уменьшается кровоток во всех органах, в том числе, через почки. Снижение почечного кровотока раздражает юкста-гломерулярные клетки почек, которые вырабатывают гормон ренин. Ренин запускает систему, которая задерживает в организме воду и суживает артерии. При этом нагрузка на сердце возрастает, и оно еще хуже выбрасывает кровь. Но, чем слабее работает сердце, тем меньше кровоток через почки и они выбрасывают еще больше ренина, а чем больше ренина, тем труднее сердцу. Формируется порочный круг болезни. В результате, при отсутствии лечения больной умирает от сердечнососудистой недостаточности.
Чаще всего в медицине рассматривают возникновение положительной обратной связи, как «плохое» явление, ведущее к катастрофическим для организма последствиям, а регуляцию с помощью отрицательной обратной связи как «хорошее» физиологическое явление.
Однако возникновение положительной обратной связи не всегда приводит к плохим последствиям. Считается, что потенциал действия в нервных и мышечных клетках является нормальным процессом, который запускается по механизму положительной обратной связи. Небольшая частичная деполяризация мембраны может включить механизм перезарядки мембраны – основу нормальных электрофизиологических процессов организма.
Отрицательная обратная связь тоже не всегда «хорошая». Так, у здорового человека в головном мозге постоянно вырабатываются эндорфины – вещества, обеспечивающие определенный порог болевой чувствительности. Поэтому мы не чувствуем боли при движении суставов, органов пищеварения при перистальтике. При серьёзных повреждениях организма собственных эндорфинов не хватает и для снятия боли вводят наркотические анальгетики (морфин, омнопон, промедол). Но, они не только снимают боль. По механизму отрицательной обратной связи происходит снижение выработки собственных анальгетиков – эндорфинов. Длительный прием наркотиков делает мозг «ленивым» и он «забывает», как делать эндорфины. Внезапная отмена наркотика, ставит больного в беззащитное состояние - он уже не может терпеть обычные нервные сигналы от рецепторов организма. Простые раздражения вызывают боль – наступает «ломка» - абстинентный синдром. Организм попадает в лекарственную зависимость от наркотика.
Фармакологические принципы регуляции больных организмов основаны на знании кибернетических механизмов их физиологической регуляции. Эти принципы включают применение средств действующих на рецепторы (адреномиметики, холиномиметики, адреноблокаторы, холинолитики), транспорт ионов через мембраны (блокаторы кальциевых каналов), концентрацию гормонов (глюкокортикоиды) и т. п.
Важным примером нарушения гомеостаза является изменение рН - водородного показателя – главного показателя кислотно-щелочного равновесия в клетках организма. Нормальное функционирование клеток и всего организма возможно только при строго определенных значениях рН. Так в клетках границы нормы для водородного показателя укладываются в промежуток от 7,2 до 7,4. Сдвиг рН в кислую сторону (рН<7,2) называют ацидоз, а в щелочную (рН>7,4) называют алкалоз. В обоих случаях происходит изменение концентрации ионов водорода и нарушение работоспособных структур белковых молекул. Органические молекулы перестают выполнять свои функции, что приводит к тяжелым нарушениям здоровья и гибели организмов.
Поддержание гомеостаза рН обеспечивают буферные системы организма (гемоглобиновая, гидрокарбонатная, фосфатная, белково-аминокислотная) и некоторые органы (почки, легкие). Эти системы и органы препятствуют изменению рН, связывая ионы водорода (Н+) или гидроксил-ионы (ОН-). Так, например, при активной физической работе в клетках сердца возникает недостаток кислорода и накапливается молочная кислота. Она диссоциирует с образованием ионов водорода. Возникает угроза ацидоза и инфаркта миокарда. Но молочная кислота действует и на артерии сердца - они расширяются. Это приводит к улучшении доставки кислорода, а в его присутствии молочная кислота окисляется до углекислого газа и воды. Концентрация ионов водорода возвращается к норме. Углекислый газ и вода удаляются легкими и почками. В этом случае гомеостаз не нарушается.
Нарушение правильной последовательности отдельных нуклеотидов в молекуле ДНК или РНК называют точковыми мутациями. Они возникают при действии мутагенных факторов: физических (ионизирующая радиация), химических (лекарства), биологических (вирусы).
Возможны 4 типа точковых мутаций.
1. Замещение. Место одного нуклеотида заменяет другой.
…ААА ТТТ ЦАЦ ЦГА ГГГ ... - норма
…ААТ ТТТ ЦАЦ ЦГА ГГГ ... - мутация
Последствия замещения (для собираемой белковой молекулы):
а - возникновение стоп-кодона и прекращение сборки белка
(УГЦ-Цис УГА-стоп)
б - замена одной аминокислоты (ААЦ-Асн ААА-Лиз)
в - сборка нормального белка, если замещение попало на вырожденный код (ААА-Лиз ААГ-Лиз)
2. Инверсия (вращение). Соседние нуклеотиды меняются местами
…ААА ТТТ ЦАЦ ЦГА ГГГ ... - норма
…ААТ АТТ ЦАЦ ЦГА ГГГ ... - мутация
Последствия инверсии (для собираемой белковой молекулы):
а - возникновение стоп-кодона и прекращение сборки белка
(УАЦ ААА-Тир Лиз УАА ЦАА-стоп Глн)
б - замена двух аминокислот (ААА ЦЦЦ-Лиз Про ААЦ АЦЦ-Асн Тре)
в - замена одной аминокислоты, если инверсия между соседними триплетами у одного из них попала на вырожденный код
(ААА ГГГ-Лиз Гли ААГ АГГ-Лиз Арг)
г - сборка нормального белка, если инверсия между соседними триплетами обоих нуклеотидов попала на вырожденный код
(ЦЦЦ АГГ-Про Арг ЦЦА ЦГГ-Про Арг)
3. Вставка. В последовательность нуклеотидов вставляется новый лишний нуклеотид
…ААА ТТТ ЦАЦ ЦГА ГГГ... - норма
…ААА ЦТТ ТЦА ЦЦГ АГГ Г... - мутация
Последствия вставки (для собираемой белковой молекулы):
а - возникновение стоп-кодона и прекращение сборки белка (УАЦ ААА-Тир Лиз УАА ЦАА А стоп-Глн)
б - сдвиг рамки триплетного кода и сборка бессмысленного белка
...ААА УУУ ЦАЦ ЦГА ГГГ...- Лиз Фен Гис Арг Гли…
...ААА ЦУУ УЦА ЦЦГ АГГ Г...- Лиз Лей Сер Про Арг…
4. Выпадение (делеция). Из последовательности нуклеотидов теряется нормальный нуклеотид
...АААТТТЦАЦЦГАГГГА... - норма
...ААТТТЦАЦЦГАГГГА... - мутация
Последствия вставки (для собираемой белковой молекулы):
а - возникновение стоп-кодона и прекращение сборки белка (УАЦ ААА-Тир Лиз УАА АА- стоп-)
б - сдвиг рамки триплетного кода и сборка бессмысленного
...ААА УУУ ЦАЦ ЦГА ГГГ...- Лиз Фен Гис Арг Гли…
...ААА УУЦ АЦЦ ГАГ ГГ...- Лиз Фен Тре Глу…
Редким видом последствий (для собираемой белковой молекулы) любого вида точковой мутации может быть исчезновение стоп-кодона.
Для реального организма любая мутация, ведущая к нарушению последовательности нуклеотидов, может иметь 3 физиологических последствия:
1 - смерть;
2 - болезнь, но возможно она проявится через несколько поколений;
3 - нет заметных изменений здоровья, если изменилась часть белка, не имеющая важного функционального значения.
Блок дополнительной информации.
Информация о структуре белков и РНК записана в участках ДНК, называемых генами и цистронами.
Ген - это участок ДНК, кодирующий один белок.
Цистрон же - участок ДНК, кодирующий одну полипептидную цепь.
Таким образом, если белок состоит из нескольких разных полипептидных цепей (субъединиц), то его ген включает несколько цистронов (рис. 1).
Однако такое подразделение относится, в основном, к бактериям, где цистроны одного гена обычно следуют на ДНК друг за другом.
У животных же и человека цистроны нередко располагаются в разных хромосомах и обычно тоже называются генами: например, ген альфа-цепи и ген бета-цепи гемоглобина.
Рис. 1. Ген и цистрон
К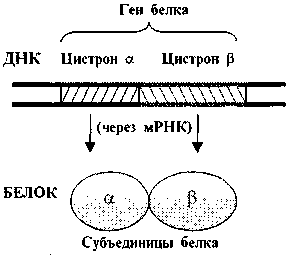 роме
генов всех белков организма, в хромосомах
имеются также гены РНК. Среди них четыре
вида генов для рибосомных РНК и нескольких
десятков генов для транспортных РНК.
роме
генов всех белков организма, в хромосомах
имеются также гены РНК. Среди них четыре
вида генов для рибосомных РНК и нескольких
десятков генов для транспортных РНК.
Общая совокупность генов, определяющих наследственную информацию организма, называется геномом.
Всего в геноме бактерий (конкретно - кишечной палочки) - около 2500 цистронов. В хромосомах человека число генов около 30 000.
Почти все гены эукариот (в отличие от генов прокариот) имеют характерную особенность: содержат не только кодирующие участки - экзоны, но и некодирующие - интроны (рис. 2.). Экзоны и интроны перемежаются друг с другом, что придает гену как бы «разорванную» структуру.
Рис. 2. Экзоны и интроны.
Ч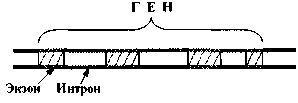 исло
интронов в гене варьирует от 2 до
нескольких десятков; в гене миозина их
около 50. Порой на интроны приходится до
90 % общей длины гена.
исло
интронов в гене варьирует от 2 до
нескольких десятков; в гене миозина их
около 50. Порой на интроны приходится до
90 % общей длины гена.
Между генами также находятся некодирующие последовательности - спейсеры. Несмотря на общее название, функциональная роль их может быть различной.
Многие спейсерные участки, видимо, выполняют структурную роль: - участвуют в правильной укладке нуклеосомной цепи в высшие структуры хроматина, в прикреплении хромосом к аппарату центриолей и т. д. Другие спейсеры являются специфическими местами (локусами) связывания определенных белков функционирующих на ДНК (ДНК-полимеразного комплекса и белков, выполняющих регуляторную функцию).
При этом участки связывания РНК-полимеразы (фермента, синтезирующего РНК на ДНК) называются промоторами. Они либо вплотную примыкают к началу гена (или группы генов), либо отделены от гена какими-либо другими функциональными локусами.
Характерный компонент промоторов у кишечной палочки - т. н. бокс (последовательность) Прибнова:
(5')-ТАТААТ-(3')
(3')-АТАТТА-(5').
Он находится за 15 нуклеотидных пар (н. п.) от стартовой точки транскрипции. Общая же протяженность промотора - несколько десятков н. п.
В случае эукариот промотор - более сложное понятие, поскольку РНК-полимераза связывается с ДНК не непосредственно, а лишь вместе с комплексом других белков - т. н. общих факторов транскрипции.
В промоторе эукариот различают небольшую область инициации, ТАТА-бокс (сходный с боксом Прибнова у бактерий) и ряд других типичных участков.
Что касается участков связывания регуляторных белков, то у бактерий их называют операторами. Они располагаются после промотора (рис. 3). При определенных условиях с оператором связывается специфический белок-репрессор, и это блокирует «прочтение» РНК-полимеразой соответствующей группы генов. Еще одним регулятором работы гена может быть - аттенюатор – это выключатель - участок ДНК между оператором и структурными генами, на котором при определенных условиях прекращается транскрипция.
Рис. 3. Функциональные отделы в ДНК бактерий.
У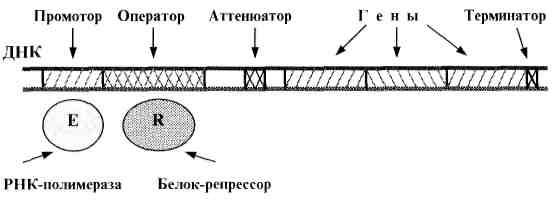 эукариот (включая человека) регуляцию
«прочтения» генов осуществляют не
только белки-репрессоры, но и
белки-активаторы - транскрипционные
факторы
(рис. 4).
эукариот (включая человека) регуляцию
«прочтения» генов осуществляют не
только белки-репрессоры, но и
белки-активаторы - транскрипционные
факторы
(рис. 4).
К последним относятся общие факторы транскрипции, необходимые для связывания РНК-полимеразы с промотором. Эти факторы имеются во всех клетках и необходимы для «прочтения» любого функционирующего гена.
Другие транскрипционные факторы повышают активность только определенных генов. Локусы ДНК, связывающие такие факторы, называются энхансерами.
Энхансеры могут располагаться достаточно далеко от регулируемого гена: на расстоянии нескольких тысяч нуклеотидных пар.
Рис. 4. Функциональные отделы в ДНК эукариот.
К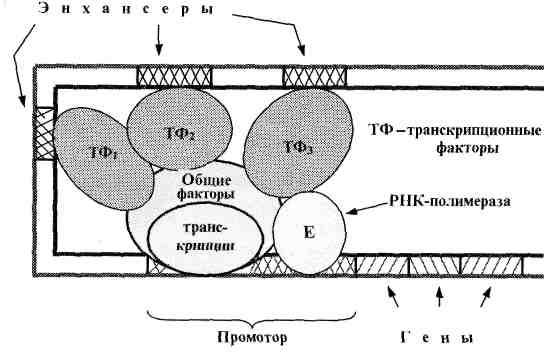 ак
же связывание с ними транскрипционных
факторов может стимулировать активность
гена?
ак
же связывание с ними транскрипционных
факторов может стимулировать активность
гена?
Видимо, дело в сложной пространственной организации ДНК. Она образует петли, благодаря чему энхансеры сближаются с промоторной зоной и связанные с ними факторы влияют на активность транскрипционного комплекса.
Причем, для некоторых ключевых генов в клетке имеется сразу несколько энхансеров, достаточно удаленных друг от друга. Следовательно, все они в результате изгибов ДНК должны собираться примерно в одном месте пространства.
Эта ситуация напоминает формирование активного центра в молекуле фермента. Там тоже аминокислотные остатки, образующие этот центр, расположены в разных участках полипептидной цепи и собираются вместе лишь при формировании третичной структуры.
Что же касается белков-репрессоров, то они при вхождении в состав транскрипционных комплексов не повышают, а понижают их активность.
Часто используется также термин «супрессоры» - так называются факторы (обычно это белки), которые угнетают тот или иной клеточный процесс. Например, опухолевые супрессоры - регуляторные белки, препятствующие опухолевому росту. Очевидно, по механизму своего действия они могут быть как репрессорами, так и транскрипционными факторами.
Наконец, в цепи ДНК после структурных генов могут содержаться короткие локусы - терминаторы, служащие сигналами об окончании (терминации) транскрипции ДНК.
Дополнительная литература.
-
Грин Н., Стаут У., Тейлор Д. Биология: В 3-х т. Т.3: Пер с англ. / Под ред. Р. Сопера. – М.: Мир, 1993. С.217-221.
-
Мушкамбаров Н.Н., Кузнецов С.Л. Молекулярная биология. М.: 2003 554 с.