


Глава 3
ПАТОГЕНЕЗ И ПАТОЛОГИЧЕСКАЯ АНАТОМИЯ ТУБЕРКУЛЕЗА
Вразвитии туберкулеза выделяют два периода — первичный
ивторичный. Необходимость такого деления обусловлена су щественными различиями в реакции организма человека на первый и повторный контакты с МБТ.
Первичный период туберкулезной инфекции начинается с мо мента первого внедрения в организм вирулентных МБТ. В большинстве случаев первичное инфицирование не приводит к заболеванию благодаря адекватной реакции защитных сис тем организма. При несостоятельности иммунной защиты, массивности и высокой вирулентности инфекции первичное экзогенное инфицирование вызывает развитие первичного ту беркулеза. Завершение первичного периода туберкулезной ин фекции связано с разрушением большей части МБТ и элими нацией из организма остатков микробных тел. Небольшая часть микобактериальной популяции инкапсулируется в оста точных изменениях. При благоприятном исходе первичного инфицирования, когда организму человека удается преодолеть бактериальную агрессию и заболевание не возникает, эти ос таточные морфологические изменения определяются лишь при микроскопическом исследовании. При клиническом ис следовании они недоступны для визуализации. В отличие от них остаточные посттуберкулезные изменения, образовавшие ся в процессе обратного развития первичного туберкулеза, бо лее значительны. Их можно выявить посредством рентгеноло гического исследования.
В течение первичного периода туберкулезной инфекции формируется противотуберкулезный иммунитет, обеспечи вающий повышение устойчивости организма к МБТ.
Клиническими формами первичного туберкулеза являются туберкулезная интоксикация, туберкулез внутригрудных лим фатических узлов и первичный туберкулезный комплекс.
Вторичный период туберкулезной инфекции (вторичный ту беркулез) развивается двумя путями. Один из них — повторное заражение МБТ человека, перенесшего первичный период ту беркулезной инфекции (экзогенная суперинфекция); другой — реактивация остаточных посттуберкулезных изменений, сфор мировавшихся в исходе первичного периода (эндогенная реак-
38
тивация). Обязательным условием развития вторичного тубер кулеза является снижение напряженности клеточного имму нитета, которое наступает под воздействием разных неблаго приятных факторов внешней и внутренней среды. При благо приятном течении вторичный период заканчивается форми рованием остаточных посттуберкулезных изменений, которые по своей морфологической структуре отличаются от остаточ ных изменений первичного генеза.
Клиническими формами вторичного туберкулеза, согласно принятой в России классификации, являются очаговый тубер кулез, инфильтративный, казеозная пневмония, туберкулема, кавернозный, фиброзно-кавернозный и цирротический тубер кулез.
Некоторые формы туберкулеза имеют своеобразное тече ние, которое нельзя в полной мере отнести к первичному или вторичному периоду туберкулезной инфекции. Туберкулез, имеющий такое течение, принято называть послепервичным. Он может развиваться при прогрессировании первичного ту беркулеза или в результате реактивации остаточных постту беркулезных изменений первичного генеза. К послепервич ным относят диссеминированные формы туберкулеза.
3.1.Первичное инфицирование микобактериями туберкулеза
искрытое течение туберкулезной инфекции
Первичное заражение человека МБТ обычно происходит аэрогенным путем. Другие пути проникновения микобакте рий — алиментарный, контактный и трансплацентарный — на блюдаются значительно реже.
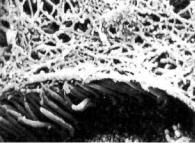
При аэрогенном заражении МБТ защитную роль играет система мукоцилиарного клиренса (рис. 3.1, 3.2). Слизь, вы
деляемая |
бокаловидными |
|
|||||
клетками слизистой оболочки |
|
||||||
бронхов, |
способствует |
склеи |
|
||||
ванию |
поступивших в |
дыха |
|
||||
тельные |
пути микобактерий. |
|
|||||
Их |
элиминацию |
обеспечива |
|
||||
ют |
синхронные |
движения |
|
||||
ресничек мерцательного эпи |
|
||||||
телия |
и |
волнообразные |
со |
|
|||
кращения мышечного |
слоя |
|
|||||
стенки |
|
главных |
бронхов |
и |
|
||
трахеи. |
|
Этот универсальный |
|
||||
механизм защиты может ока |
Рис. 3.1. Поверхность эпители |
||||||
заться |
весьма эффективным. |
альной выстилки слизистой обо |
|||||
В ряде случаев при эпизоди |
лочки трахеи в норме. Электро- |
||||||
ческом, |
кратковременном |
нограмма. х 16 400. |
|||||
39
контакте с бактериовыделителем он позволяет избежать инфицирования МБТ. При более длительном контакте здорового человека с источ ником инфекции мукоцилиарный клиренс способствует уменьшению числа микобак терий, проникающих в кон цевые отделы дыхательных путей. В результате, несмотря на происходящее инфициро вание, вероятность заболева ния туберкулезом снижается.
Нарушения мукоцилиарного клиренса, возникающие при остром или хроническом воспалении верхних дыха тельных путей, трахеи и круп
ных бронхов, а также при воздействии токсичных веществ, создают предпосылки для поступления МБТ в бронхиолы и альвеолы. В этих случаях вероятность аэрогенного инфициро вания МБТ и заболевания туберкулезом при прочих равных условиях значительно увеличивается.
При алиментарном пути заражения МБТ возможность и исход первичного инфицирования в значительной степени за висят от состояния кишечной стенки и всасывающей функ ции кишечника.
В зависимости от места внедрения МБТ первоначально мо гут проникать в легкое, миндалины, кишечник и другие орга ны и ткани. Поскольку возбудители туберкулеза не выделяют экзотоксин, а возможности для их фагоцитоза на этом этапе весьма ограничены, присутствие в тканях небольшого числа микобактерий обычно проявляется не сразу. Микобактерии находятся внеклеточно, медленно размножаются, а окружаю щая их ткань сохраняет нормальную структуру. Такое состоя ние определяется как латентный микробизм, при котором макроорганизм проявляет толерантность к МБТ. Независимо от начальной локализации микобактерии с током лимфы до вольно быстро попадают в регионарные лимфатические узлы, а затем лимфогематогенным путем распространяются по орга низму. Возникает первичная облигатная (обязательная) микобактериемия. Микобактерии оседают в органах с наиболее развитым микроциркуляторным руслом — в легких, лимфати ческих узлах, корковом слое почек, эпифизах и метафизах трубчатых костей, ампуллярно-фимбриональных отделах ма точных труб, увеальном тракте глаза.
МБТ, оседая в различных тканях, продолжают размножать-
40
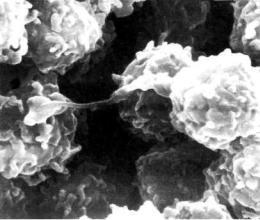
Рис. 3.3. Альвеолярные макрофаги. Электронограмма. х 6000.
ся. Популяция возбудителей туберкулеза может значительно увеличиться, прежде чем сформируется иммунитет и появится реальная возможность для их разрушения и элиминации.
В месте расположения микобактериальной популяции воз никает неспецифическая защитная реакция — фагоцитоз. Первыми фагоцитирующими клетками, которые пытаются поглотить и разрушить МБТ, являются полинуклеарные лейко циты. Однако их бактерицидный потенциал оказывается не достаточным для защитной функции. Полинуклеарные лейко циты, вступившие в контакт с МБТ, погибают.
Вслед за полинуклеарами с МБТ взаимодействуют макро фаги (рис. 3.3). Первая фаза такого взаимодействия состоит в фиксации МБТ на клеточной мембране макрофага специаль ными рецепторами. Следующая, вторая фаза направлена на поглощение МБТ. Участок плазмолеммы макрофага погружа ется в цитоплазму и образуется фагосома, содержащая МБТ. Третья, завершающая фаза связана с формированием фаголизосомы, которая возникает при слиянии фагосомы и лизосомы макрофага. В этих условиях протеолитические лизосомальные ферменты могут оказывать расщепляющее действие на поглощенные МБТ и разрушать их (рис. 3.4).
В большинстве случаев первичный контакт МБТ и макро фага происходит на фоне дисфункции лизосом фагоцитирую щей клетки. Появление этой дисфункции связано с повреж дающим воздействием на лизосомальные мембраны АТФ-по- ложительных протонов, сульфатидов и корд-фактора, которые синтезируются МБТ. Дисфункция лизосом препятствует обра зованию фаголизосомы, и лизосомальные ферменты не могут воздействовать на поглощенные микобактерии. В этих случаях
41
|
макрофаг |
|
становится |
||||
|
своеобразным |
контей |
|||||
|
нером |
для |
возбудителя |
||||
|
туберкулеза |
(рис. |
3.5). |
||||
|
Внутриклеточно |
распо |
|||||
|
ложенные МБТ продол |
||||||
|
жают |
расти, |
размно |
||||
|
жаться |
и |
инициировать |
||||
|
образование |
|
веществ, |
||||
|
которые оказывают по |
||||||
|
вреждающее |
|
действие |
||||
|
на |
клетку-хозяина. |
|||||
|
Макрофаг |
|
постепенно |
||||
|
погибает, |
и |
микобакте |
||||
|
рии вновь |
попадают в |
|||||
|
межклеточное |
|
про |
||||
|
странство. |
Такое |
взаи |
||||
|
модействие МБТ и мак |
||||||
Рис. 3.4. Взаимодействие МБТ и мак |
рофага |
называют |
неза |
||||
вершенным |
фагоцито |
||||||
рофага [по М. А. Пальцеву]. |
|||||||
зом. Дальнейшая |
судьба |
||||||
|
|||||||
|
микобактерий |
и |
исход |
||||
первичного инфицирования зависят от способности организ ма активировать макрофаги и создавать условия для завершен ного фагоцитоза.
В активации макрофагов и повышении устойчивости орга низма к действию МБТ ведущая роль принадлежит приоб ретенному клеточному иммунитету.
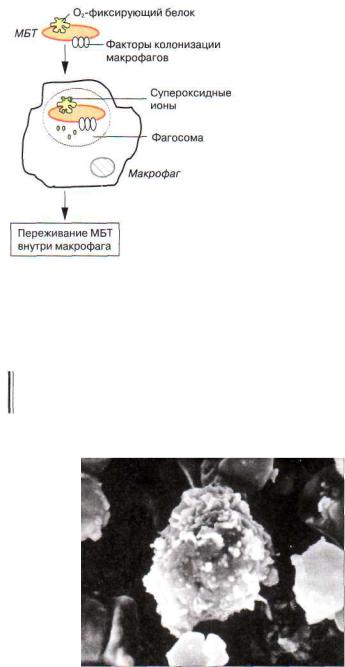
Рис. 3.5. Т-лимфоциты. Электронограмма. х 10 000.
42
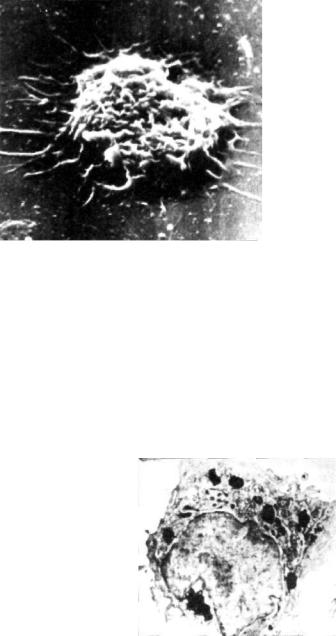
Рис. 3.6. Активированный макрофаг. Электронограмма. х 6000.
В основе приобретенного клеточного иммунитета лежит эффективное взаимодействие макрофагов и лимфоцитов. Особое значение приобретает контакт макрофагов с Т-хелпе- рами (CD4+) и Т-супрессорами (CD8+). Макрофаги, погло тившие МБТ, экспрессируют на своей поверхности антигены микобактериальных клеток в виде пептидов. Они также выде ляют в межклеточное пространство медиаторы, в частности интерлейкин-1 (ИЛ-1), которые активируют Т-лимфоциты (CD4+). В этих условиях Т-хелперы (CD4+) взаимодействуют с макрофагами и восприни мают информацию о генети ческой структуре возбудите ля. Сенсибилизированные
Т-лимфоциты (CD4+;
CD8+) выделяют медиато ры-лимфокины — хематаксины, гамма-интерферон, интерлейкин-2 (ИЛ-2), кото рые активируют миграцию макрофагов в зону располо жения МБТ, повышают фер ментативную и общую бак терицидную активность мак рофагов (рис. 3.6, 3.7).
Активированные макро фаги способны усиленно ге нерировать весьма агрессив-
Рис. 3.7. Активированный макро фаг. Трансмиссионная электро нограмма. х 14 000.
43
ные формы кислорода и перекись водорода, что сопровождается так на зываемым кислород ным взрывом, воздейст вующим на фагоцити руемый возбудитель ту беркулеза. Одновремен но с участием L-аргини- на и фактора некроза опухоли-альфа (ФНО-α) образуется оксид азота (NO), который также обусловливает выражен ный антимикобактериальный эффект. Под влиянием всех этих
факторов способность микобактерий препятствовать образо ванию фаголизосомы значительно ослабевает. Завершающая стадия фагоцитоза, направленная на переваривание возбуди теля, протекает благополучно, и МБТ подвергаются разру шающему действию лизосомальных ферментов (рис. 3.8).
При адекватном развитии иммунного ответа каждое после дующее поколение макрофагов, вступающее во взаимодейст вие с туберкулезным возбудителем, становится все более иммунокомпетентным. Высокий бактерицидный потенциал ак тивированных макрофагов обеспечивает возможность разру шения поглощенных МБТ и защиту человека от возбудителя туберкулеза.
Выделяемые макрофагами медиаторы активируют и В-лим фоциты, ответственные за синтез иммуноглобулинов. Однако накопление в крови имуноглобулинов практически не повы шает устойчивость организма к МБТ. Полезным можно счи тать лишь образование опсонизирующих антител, которые формируются к полисахаридным компонентам МБТ. Они об волакивают микобактерии и способствуют их склеиванию, об легчая последующий фагоцитоз.
При первичном инфицировании МБТ становление имму нитета происходит одновременно с медленным размножением микобактерий и развитием локальных воспалительных изме нений. Повышение ферментативной активности макрофагов и лимфоцитов приводит к дополнительному синтезу веществ, инициирующих повышение сосудистой проницаемости и раз витие воспалительной реакции. Такими веществами являются фактор роста, фактор переноса, кожно-реактивный фактор, ФНО-α, оксид азота. С их действием связывают появление у клеток повышенной чувствительности замедленного типа (ПЧЗТ) к антигенам МБТ. В месте локализации туберкулезно-
44
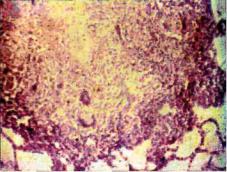
го возбудителя возникает специфическая клеточная реакция, способная ограничить распространение микобактерий. Под воздействием медиаторов иммунного ответа фагоцитирующие и иммунокомпетентные клетки устремляются к месту локали зации микобактерий. Макрофаги трансформируются в эпителиоидные клетки и гигантские многоядерные клетки Пирого- ва—Лангханса, участвующие в ограничении зоны воспаления. Образуется экссудативно-продуктивная или продуктивная ту беркулезная гранулема, которая по сути является морфологиче ским проявлением иммунной реакции организма на микобактериальную агрессию. Формирование гранулемы свидетельст вует о высокой иммунологической активности и способности организма локализовать туберкулезную инфекцию. Компакт ное расположение клеток гранулемы обеспечивает лучшие ус ловия для взаимодействия фагоцитирующих и иммунокомпетентных клеток. На высоте гранулематозной реакции в грану лемах преобладают Т-лимфоциты, присутствуют и В-лимфо циты. В гранулеме находится множество макрофагов, которые продолжают выполнять фагоцитарную, аффекторную и эффекторную функции в иммунном ответе. Эпителиоидные клетки менее способны к фагоцитозу, они активно осуществ ляют пиноцитоз и синтез гидролитических ферментов. В цен тре гранулемы может появиться небольшой участок казеозного некроза, который формируются из тел макрофагов, погиб ших при контакте с МБТ.
Реакция ПЧЗТ появляется через 2—3 нед после инфициро вания, а достаточно выраженный клеточный иммунитет формируется через 8 нед.
По мере развития иммунного ответа размножение мико бактерий замедляется, их общее число уменьшается, специ фическая воспалительная реакция затихает. Однако оконча тельной ликвидации возбудителя туберкулеза не происходит даже при полноценном взаимодействии макрофагов и Т-лим- фоцитов. Определенная популяция МБТ сохраняется в орга низме хозяина в виде живых, нередко биологически изменен ных особей (в частности, L-форм). Они локализуются в еди ничных туберкулезных гранулемах, окруженных плотной фиб розной капсулой. Сохранившиеся МБТ локализуются внутриклеточно и предотвращают формирование фаголизосомы, по этому для лизосомальных ферментов они недоступны. В связи с сохранением микобактерий противотуберкулезный иммуни тет называют нестерильным. Оставшиеся в организме МБТ поддерживают популяцию сенсибилизированных Т-лимфоци- тов и обеспечивают достаточную эффективность защитных иммунологических реакций. Инфицированный микобакте риями человек сохраняет их в своем организме весьма дли тельное время, иногда всю жизнь. При возникновении нару-
45