

Лекция №4. А.Т
..docxЛекция №4.
Тема: Организация наследственного материала (I).
1. Наследственность и изменчивость – фундаментальные свойства живого.
2. Эволюция понятия «ген».
3. Доказательства роли ДНК в передачи генетической информации.
4. Строение и функции нуклеиновых кислот.
5. Генетической код и его свойства.
6. Свойства и классификация генов.
1. Наследственность и изменчивость – это важнейшие свойства живого, которые не только отличают живое от неживого, но и определяют совместно с размножением бесконечное продолжение (непрерывность) жизни.
Из всех органических молекул способностью к саморепродукции обладают только нуклеиновые кислоты. Между тем, находясь в клетках, они контролируют их структуру и свойства (активность). Поэтому уникальность жизни в генетическом смысле заключается в том, что нуклеиновые кислоты через половые клетки обеспечивают химическую связь между поколениями. Благодаря размножению, наследственности и изменчивости жизнь видов продолжается бесконечно долго как непрерывное чередование поколений с сохранением между ними химических связей.
2. О единицах наследственности впервые написал Ч. Дарвин. Он назвал их наследственными факторами. В 1865 г. вышла работа Г. Менделя «Опыты над растительными гибридами». В ней он писал о наследственных задатках, которые родительские особи передают потомкам при половом размножении. Мендель проводил опыты на горохе. Он писал, что наследственные задатки находятся в половых клетках родителей, а при оплодотворении они соединяются в зиготе. В 1902 году Т. Бовери, Э.В. Вильсон и Д. Сеттон предположили, что наследственные факторы связаны с хромосомами. В 1906 г. У. Бэтсон ввел в биологию термин «генетика», а в 1909 г. В. Иогансен – термин «ген». В 1911 году Т. Морган с сотрудниками, проводя опыты на мухе дрозофиле, пришли к выводу, что гены расположены в хромосомах в линейном порядке, и сформулировали хромосомную теорию наследственности. Неясным оставался вопрос – что является веществом наследственности? 1928 г. Н.К. Кольцов предположил, что хромосома – это крупная белковая молекула, радикалы которой выполняют функции генов.
3. В соответствии с современными представлениями генетическим материалом являются нуклеиновые кислоты.
Нуклеиновые кислоты были обнаружены в ядрах клеток гноя в 1869 г. швейцарским физиологом Фридрихом Мишером. Это открытие является настолько важным, что заслуживает приведения цитаты из работы Ф. Мишера, в которой он описывал свои опыты. «Обрабатывая клетки гноя слабым щелочным раствором, я получил в результате нейтрализации раствора осадок, который не растворялся ни в воде, ни в уксусной кислоте, ни в разведенной соляной кислоте, ни в обычном солевом растворе и который не мог принадлежать ни к одному из белков, известных в настоящее время». Обнаруженное вещество Ф. Мишер назвал нуклеином. Как считают, он не мог не знать, что открыл ДНК и что оказался в начале исследований ДНК.
Важнейшим объектам современных исследований молекулярной генетики являются бактериальные вирусы – фаги. Открытие бактериофагов связано с именем французского

Рис. 1. Жизненный цикл вирулентного фага, например Т2 или Т4.
исследователя д′Эрелля (1917), обнаружившего лизис бактериальной культуры под влиянием фильтрата испражнений больных людей. Присутствующий в фильтратах агент был назван бактериофагом (т.е. «пожирателем бактерий»). Все фаги могут быть разделены на два класса – вирулентные и умеренные. Фаг, способный вызвать лизис зараженной бактерии, называют вирулентным. Умеренные фаги, проникая в клетку, не разрушают ее,

Рис. 2. Жизненный цикл умеренного фага, например λ.
а остаются в ней в качестве «жильцов» - симбионтов. При этом умеренный фаг пребывает в ней в форме профага, который размножается синхронно с делением самой клетки. Профаг, таким образом, представляет собой невирулентное состояние фага.
Присутствие профага в бактерии делает ее иммунной по отношению к заражению частицами генетически родственного фага. К частицам фага другой генетической природы бактерия не имеет иммунитета, и она может быть ими заражена. Поэтому в клетку могут проникать частицы фагов разной генетической природы.
Нормально размножающаяся бактерия с находящимся в ней профагом называется лизогенной бактерией. Явление симбиотических отношений фага с бактерией называют лизогенией. Отношение бактериальных клеток к фагам, лизирование бактерии, или лизогенизация ее, определяются генотипом бактерии и фага.
Строение фагов в настоящее время хорошо изучено. Они состоят из следующих структур:
1) сердцевины – генетического материала, представленного либо ДНК, либо РНК; ДНК или РНК может быть одноцепочечной или двухцепочечной;
2) капсида – защитной белковой оболочки, окружающей сердцевину;
3) нуклеокапсида – сложной структуры, образованной сердцевиной и капсидом;
4) оболочки – у некоторых вирусов, таких как ВИЧ и вирусы гриппа, имеется дополнительный липопротеиновый слой, происходящий из плазматической мембраны клетки-хозяина;
5) капсомеров – идентичных повторяющихся субъединиц, из которых часто бывают построены капсиды.
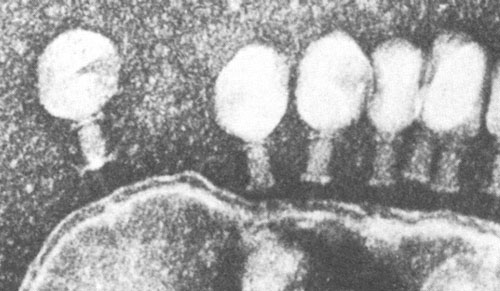 А.
А.
 Б.
Б.
Рис.3. А. Электронная микрофотография бактериофага, полученная методом негативного контрастирования. Б. Строение бактериофага Т2.
Изучение бактерий открыло целый ряд явлений, осветивших с новой стороны источники наследственной изменчивости и механизмы наследственной передачи. Описаны три основных механизма обмена генетическим материалом у бактерий: трансформация, трансдукция и конъюгация.
Трансформация – это перенос генетического материала от одного организма к другому. Посредством генетической рекомбинации часть трансформирующей молекулы ДНК может обмениваться с частью хромосомной ДНК донора. Явление трансформации было обнаружено в опытах английского микробиолога Ф.Гриффитса (1928 г.), работавшего с двумя штаммами пневмококка. У этих микроорганизмов обычно бывает два вида колоний: гладкие (S) и шероховатые (R). Крупные, гладкие колонии образуются бактериями, которые имеют полисахаридную капсулу, защищающую их от фагоцитоза, и из-за этого обладают вирулентностью. Бактерии, образующие мелкие, шероховатые колонии, не имеют такой капсулы и лишены вирулентности. В опытах Гриффита при введении мышам смеси живых клеток R и убитых нагреванием клеток S животные заболевали; из их органов можно было выделить клетки S, образующие при любом количестве пересевов такие же гладкие колонии. Таким образом, было показано, что клетки бактерий, обладающие определенным признаком, под влиянием какого-то вещества, содержащегося в клетках с противоположным признаком, преобразовывались, трансформировались. Это изменение свойств передавалось из поколения в поколение.
Введение мышам:
1. авирулентного штамма пневмококков (R). |
мыши живут |
2. вирулентного штамма пневмококков (S). |
мыши гибнут |
3. вирулентного, убитого нагреванием 65º, штамма пневмококков |
мыши живут |
4. вирулентного, убитого нагреванием, + авирулентной живой штаммы |
мыши гибнут |
Схема опытов Ф. Гриффитса.
Природа трансформирующего вещества оставалась невыясненной вплоть до 1944 г., когда О. Эвери с сотрудниками удалось выяснить природу этого загадочного явления. Они взяли те же два штамма – R и S. Перед началом решающих опытов было изучено спонтанное мутирование обеих форм. Оказалось, что гладкая S-форма хотя и очень редко, но спонтанно мутирует в R-форму, а R-форма практически вовсе не мутирует в S-форму, т.е. мутации происходят почти исключительно в одном направлении: S→R. Стало очевидным, что признак одного штамма (S) через какое-то вещество экстракта передавался другому штамму (R), т.е. возникало направленное наследственное изменение.

Рис. 4. Схема опыта, демонстрирующего явление трансформации.
Далее была произведена тщательная очистка – выделение этого вещества из экстракта клеток S-формы. Вещество было названо трансформирующим фактором (ТФ), а само явление – трансформацией.
Явление трансформации стало одним из основных доказательств роли ДНК как носителя наследственной информации.
Изучение явления трансформации послужило толчком к открытию другого явления – трансдукции – переноса и рекомбинации генов у бактерий. Трансдукцией называется перенос ДНК из одной клетки (донора) в другую (реципиент) с помощью бактериофагов и передавать соответствующие свойства. Этот способ генетического обмена был открыт в 1952 г. Н. Зиндером и Дж. Ледербергом в отношении штаммов Salmonella и фага Р22. С тех пор возможность трансдукции была подтверждена на различных бактериях и фагах. Опыт, позволивший открыть этот новый генетический механизм и новый способ изучения наследственности, заключается в следующем. U-образная трубка в нижней части была разделена посредине бактериальным фильтром. В одну половину этой трубки были помещены тифозные бактерии (Salmonella typhimurium) штамма 22А, а в другую половину трубки – штамма 2А. При этом бактериальные клетки не могли переходить сквозь перегородку.
Штамм 22А нес мутацию, блокирующую синтез триптофана Т-, и поэтому при культивировании бактерии нуждались в добавки триптофана в среду. Штамм бактерии 2А имел мутацию, блокирующую синтез гистидина Н-, и поэтому нуждался в нем при культивировании.
После инкубации этих двух разных штаммов в трубке, разделенной только бактериальным фильтром, был произведен рассев клеток обоих штаммов. При рассеве клеток штамма 22А на среде, лишенной триптофана, было обнаружено небольшое число колоний. Следовательно, некоторые клетки штамма 22А каким-то образом приобрели способность синтезировать триптофан и смогли дать колонии на среде без этой аминокислоты.
Можно было предположить, что эти измененные клетки появились или в результате обратной мутации от Т- к Т+ или перехода трансформирующего фактора от штамма 2А. Но штамм 22А отличался высокой стабильностью, и поэтому указанную частоту появления (10-5) клеток генотипа Т+ нельзя было объяснить возникновением обратных мутаций. Трансформирующий фактор в среде также не был обнаружен. Фильтрующимся агентом, переносящим ген Т+ от штамма 2А к штамму 22А, оказался бактериофаг.
2А 22А

Бактериальный фильтр
Рис. 5. Схема опыта, демонстрирующего явление трансдукции.
Таковы первые факты, доказавшие передачу наследственной информации с помощью бактериофага от бактерии одного генотипа к бактерии другого генотипа.
Таким образом, трансдукция так же, как и трансформация, является своеобразным процессом рекомбинации генов. Рекомбинация генов является одним из механизмов, осуществляющих у бактерий комбинативную изменчивость, которая у высших организмов обеспечивается мейозом.
С помощью трансформации и трансдукции осуществляется односторонний обмен наследственными факторами между бактериями. И эти процессы в какой-то мере компенсируют отсутствие у них настоящего полового процесса.
Поиски полового процесса у бактерий в течение длительного времени были безуспешными. Лишь после того, как были разработаны методы селективных сред и получены штаммы биохимических мутантов, Дж. Ледербергу и Е. Татуму удалось в 1946 г. доказать наличие своеобразного полового процесса у Escherichia coli на примере штамма К12. Процесс переноса генетической информации от одной бактерии к другой при контакте клеток получил название конъюгации.
В опытах Дж. Ледерберга и Э. Татума (1946 г.) были взяты два штамма, различающиеся по генотипу: В-М-Р+Т+ и В+М+Р-Т-. Клетки обоих ауксотрофных штаммов в течение некоторого времени выращивали в смешанной культуре, а затем высевали на минимальную среду. Ни один из двух исходных штаммов не мог расти на этой среде. Однако на каждые 109 посеянных клеток из смешанной культуры на минимальной среде вырастало около 100 колоний. По генотипу эти клетки могли быть только В+М+Р+Т+.
Чтобы установить, нужен ли для процесса передачи информации физический контакт клеток, Б. Дэйвис поместил оба штамма в U-образную трубку, плечи которой были разделены бактериальным фильтром. После нескольких часов инкубации он высеял клетки на минимальную среду. Колонии на ней не выросли. Отсюда следовал вывод о том, что для появления рекомбинантов необходим непосредственный контакт между бактериальными клетками. Это заключение было сделано на основе чисто генетических экспериментов. Позднее с помощью электронной микроскопии были получены фотографии конъюгирующих бактерий, соединенных друг с другом тонким мостиком-пилем. Эти данные свидетельствовали о том, что E. сoli имеет определенный тип

Рис. 6. Схема опыта, демонстрирующего явление конъюгации.
скрещивания, называемый конъюгацией, когда генетический материал может передаваться между клетками, временно находящимися в контакте.
4. Материалом, из которого состоят гены, является дезоксирибонуклеиновая кислота (ДНК), а материалом, обеспечивающим декодирование генетической информации, является РНК. Таким образом, нуклеиновые кислоты – это хранители (носители) и переносчики генетической информации. Структура молекулы ДНК была расшифрована в 1953 г. Дж. Уотсоном, Ф. Криком и М. Уилкинсом. Они назвали ее «нить жизни».
Нуклеиновые кислоты является полимерами. Их мономеры – нуклеотиды. Нуклеотид содержит азотистое основание, сахар дезоксирибозу или рибозу и остаток фосфорной кислоты. Азотистых оснований 5 типов: аденин, гуанин, цитозин, тимин, урацил. Нуклеотиды ДНК содержат аденин, гуанин, цитозин, тимин. Нуклеотиды РНК содержат аденин, гуанин, цитозин, урацил. Азотистые основания обозначаются первыми буквами: А, Г – пуриновые; Т, Ц, У – пиримидиновые.
Молекула ДНК состоит из двух спиралей. Цепочка нуклеотидов соединяется ковалентными фосфодиэфирными связями между дезоксирибозой одного одного и остатком фосфорной кислоты другого нуклеотида. Внутри спирали находятся соединенные по принципу комплементарности (взаимодополняемости) азотистые основания: А – Т – две водородные связи Г – Ц – три водородные связи.
Свойство комплементарности азотистых оснований выражается в правилах Чаргаффа: - количество аденина равно количеству тимина (А=Т), количество гуанина равно количеству цитозина (Г=Ц).
ДНК находится в клетке в ядре, в митохондриях и пластидах.
Классификация нуклеиновых кислот.
Рис. 1.

Свойства ДНК: репликация (самовоспроизведение) и способность к репарации (восстановление структуры после нарушений молекулы).
Функция ДНК: сохраняет и передает генетическую информацию при размножении клеток и организмов.


Рис. 2. т-РНК.
Молекула РНК также является полинуклеотидом, но имеет одну цепочку. Вместо тимина в ее состав входит урацил, а вместо дезоксирибозы сахар рибоза.
У некоторых вирусов РНК является хранителем наследственной информации и имеет в молекуле 2 цепочки.
В клетке имеются три вида РНК. 3 – 4% от всей РНК составляет информационная РНК (и-РНК): она «переписывает» генетическую информацию с ДНК и переносит ее в рибосомы – место сборки белковых молекул. Рибосомальная РНК (р-РНК) составляет 80 -85 % от всей РНК. Она входит в состав рибосом и обеспечивает пространственное взаиморасположение и-РНК и т-РНК. Транспортная РНК (т-РНК) транспортирует (переносит) аминокислоты из цитоплазмы в рибосомы. т-РНК составляет 10-20% от всей РНК.
Рибонуклеиновые кислоты находятся в ядре, в цитоплазме, в митохондриях и пластидах.
Функции РНК: участие в синтезе белковых молекул (молекул полипептидов).
Формулируя свою модель, Д. Уотсон и Ф. Крик предположили, что репликация ДНК происходит в несколько последовательных этапов, а именно: а) разрыв водородных связей между двумя полинуклеотидными цепями и разделение последних; б) разматывание полинуклеотидных цепей; в) синтез вдоль каждой из полинуклеотидных цепей новой цепи с комплементарной последовательностью азотистых оснований. Они предположили далее, что разделение и разматывание полинуклеотидных цепей начинается с одного конца молекулы, продолжается по направлению к другому ее концу и сопровождается одновременно идущим с того же конца молекулы синтезом новых полинуклеотидных цепей.
Репликация ДНК – протекает в S – период интерфазы на матрице ДНК в направлении 5' - 3', одновременно на обеих цепях. Одна цепь («лидирующая») синтезируется по принципу комплементарности и опережает по времени другую. Образование второй цепи («отстающей») осуществляется фрагментами по 150 – 200 нуклеотидов (фрагменти Оказаки).
Для начала синтеза необходимы короткие последовательности РНП (10 - 200), играющие роль затравок (праймеров). Основной фермент синтеза – ДНК – α – полимераза (А.Корнберг, 1956 г.).
При репликации двухцепочечная ДНК делиться на индивидуальные цепи, каждая из которых выполняет роль матрицы. Параллельно с разделеннем цепей происходит раскручивание спирали, которое сопровождается скручиванием вновь синтезируемых цепей. Завершает синтез ДНК – лигаза, которая соединяет вместе фрагменты Оказаки и всю молекулу ДНК.
Репликация начинается в нескольких местах молекулы ДНК. Участок ДНК от точки начала одной репликации до точки начала другой, называется репликоном. Нуклеоид бактерий содержит 1 репликон, хромосомы эукариот – много репликонов.
Репликон имеет контролирующее элементы: точка начала (инициируется репликация), точка окончания (репликация останавливается). Точка, в которой происходит репликация - репликационная вилка. Она движется вдоль молекулы ДНК от точки начала.
В каждой вилке синтез новых цепей ДНК ведут одновременно несколько ДНК – полимераз, движующихся в противоположных направлениях. Это антипараллельный и прерывистый принцип синтеза.
Новая молекула ДНК состоит из 1 «старой» цепочки и 1 новой (полуконсервативный синтез). Весь геном клетки реплицируется 1 раз в S – периоде интерфазы митотического цикла. Репликация ДНК обеспечивает преемственность генетического материала («дочерние» молекулы идентичны «материнским» молекулам ДНК).

Рис.3.Схема удвоения ДНК.
5. Информация о структуре белков «записана» в ДНК в виде последовательности нуклеотидов. В процессе транскрипции она переписывается на синтезирующуюся молекулу и-РНК, которая выступает в качестве матрицы в процессе биосинтеза белка. Определенному сочетанию нуклеотидов ДНК, а следовательно, и и-РНК соответствует определенная аминокислота в полипептидной цепи белка.
Запись генетической информации в виде последовательности нуклеотидов в молекулах ДНК и и-РНК, называется генетическим кодом. Триплет нуклеотидов, кодирующий определенную аминокислоту, называется кодоном. Кодон – это элементарная функциональная единица гена.
Свойства генетического кода:
- триплетность: одной аминокислоте в молекуле полипептида соответствует один кодон;
- универсальность: у всех живых организмов один и тот же кодон определяет одинаковые аминокислоты;
- неперекрываемость: один нуклеотид входит в состав только одного триплета;
- вырожденность, или избыточность: одну аминокислоту может кодировать несколько триплетов (аминокислот – 20, возможных триплетов - 64);
- непрерывность (нет разделительных знаков между нуклеотидами);
- однонаправленность (образование и-РНК происходит в направлении от 3' конца к 5' концу).
- наличие среди триплетов инициирующих кодонов (с них начинается биосинтез белка), кодонов – терминаторов (обозначают конец биосинтеза белка).
Соответствие порядка нуклеотидов в молекуле ДНК порядку аминокислот в молекуле полипептида называется колинеарностью.
Табл. 1.

6. Свойства гена:
• специфичность (структурный ген детерминирует синтез данного полипептида);
• целостность (при программировании синтеза полипептида ген – неделимая единица);
• дискретность (наличие субъединиц);
• стабильность (редко изменяются);
• лабильность (способность мутировать);
• плейотропия (детерминирует развитие нескольких признаков);
• экспрессивность (степень фенотипического проявления);
• пенетрантность (частота фенотипического проявления гена).
Классификация генов по функциям:
1. Структурные гены несут информацию о различных видах РНК, ферментах и белках-гистонах.
2. Функциональные гены: гены модуляторы, усиливающие или ослабляющие действие структурных генов (ингибиторы, интеграторы, модификаторы) и гены-регуляторы, контролирующие работу структурных генов (регуляторы и операторы).
Классификация последовательностей ДНК:
1. Уникальные (1 геноме) – входят в состав структурных генов и детерминируют структуру полипептидов.
2. Повторяющиеся (десятки, сотни, миллионы раз) – промоторы, регулируют репликацию ДНК, участвуют в кроссинговере и др.
3. Транспозоны (прыгающие гены) – мобильные генетические элементы, способные встраиваться в хромосому, перемещаться вдоль нее, регулировать процессы обмена веществ, создавать устойчивость к антибиотикам.
По месту действия гены подразделяют на:
а) функционирующие во всех клетках (гены, кодирующие ферменты энергетического обмена):
б) функционирующие в клетках одной ткани (гены, детерминирующие синтез миозина в мышечной ткани):
в) специфичные для одного типа клеток (гены гемоглобина в незрелых эритроцитах).
Генотип соматических клеток одинаков, но клетки разных тканей отличаются. В различных клетках работают разные гены. Область проявления действия гена – поле действия гена (гены детерминирующие развитие определенных дерматоглифических показателей на пальцах, ладонях и стопах). Гены функционируют непостоянно (гены, детерминирующие синтез половых гормонов, работают с момента полового созревания, а к старости их функция снижается). Время работы гена – период его функционирования.
Гены выполняют в клетке две основные функции.
а) Гетеросинтетическая функция – это программирование биосинтеза белка в клетке.
б) Аутосинтетическая функция – репликация ДНК (самоудвоение ДНК).