

4.1. Основные представления о строении
и функционировании иммунной системы
Распознавание агрессивных агентов при иммунной защите не может основываться на регистрации их вредного воздействия на организм (такая защита была бы запоздалой). В процессе эволюции реализовалась другая стратегия, которая основывается на том, что агрессивные агенты чужеродны для организма. Это относится как к возбудителям инфекций так и к опухолевым клеткам, которые приобретают черты чужеродносги вследствие мутаций. Поэтому в основе иммунного распознавания лежит выявление в составе клеток, присутствующих во внутренней среде организма, маркеров чужеродности — субстанций, обозначаемых как антигены.
Иммунная защита включает триаду процессов
распознавание чужеродных агентов;
удаление их из организма;
запоминание, обусловливающее ускоренную реакцию на повторное воздействие тех же агентов
Эти процессы в своей совокупности образуют иммунный ответ Огромный объем сведений о строении и функционировании иммунной системы может быть представлен здесь лишь в максимально схематизированной форме
Структурная организация иммунной системы
Клетки иммунной системы можно разделить на антигенраспознаю- щие, вспомогательные и стромальные.
Антигенраспознающими клетками являются лимфоциты, 1 которые несут на своей поверхности рецепторы, предназна- | ченные для связывания антигенов.
Лимфоциты обусловливают специфический компонент иммунной защиты, направленный против конкретного антигена. К вспомогательным клеткам относят макрофаги, сегментоядерные лейкоциты, дендритные, тучные и другие клетки, которые осуществляют различные функции — подготавливают антигены к их распознаванию лимфоцитами, осуществляют фагоцитоз, секретируют активные субстанции и т д Стромальные клетки служат каркасом лимфоидных органов и формируют микроокружение, обеспечивающее функционирование других клеток иммунной системы.
Органы иммунной системы (лимфоидные органы) разделяются на центральные (вилочковая железа, у птиц — сумка Фабриция), в которых происходит развитие клеток иммунной системы, и периферические (лимфатические узлы, селезенка, лимфоидные образования пищеварительного тракта), где эти клетки реализуют свои функции (схема 4 1) В зависимости от того, в каких центральных органах происходит развитие лимфоцитов, их разделяют на тимусзависимые (Т-клетки) и бурсазави- симые (В-клетки; у млекопитающих В-клетки развиваются в костном мозге) Развитие лимфоцитов контролируется клетками стромы и их продуктами, развитие Т-клеток — эпителием вилочковой железы и вырабатываемыми им пептидными гормонами В периферических лимфоидных органах есть структуры, в которых преимущественно локализуются В- или Т-клетки, и отделы, в которых эти клетки соседствуют друг с другом.
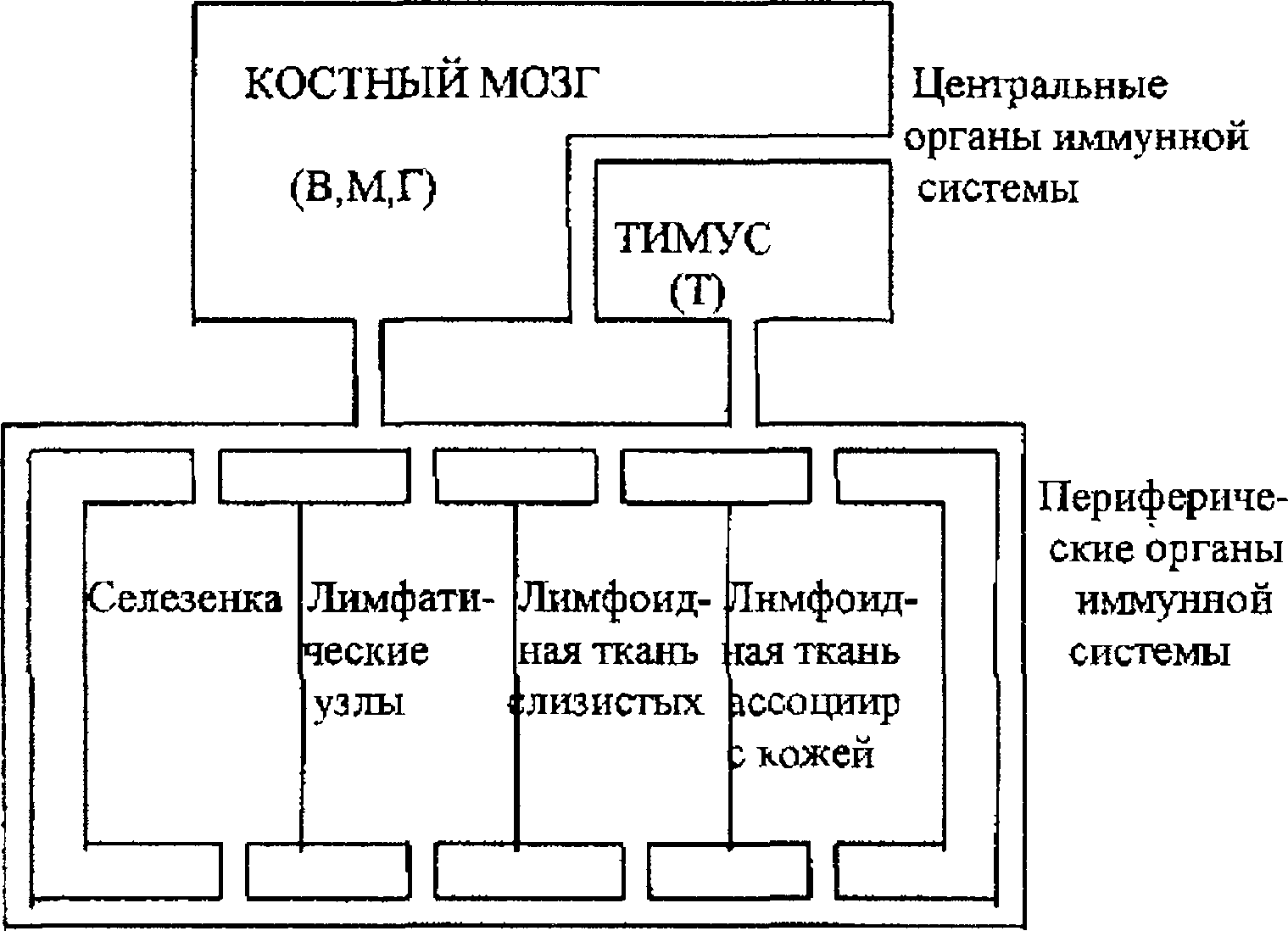
Пути рециркуляции лимфоцитов
Схема 4 1. Структура иммунной системы. Отражены взаимоотношения центральных и периферических органов иммунной системы, направление миграции лимфоцитов (указано стрелками) в процессе созревания и рециркуляции.
В — В-лимфоциты; Т — Т-лимфоциты, М — моноциты, Г — гранулоциты
3. Лимфоциты непрерывно рециркулируют, т.е перемещаются из органов в лимфо- и кровоток и возвращаются обратно. При этом сохраняется избирательность их поступления в органы иммунной системы, хотя не обязательно именно в те органы, из которых они вышли. Это сродство обеспечивается благодаря распознаванию рецепторами лимфоцитов молекул адгезии, характерных для лимфоидныхорганов. Существует два относительно автономных круга рециркуляции. Один из них включает лимфатические узлы и селезенку, второй — лимфоидные скопления и структуры слизистых оболочек пищеварительного, дыхательного, уроге- нитального и другихтрактов, т.е. лимфоидную ткань слизистых оболочек. В организме есть участки (иммунологически привилегированные зоны), недоступные для рециркулирующих лимфоцитов.
Основы иммунного распознавания
1. Антигены — это вещества (чаще всего белки или полисахариды), генетически чужеродные для конкретного организма и способные вызвать в нем иммунный ответ — реакцию, направленную на удаление антигена из внутренней среды организма. Основные свойства антигенов — специфичность, иммуногенность и чужеродность. Специфичность антигенов определяется небольшими химическими группами, обладающими химической индивидуальностью. Их называют антигенными детерминантами, или эпитопами. Обычно эти структуры отсутствуют в реагирующем организме; это и означает, что антиген для него чужероден. Именно эпитопы распознаются специфическими рецепторами лимфоцитов. Для обеспечения иммунного ответа на антиген, помимо присутствия в его молекуле эпитопов, необходимы дополнительные свойства, обеспечивающие им- муногенность молекулы (наличие нескольких эпитопов, гибкость и стабильность структуры, некоторые особенности метаболизма и т.д.). Для обозначения антигенов, к которым в организме формируется повышенная чувствительность, т.е. развивается иммунный ответ аллергического типа, используют термин аллерген. Аллерген может не обладать всеми атрибутами антигенов. Так, нередко аллергенами служат низкомолекулярные соединения, которые определяют специфичность аллергической реакции, но лишены иммуногенности; они приобретают ее, соединяясь с белками организма.
Каждый лимфоцит несет на своей поверхности рецепторы, предназначенные для распознавания антигена [антигенраспознающие рецепторы). Они идентичны по специфичности (т.е. распознают один и тот же эпитоп) и другим свойствам. Клетки, несущие одинаковые рецепторы, происходят от одного предшественника и образуют клон.
Важнейшими молекулами, предназначенными для распознавания и связывания антигенов (точнее их эпитопов), являются иммуноглобулины. Они существуют в двух формах — связанной с мембранами В-лимфоцитов и свободной растворимой форме. В первом случае они выполняют роль антигенраспознающих рецепторов В-клеток, во втором — функцию антител. Антитела секретируются плазматическими клетками, которые образуются в результате дифференцировки В-лимфоцитов. Иммуноглобулины — рецепторы и антитела — способны связывать и, следовательно, распознавать антигены как в растворимом состоянии, так и в форме, связанной с клеточной мембраной. Иммуноглобулины состоят из тяжелых и легких полипептидных цепей (обычно две пары), N-концевые части которых формируют антигенсвязывающий участок, а С-концевая (Fc) часть определяет разновидности иммуноглобулинов — изотипы (IgM, IgG, IgA, IgD, IgE).
Распознавание антигена T-лимфоцитами обеспечивается благодаря присутствию на их поверхности рецепторов, в составе которых содержатся димеры (ар или уб), по своей структуре родственные, но не идентичные иммуноглобулинам. Они распознают не свободный антиген, а его пептидные фрагменты, встроенные в аутологичные молекулы главного комплекса гистосовместимости (МНС — от англ. «Major histocompatibility complex»). Пептидные фрагменты могут формироваться при расщеплении внутриклеточных антигенов или при поглощении клеткой экзогенных молекул. В первом случае их фрагменты встраиваются в молекулы МНС I класса, во втором — в молекулы МНС II класса. Молекулы МНС-1 содержатся на поверхности всех ядросодержащих клеток, а молекулы МНС-И — только на клетках, специализированныхдля «презентации», т.е. представления антигенов Т-лимфоцитам. Эти клетки называют антигенпрезенти- рующими; к ним относят дендритные клетки, макрофаги, В-лимфоциты.
В особых случаях, иногда связанных с патологией, функцию презентации антигена приобретают другие клетки, в частности эпителиальные.
В распознавании комплекса молекул МНС и антигенных пептидов участвуют вспомогательные молекулы — корецепторы CD4 и CD8 (молекулы, расположенные на поверхности клеток иммунной системы, обычно обозначают в соответствии с международной классификацией символами CD с соответствующим номером; эти молекулы выявляют с помощью моноклональных антител, объединенных в группы, кластеры, и CD — это сокращение слов «cluster designation»). CD8 обладают сродством к молекулам МНС-1 и участвуют в распознавании пептидов, находящихся в составе этих молекул; CD4 обладают сродством к молекулам MHC-II и облегчают распознавание связанных с ними пептидов. Зрелые Т-клетки экспрессируют лишь один тип корецепторов. Т-клетки, несущие на своей поверхности CD8, выполняют функции цитотоксических Т-лимфоцитов (киллеров), а С04+-клетки — функции клеток-помощников (хелперов).
Специфичность рецепторов лимфоцитов формируется в процессе их дифференцировки в результате комбинации и видоизменения нескольких генетических участков, ответственных за структуру вариабельной части молекул иммуноглобулинов или рецепторов Т-клеток. В конечном счете в клетке создается «зрелый V-ген», который и определяет специфичность антигенраспознающего рецептора лимфоцита. В каждой клетке этот ген индивидуален, и специфичность рецептора уникальна. Совокупность рецепторов, экспрессированных на всех лимфоцитах, формирует «первичный антигенраспознающийрепертуар». Он содержит наряду с необходимыми для защиты организма ненужные и потенциально аутоагрессивные специфические структуры. Поэтому он подвергается коррекции путем поддержания необходимых для организма клонов лимфоцитов (положительная селекция) и удаления лишних и вредных клонов (отрицательная селекция). Особенно важной является селекция клонов Т-лимфоцитов, которая происходит в вилочковой железе. В результате формируется окончательный антигенраспознающий репертуар Т-клеток, включающий клоны, которые распознают чужеродный пептид в составе аутологичной молекулы МНС («измененное свое»). Нарушение процессов селекции клонов лимфоцитов может служить основой аутоиммунных процессов.
Иммунный ответ
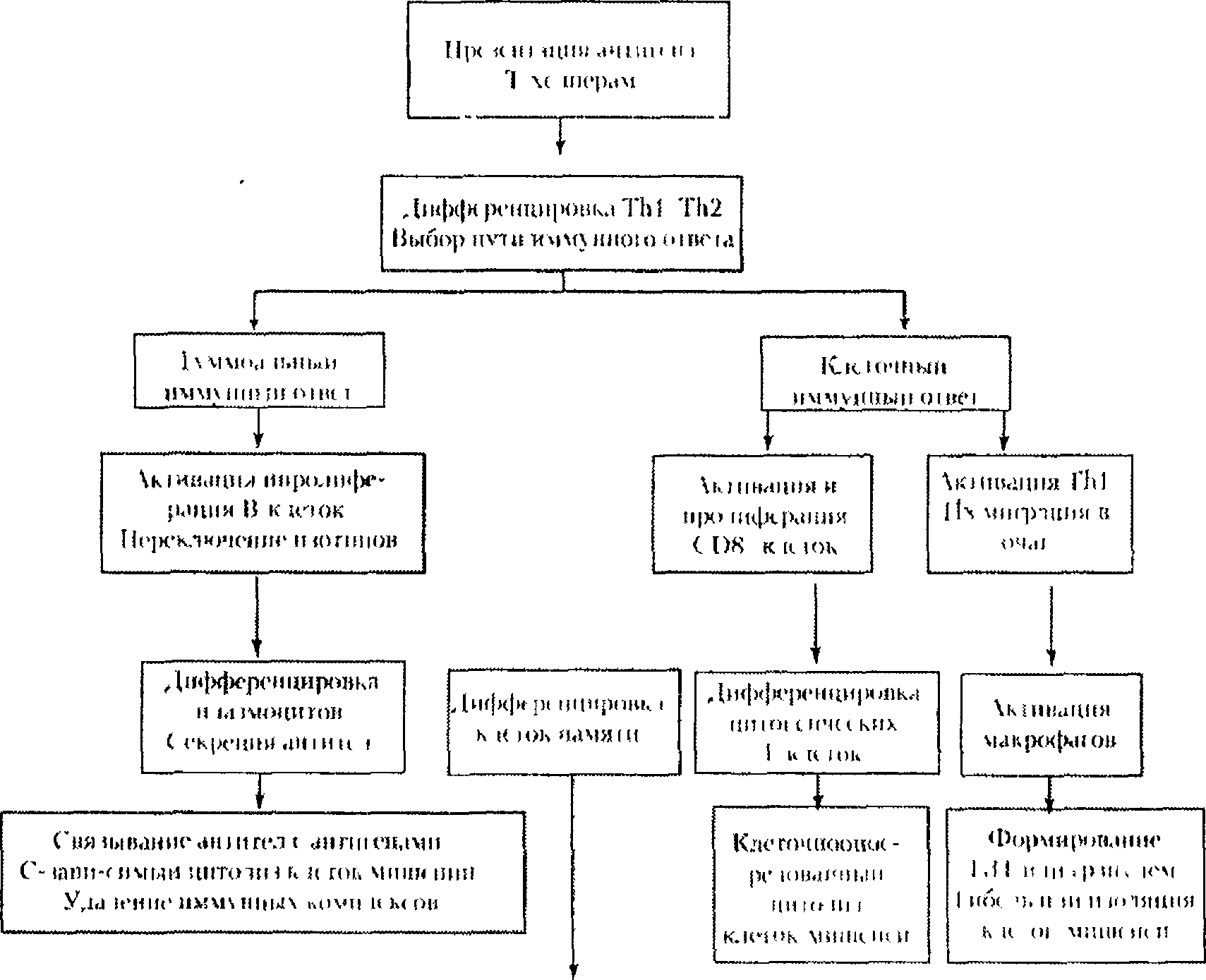
1. Иммунным ответом называют специфическую реакцию, направленную на удаление конкретных чужеродных субстанций (антигенов) из внутренней среды организма и формирование иммунологической памяти об этих антигенах. Иммунный ответ имеет две взаимосвязанные составляющие, основой которых служат механизмы естественной резистентности и специфической иммунной реактивности (схема 4.2.)
.
N ( коржики pa шигиг
«UvpHMlUH О ОНК'ТЛ
Схема 4.2. Развитие иммунного ответа.
Филогенетически более древние механизмы естественной резистентности связаны с воспалительной реакцией. Они включают примитивные механизмы обнаружения чужеродных субстанций, в частности, основанные на узнавании белками-лектинами определенных углеводных групп, которые несвойственны данному организму или присутствуют на его клетках в замаскированной форме (лектиновое распознавание).
Основным результатом действия факторов естественной резистентности является цитолиз — гибель и расщепление чужеродных клеток. Существует три типа механизмов цитолиза: внеклеточный (обусловленный секретируемыми продуктами), внутриклеточный (осуществляемый путем фагоцитоза) и контактный (его основой служат сигналы, возникающие при межклеточных контактах, которые включают активную, т.е. требующую затрат энергии и участия ферментов форму гибели клеток-мишеней — апоптоз). Внеклеточный цитолиз обычно осуществляют эозинофилы, внутриклеточный — нейтрофилы и макрофаги, контактный — естественные киллеры.
Наиболее эффективной формой цитолиза является внутриклеточный цитолиз, осуществляемый по механизму фагоцитоза, т.е. поглощения объекта клеткой. Это связано с тем, что внутриклеточная среда содержит наиболее полный набор факторов, способных вызвать гибель и расщепление клеток. Важными предпосылками фагоцитоза являютсяхе- мотаксис (направленное движение фагоцита в сторону объекта, обусловленное химическими веществами, которые он выделяет) и распознавание объекта фагоцитоза мембранными рецепторами фагоцитов. Последнее облегчается при опсонизации клеток — связывании с их поверхностью антител или компонентов комплемента, которые распознаются рецепторами фагоцитов. При поглощении объекта формируется фагосома, которая сливается с лизосомой, содержащей необходимый набор бактерицидных агентов. Здесь происходят гибель объекта фагоцитоза и его расщепление с последующим выбросом продуктов деградации. Основными агентами, вызывающими гибель фагоцитированных клеток, являются продукты кислородного метаболизма (активные формы кислорода, 4 перекиси), производные галогенов, окись азота, некоторые ферменты и другие факторы.
Естественные киллеры лишены антигенраспознающих рецепторов; им свойственно лектиновое распознавание углеводных структур, которые присутствуют на поверхности быстро делящихся (в частности, опухолевых и трансформированных вирусами) клеток и некоторых чужеродных агентов. NK-клетка выделяет в зону контакта перфорин, формирующий в мембране клетки-мишени пору, через которую в нее проникают молекулы, включающие апоптоз клеток-мишеней. Распознавание NK-клетками аутологичных молекул МНС-1 приводит к блокаде цитолиза, что служит механизмом защиты собственных клеток организма.
Цитолиз может быть вызван также гуморальными факторами системы комплемента. Эта система включает несколько компонентов комплемента, активируемых по каскадному принципу. Запуск активационно- го каскада вызывают иммунные комплексы (классическая активация комплемента) или взаимодействие с поверхностью чужеродных клеток (альтернативная активация). Результатом активации системы комплемента является опсонизация клеток его компонентами (в основном СЗЬ) и формирование пор в клеточной мембране. Активация сопровождается освобождением пептидов — продуктов расщепления компонентов комплемента (СЗа, С5а), которые оказывают хемотаксическое и сосудорасширяющее действие.
Клетки, обеспечивающие естественную резистентность, особенно макрофаги, секретируют бактерицидные и другие факторы, в частности провоспалительные цитокины — интерлейкины (ИЛ) 1 и 6, фактор некроза опухоли, хемокины (цитокины с хемотаксической активностью).
Цитокины — белковые продукты клеток, лишенные специфичности в отношении антигенов и определяющие межклеточные взаимодействия при воспалении, иммунном ответе, гемопоэзе, межсистемных коммуникациях.
Связующим звеном между факторами естественной резистентности и антигенспецифической составляющей иммунного ответа служат ан- тигенпрезентирующие клетки. Дендритные клетки кожи (клетки Лангер- ганса) и слизистых оболочек поглощают, обрабатывают антиген, мигрируют в лимфоидные органы и в них «представляют» CD4+ Т-хелпе- рам антигенный пептид в составе молекул МНС-Н. Этот процесс является ключевым событием иммунного ответа. Для активации Т-хелперов, помимо распознавания комплекса антигенный пептид— молекула МНС-И, требуется действие дополнительных сигналов — костимуляция. Она достигается благодаря взаимодействию молекул CD80 и CD86 антигенпред- ставляющей клетки с молекулой CD28 Т-хелпера и молекулы CD40 анти- генпредставляющей клетки с молекулой CD154 на поверхности Т-хелпера.
Активация лимфоцитов является результатом передачи сигнала от антигенспецифического рецептора и дополнительных рецепторов (коре- цепторов) к регуляторным участкам генов, индукция которых необходима для проявления функций лимфоцитов. Сигнал передается путем последовательной активации ферментов (киназ), с участием которых происходит формирование активных форм транскрипционных факторов, способных индуцировать активность соответствующих генов. У лимфоцитов это прежде всего гены ростовых факторов и их рецепторов.
Активация создает предпосылки для пролиферации и дифференцировки лимфоцитов. Важность пролиферации обусловлена исходной малочисленностью клеток в тех клонах,
которые вовлекаются в иммунный ответ.
Для достижения эффективного иммунного ответа численность клеток должна существенно увеличиться. Это достигается путем действия на активированные клетки цитокинов, обладающих активностью факторов роста. Для Т-лимфоцитов это преимущественно ИЛ-2, для В-лимфоцитов — ИЛ-4.
Дифференцировка Т-хелперов происходит при участии контактных стимулов со стороны антигенпрезентирующих клеток, а также действия цитокинов. ИЛ-12 и интерферон у способствуют дифференцировке CD4f- клетоквхелперы первого типа (Th1), ИЛ-4 — вхелперы второго типа (Th2). Th1 - и ТИ2-клетки секретируют различные наборы цитокинов; для Th1 — это интерферон у, ИЛ-2, лимфотоксин и ряд цитокинов, общих с ТН2 (ИЛ-3, ГМ-КСФ и т.д.), для Th2 — ИЛ-4, ИЛ-5, ИЛ-6, ИЛ-9, ИЛ-13 и упомянутые «общие» цитокины. Th1-лимфоциты направляют развитие иммунного ответа в сторону реакций клеточного типа, ТИ2-клетки — в сторону реакций гуморального типа. Между Th1 - и ТИ2-клетками существуют отношения антагонизма.
Основой гуморального иммунного ответа служит реакция В-лимфоцитов на связывание свободного антигена и костимуляцию со стороны ТИ2-хелперов. Взаимодействие В-клеток и Т-хелперов является двусторонним: В-лимфоциты выполняют роль антигенпрезентирующих клеток и в то же время получают от них костимулирующий сигнал, запускаемый через молекулу CD40. Он обеспечивает выживаемость активированных В-лимфоцитов, переключение изотипов иммуноглобулинов в последовательности IgM, IgG, IgE, IgA и обеспечивает непрерывное повышение сродства (аффинитета) рецептора и антител к антигену. Эти процессы завершаются дифференцировкой В-лимфоцитов в плазматические клетки, которые секретируют антитела,
lgM-антитела, образующиеся на начальных этапах иммунного ответа, обладают низким сродством к антигену; lgG-антитела, представляющие собой основной тип антител, обладают более высоким сродством к антигену и разнообразным спектром биологических эффектов. lgA-анти- тела способны проникать сквозь эпителиальные клетки слизистых оболочек и обеспечивать их защиту. lgE-антитела являются ключевыми факторами аллергических реакций I типа, участвуют в защите от паразитов. При связывании антител с растворимым антигеном образуются иммунные комплексы, которые легко фагоцитируются и расщепляются макрофагами. Фиксация антител, особенно изотипа IgG, на клетках-мишенях (опсонизация) делает их более чувствительными к действию факторов комплемента, а также фагоцитов, естественных киллеров, которые имеют Fc-рецепторы, взаимодействующие с «хвостовой» Fc-частью lgG-ан- тител.
Клеточный иммунный ответ обеспечивается Т-клетками. Существует две его разновидности — цитотоксический ответ и гиперчувствительность замедленного типа (ГЗТ). Первый тип ответа осуществляется цитотоксическими (CD8+) Т-лимфоцитами; ГЗТ реализуется в основном С04+-клетками и макрофагами. Цитотоксические Т-лимфоциты, дифференцировавшиеся и размножившиеся под влиянием ИЛ-2, распознают антигенный пептид, связанный с молекулами MHC-I, и лизируют их. Лизис осуществляется путем индукции апоптоза клеток-мишеней в основном по перфориновому механизму (см. п. 5).
ГЗТ развивается в две фазы. Во время индукторной фазы происходят сенсибилизации CD4+ Т-клеток в ответ на действие антигена в составе молекулы MHC-II и их дифференцировка в хелперы Thl-типа, При повторном поступлении антигена развивается эффекторная фаза ГЗТ: Th1-клетки мигрируют в очаг поражения, выделяют интерферон <gamma>, активирующий макрофаги, которые проявляют фагоцитарную, секреторную и киллерную активность. Цитокины, выделяемые Т-клетками при всех вариантах иммунного ответа, активируют естественные киллеры, значительно повышая цитолитическую эффективность.
Параллельно формированию эффекторных механизмов в процессе иммунного ответа образуются клетки памяти. Это — долгоживущие лимфоциты Т- и В-классов, которые непосредственно не участвуют в первичном иммунном ответе, но обеспечивают ускоренное развитие более интенсивной и генерализованной реакции на повторное поступление того же антигена — вторичного иммунного ответа.Контроль и регуляция иммунного ответа
Способность к иммунному ответу контролируется генетически. Уровень ответа на конкретные антигены определяется генами, локализованными в главном комплексе гистосовместимости (МНС), эффект которых реализуется на уровне обработки и «представления» антигенных пептидов антигенпрезентирующими клетками Т-хелперам.
Регуляция уровня иммунного ответа осуществляется гормонами и медиаторами вегетативной нервной системы, а также регуляторными пептидами. Характер действия гормонов определяется их концентрацией в крови. При физиологической концентрации они, как правило, способствуют развитию иммунного ответа. При высокой (фармакологической) концентрации некоторые из них по преимуществу ингибируют иммунные процессы (это относится к гормонам коры надпочечников, являющимся классическими индукторами апоптоза лимфоцитов, АКТГ, адреналину, половым гормонам, (3-эндорфинам, адренергическим медиаторам), другие (гормон роста, пролактин, гормоны щитовидной железы, инсулин, а-эн- дорфины, холинергические медиаторы), как правило, стимулируют иммунный ответ. В то же время ряд цитокинов (особенно ИЛ-1) оказывает влияние на активность нервной и эндокринной систем. Пептидные гормоны, выделяемые эпителиальными клетками вилочковой железы — ти- мулин, тимозины, тимопоэтины — могут быть отнесены одновременно к продуктам иммунной и эндокринной систем.
Иммунный ответ подвержен ауторегуляции. Накопление lgG-анти- тел в свободном и особенно связанном с антигенами виде подавляет дальнейшее развитие гуморального ответа, действуя через Fc-рецепторы ре- гуляторных и эффекторных клеток. Ингибирование достигается также по механизму идиотипического контроля — через образование антител к эпитопу антигенраспознающей части молекулы антител (их идиотопу).
Основной клеточный механизм иммунорегуляции реализуется с участием супрессорных Т-клеток. Супрессорную функцию выполняют в основном CD8+- клетки. Главным объектом ихантигенспецифическогоин- гибирующего влияния являются Т-хелперы. CD8+-T-cynpeccopbi оказывают свое влияние как путем контактных взаимодействий, так и через выделение гуморальных факторов. С04+-клетки также могут выступать в роли супрессоров, что проявляется во взаимном подавлении активности Th1- и Т112-хелперов. Супрессорной активностью обладают также активированные макрофаги и NK-клетки.