

9654
.pdf31
Таким образом, зеленый цвет листа обеспечивает наиболее эффективное
«энергоснабжение» фотосинтеза.
Рис. 6. Схема поступления прямой (S) и рассеянной (D) солнечной радиации в растении с горизонтальными (А), вертикальными (Б) и различно ориентированными (В) листьями (по Шульгину И.А., 1967):
1, 2 ― листья с разными углами наклона, S1, S2 ― поступление к ним прямой радиации, Sобщ ― ее суммарное поступление к растению
Лист как оптическая система отличается чрезвычайно сложной и неоднородной внутренней структурой: и на уровне тканей (различные слои клеток, межклетники, проводящая система), и на уровне клетки
(хлоропласты, способные к перемещениям и поворотам), и на уровне хлоропласта (система ламелл и гран, распределение молекул пигмента в гранах). Разнообразие структурных элементов и обилие межфазных границ обусловливают большое внутреннее рассеяние и отражение света. Поэтому даже тонкий лист поглощает света значительно больше, чем слой раствора пигментов соответствующей концентрации. Вместе с тем сложность внутренней структуры листа предоставляет большие возможности ее
32
перестройки на разных уровнях (тканевом, клеточном, внутриклеточном) в
зависимости от условий освещенности (см. рис. 8—11).
Кроме собственно оптических свойств листа на количество по-
лучаемой им радиации большое влияние оказывает положение листа как приемника солнечной радиации в пространстве. Большое значение имеет угол наклона листовой пластинки, от которого зависит приход солнечной энергии к листу. На рис. 6 показано, как различаются по получаемой радиации растения с различными типами расположения листьев. Следует отметить, что в связи с неодинаковой высотой Солнца на разной географической широте одни и те же углы наклона листьев означают для растений различный радиационный режим.
Поскольку растения получают прямую солнечную радиацию от направленного (хотя и перемещающегося) источника, большую значимость имеет азимутальная ориентация листовой пластинки (если она наклонена). Иными словами, поступление радиации к листу будет весьма различаться в зависимости от обращения листовой пластинки к югу или к северу (рис. 7).
Идеальным решением вопроса был бы «следящий» лист,
поворачивающийся вслед за солнцем, однако такие случаи в растительном мире очень редки (известен пример подсолнечника, поворачивающего листья обвертки вместе с соцветием). Обычно ориентация листьев на растении более или менее диффузна, т. е. листья «смотрят во все стороны»,
что дает возможность в разные часы дня получать наибольшее количество радиации то одним, то другим частям растения. По выражению И. А.
Шульгина (1973), растение можно назвать своего рода неподвижной локаторной системой, в которой часть приемников лучистой энергии всегда ориентирована к оптимальному потоку радиации в течение большей части дня.

33
Рис. 7. Приход ФАР на площадки |
Рис. 8. Разные величины хлоропластов |
различной азимутальной ориентации |
у теневыносливых (А) и светолюбивых |
(для 55о с. ш. по Шульгину) |
(Б) растений |
Рис. 9. Изменение положения хлоро- |
Рис.10. Перемещение хлоропластов |
|||
пластов в клетках листа ряски при |
в клетках ряски из диффузного |
|||
различной освещенности. Вверху – |
положения |
в |
постенное |
при |
древесные породы (по Любименко В. |
действии |
сильного |
света |
|
Н., 1924): |
(киносъемка) (по Zurzycki J., 1953): |
|||
1 ― тис, 2 ― лиственница; внизу – |
Заштрихованные |
кружки |
– |
|
травянистые растения дубового леса |
положение |
хлоропластов |
до |
|
(по Горышиной Т. К. и Пружиной Е. |
освещения, черные кружки – ко- |
|||
Г., 1978): 3 ― копытень, 4 ― чистяк |
нечное положение, извилистые |
|||
весенний |
линии – пути перемещения |
|
||

34
Однако в определенных случаях наблюдается строго направленное расположение листовых пластинок. Это бывает или при одностороннем освещении при общем недостатке света (например, листья лесных трав близ опушки или близ «окна» в пологе древостоя все повернуты в сторону света), или при избыточном освещении.
Одна из защитных реакций хлоропластов на избыточный свет – их способность к изменению ориентировки и к перемещениям в клетке,
особенно выраженная у световых растений; в листьях с многослойным мезофиллом эта способность проявляется в наиболее сильно освещаемой палисадной ткани. На сильном свету хлоропласты занимают в клетке постенное положение и становятся «ребром» к направлению лучей. При слабом освещении они распределяются в клетке диффузно или скапливаются в ее нижней части (рис. 9). Такие движения хлоропластов наблюдаются как при естественной суточной смене освещения, так и в экспериментах. С помощью микрокиносъемки удается проследить перемещение отдельных хлоропластов, которое происходит в течение 15—
20 мин после изменения освещенности (рис. 10). Изменение ориентировки в зависимости от силы и направления света отмечены и у хроматофоров водорослей (рис. 11).
Рис. 11. Схема изменения положения хлоропластов и хроматофоров в клетках растений под влиянием света (по данным разных авторов):
А — Letnna trisulca, Б — Моиgeotia sp., В — Selaginella martensii; 1 — слабый свет, 2—
сильный свет, 3 — темнота. Стрелки показывают направление освещения
35
2.2. Физиологические адаптации
Физиологические адаптации растений к световым условиям ме-
стообитания охватывают различные жизненные функции. Известно,
например, что у светолюбивых растений ростовые процессы более чутко реагируют на недостаток света, чем у теневых. В результате наблюдается усиленное вытягивание стеблей, помогающее растениям «пробиться» к
свету, в верхние ярусы растительных сообществ. Особенно большое значение это свойство имеет для светолюбивых лиан.
Основные физиологические адаптации к свету лежат в сфере фотосинтеза и фотосинтетического аппарата. Остановимся вначале на особенностях пигментной системы световых и теневых растений. Еще в начале века один из первых исследователей эколого-физиологических адаптаций растений к условиям освещенности акад. В. Н. Любименко на основании исследования более чем 600 видов растений пришел к выводу о том, что листья теневых растений по сравнению со световыми содержат повышенное количество хлорофилла. Это хорошо заметно уже по внешнему виду листьев, обычно в затенении имеющих более густую темно-зеленую окраску. Правда, это различие не всегда улавливается при расчете количества хлорофилла на единицу площади листа, поскольку листья световых растений толще, но оно хорошо заметно при сравнении содержания хлорофилла в единице массы листа. Так, у растений хорошо освещенных местообитаний листья содержат 1,5—3 мг хлорофилла в 1 г
листа, а у видов, растущих в тени, – 4–6 мг (в отдельных случаях до
7–8 мг). В последние годы обнаружено, что у теневых растений концентрация хлорофилла в одном хлоропласте значительно больше, а
поскольку сами хлоропласты крупнее, то общее количество пигментов в одном хлоропласте может быть в 5–10 раз больше, чем у световых растений.
36
Таблица 1
Средние величины отношений хлорофилл «а»/хлорофилл «b» (I) и хлорофиллы/каратиноиды (II) у растений из мест обитаний с различной освещенностью (по Egle K., 1960)
Группа растений |
|
I |
|
II |
|
цветковые |
папоротники |
цветковые |
папоротники |
||
|
|||||
|
|
|
|
|
|
Альпийские растения |
5,6 |
- |
4,6 |
- |
|
Водные растения |
4,4 |
- |
4,0 |
- |
|
(плавающие) |
|||||
|
|
|
|
||
Растения открытых, |
|
|
|
|
|
хорошо освещенных |
|
|
|
|
|
местообитаний |
4,4 |
4,0 |
3,6 |
4,7 |
|
Растения «синей тени» |
|
|
|
|
|
(использующие |
|
|
|
|
|
рассеянный свет неба) |
3,0 |
2,9 |
5,5 |
5,8 |
|
Растения «зелѐной тени» |
|
|
|
|
|
(в нижних ярусах |
|
|
|
|
|
растительных сообществ) |
2,6 |
2,4 |
5,3 |
5,5 |
|
Водные растения |
2,3 |
- |
5,7 |
|
|
(погруженные) |
|
||||
|
|
|
|
От условий освещенности зависит не только количество, но и состав пигментов листа. Известно, что хлорофилл в листе представлен целым рядом форм с несколько различным строением молекулы и разными спектрами поглощения света. У растений открытых и хорошо освещенных местообитаний форма «а» значительно преобладает над формой «b», а в затенении доля хлорофилла «а» падает и отношение а : b гораздо меньше
(табл. 1).
2.3. Сезонные адаптации растений к световому режиму
В тех местообитаниях, где затенение действует на растения не все время, а в течение определенного отрезка вегетационного сезона,
существует еще одна возможность адаптации растений к световому режиму – такая организация жизненных процессов во времени, при которой основной период ассимиляции не совпадает с периодом сильного затенения. Яркий пример такой сезонной адаптации представляют травянистые растения листопадных лесов. Как уже было показано, в
37
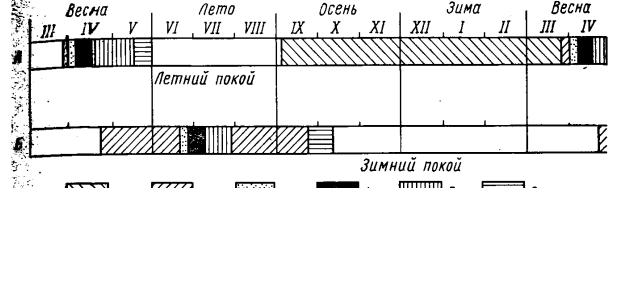
течение вегетационного сезона в их травяном покрове освещенность неодинакова: непродолжительный светлый весенний период до распускания почек на деревьях сменяется глубоким и длительным летним затенением, а осенью после листопада наступает осветление. Весенняя экологическая ниша используется светолюбивыми эфемероидами – многолетними травянистыми растениями с краткой (всего несколько недель) вегетацией и длительным покоем. В наших дубовых лесах это пролеска Scilla sibirica, хохлатка Corydalis halleri, гусиные луки Gagea lutea, G. minima, ветреница лютиковая Anemone ranunculoides и др. Они прорастают и начинают цвести сразу же после таяния снега в лесу, а ко времени полного развития листвы на деревьях и установления тени уже заканчивают вегетацию и теряют надземные части. Период глубокого затенения эфемероиды переживают в состоянии летнего покоя, в виде подземных органов – луковиц, клубней, корневищ.
Рис. 12. Схема годичных фенологических циклов у основных видов двух сезонных синузий в травяном покрове дубравы:
А – ранневесенний эфемероид Scilla sibirica; Б – летний доминант Aegopodium podagraria;
фенофазы: 1 – подземный и подснежный рост, 2 – надземная вегетация, 3 – бутонизация, 4 – цветение, 5 – плодоношение, 6 – отмирание
В западноевропейских листопадных лесах (дубовых, буковых,
грабовых) группа ранневесенних эфемероидов представлена такими видами, как Scilla bifolia, Galanthus nivalis, Endimion non-scriptus, Eranthis
38
hiemalis и др.; в североамериканских дубовых и кленовых лесах —
Erythronium americanum, E. canadense, Allium tricoccum и др. Есть своя группа эфемероидов и в дальневосточных широколиственных лесах.
Таким образом, это довольно распространенный способ адаптации к временному затенению, который состоит в перемещении всей вегетации на светлый отрезок благоприятного сезона. Это перемещение достигается путем сдвига всего годичного цикла, так что он не совпадает с годичной климатической ритмикой умеренных широт и обычной сезонной ритмикой летневегетирующих растений (рис. 12). Ранневесенняя вегетация дает возможность эфемероидам избежать затенения под пологом леса, но зато требует повышенной холодостойкости, способности к быстрому росту и развитию при низких температурах, заблаговременной подготовки к цветению и т. д.
2.4. Фотопериодизм
Важнейшая характеристика светового режима – фотопериод, т. е.
продолжительность дня (или, точнее, соотношение длины светлой и темной частей суток), которая неодинакова в течение года.
Закономерность изменения длины дня на разных широтах представлена на рис. 13. Длина дня не безразлична для растений. Очень многие виды переходят от вегетативного развития к генеративному (цветение и плодоношение) только в том случае, если они развиваются при фотопериоде, не превышающем (или в других случаях – не ниже)
определенной критической величины. Способность растений реагировать на длину дня получила название фотопериодической реакции (ФПР), а
круг явлений, регулируемых длиной дня, именуется фотопериодизмом.
Фотопериодизм был открыт в 1920 г. американскими учеными У.
Гарнером и Г. Аллардом. Проводя селекционные работы с табаком, они обнаружили, что один из сортов, который цвел весной и осенью в теплице,
не зацветает летом в открытом грунте. Поскольку летние условия почти не
39
отличались от тепличных, они предположили, что цветению препятствует длинный летний день. Это предположение подтвердилось, когда удалось получить цветение табака летом, но при искусственно укороченном дне.
С тех пор в исследованиях по фотопериодизму накоплен огромный экспериментальный материал. Показано, что фотопериодическая реакция свойственна растениям самых разных таксономических групп и жизненных форм, причем не только цветковым, но мохообразным и водорослям; пока мало сведений о ФПР у плаунообразных и папоротникообразных. Способность воспринимать длину дня и реагировать на нее широко распространена и в животном мире.
По типу ФПР различают следующие основные группы растений:
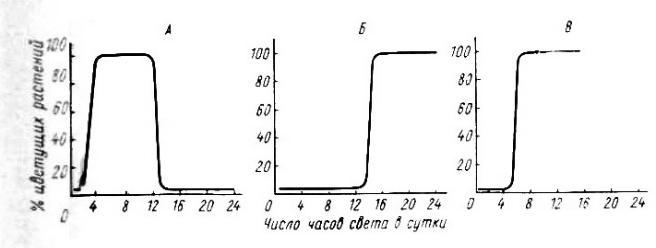
1. Растения с короткодневной ФПР (или растения короткого дня),
которым для перехода к цветению требуется 12 ч. и менее света в сутки
(например, конопля, табак, перилла).
Рис. 13. Типы фотопериодической реакции у растений (по Мошкову Б. С, 1961):
А — короткодневный (перилла); Б — длиннодневный (шпинат); В — нейтральный (горчица)
2. Растения с длиннодневной ФПР (растения длинного дня); для цветения им нужна продолжительность дня 12 и более часов (таковы картофель, пшеница, шпинат).
40
3.Некоторые виды (например, гваюла) имеют ФПР промежуточного типа, т. е. цветение у них наступает при определенном, сравнительно узком диапазоне фотопериода – не длиннее, но и не короче юитических величин.
4.Наконец, есть растения, фотопериодически нейтральные – для них длина фотопериода безразлична, и цветение наступает при любой длине дня (кроме очень короткой, означающей для растений световое голодание).
Таковы томат, одуванчик и др.