

3 курс / Патологическая физиология / Основы общей патологии
.pdf! |
101 |
Рис. 6Связь. междунаследственност |
ью,влиянвнешнейсредыиреактивностьюем |
разаболеваниявитиипо(В.И.Арнольду). |
|
В — врачебноевоздействие; |
|
С — влияниевнешнейсреды; |
|
Р — реактивность; |
|
1, 2 — точкисостоянийсистемы: |
|
3,4 — траектоизмененияпарамеуправленияотк12ров. |
|
Этапроеимеетскладкициясборку.Совокупностьскладок |
— |
||
криваякатастроф.При |
[57] переходеэтойкривойчислопрообразов |
||
проекцийскачкообразноменяется.Нобудетли,зависитотк |
|
|
|
траекторииизменений, толькоотначальногоконечн го |
|
— определяетсянетолько |
|
пунктов. |
Тоесть,будетлиболезнь |
|
|
начальконечными з ачениемпарамет,которыеизменилисьов подвнешнвоздействием,нотемспособом,кот стигнуторым этоизменение.
! |
102 |
|
При изменении параметров управления (рис. 6) от точки 1 к |
точке 2 по траектории 3 произойдет пересечение кривой катастроф, и проекция точки окажется на качественно новом уровне.
Это можно уподобить такой комбинации С, В и Р в предыдущие моменты, когда наступила болезнь (здоровый организм с нормальной индивидуальной реактивностью превратился в больной, с качественно особой жизнедеятельностью).
Тот же путь от точки 1 к точке 2 мог быть проделан по другой траектории 4, при этом линия катастроф не пересекается и все точки сохраняют то же число прообразов, то есть переход идет без скачка и болезнь не наступает, хотя значения С и В пришли к тому же уровню, что в первом варианте.
Образно говоря, в зависимости от реактивности, индивид может выбрать различные пути, ведущие с шестнадцатого этажа на первый: через лифт, по черной лестнице и … через окно. Очевидно, что достигнув одних и тех же конечных координат, он окажется, в зависимости от выбранного пути и стоимости адаптации, в весьма разном, не обязательно — здоровом, состоянии.
В свете теории катастроф реактивность выглядит как индивидуальный внутренний механизм выбора системой того или иного пути в пространстве адаптивных состояний.
СУБСТРАТ РЕАКТИВНОСТИ И ЭВОЛЮЦИЯ ЕЕ ИНТЕГРАТИВНЫХ МЕХАНИЗМОВ
Хотя понятие «реактивность» принадлежит организму как целому, конкретные механизмы реактивности могут реализоваться, преимущественно, на каком-либо из структурно-функциональных подуровней. Поэтому субстрат реактивности может быть условно отнесен к какому-то уровню организации живой системы. Говорят о реактивности на молекулярном, субклеточном, клеточном, тканевом и органном уровне, выделяют организменные и даже популяционные формы реактивности.
На молекулярном уровне реактивности ключевое значение
Рекомендовано к покупке и изучению сайтом МедУнивер - https://meduniver.com/
! |
103 |
имеюткомплементарныевзаимодействия,ос |
нованные |
однозначномструктурномсоответствиираспознающихмолекул. |
|
Так,рецепторсубъединиаденилатциклазыаяспецифически узнаетпептидныйбиорегуляторвзаимодействуетнимпо принципуключ« -замок»,чтоведетак каталитическойивц субъединицыэтогофермента.Комплементарностьпроявляетсяво взаимодействферментовихсубстратов,антигеновантителях,
цис-регуэляторныхементовхроматиналигандов,изменяющих экспрессиюгенов.
[58] ВпослврработамиеднеемяДж. |
Блэлокаисоавторов |
|
||
(1986-1990)пока,чтовзакоиномноплементарныенитиДНК |
|
|||
кодируютбелковыемолекулы,котор,своюоче,преоявляютедь |
|
|
|
|
междусобойструктурноеоднозначноесоответствие.Иными |
|
|
|
|
слова,гориихмрецепторыионынапример( ,рецепторАКТГ |
уются,соответственно,смысловойи |
|
||
адренокортикотропин)кодир |
|
|||
антисмысловойРНК,которыесписанысимметричных |
|
|
|
|
комплементарныхкодирующучастковДНК.Еслэтотпринцип |
|
|
|
|
окажеуниве,тпсяорсальпризиде,чтраспознающиесяымать |
|
|||
молезакодирвулыгеномепоп«симметричварноны |
|
|
но»и |
|
каждбелкусоответствуетмукодегоантибелка |
-визавиилисвоего |
|
||
родааутоантитела« ». |
|
|
|
|
Вэтомслучаедревнепик,итайскуюостоящграммуиз ю |
но |
|||
взаикомноплементарныхфигурИнь«»Ян»« ( |
рис. 7)мож |
|||
будетсчитатьпророческимизображениемсмысловойструктуры |
|
|
|
|
геномаилипринципакомплементреактивности.Пок,чторнойзано |
возникактахтр ютнскрипцииЖ.( |
- |
||
антисмысловыеРНК |
||||
Ж. Тульме, и1986)служатрегулят,таккакспокрамисобны |
|
|
|
|
комплементарному блокированиютрансляциисмысловыхрепл к
{6}.

! |
104 |
Рис. 7. Древнекитайский символ "Инь" и "Ян" — символическое изображение комплементарности
Принцип комплементарности, с которым мы столкнулись уже при рассмотрении элементарного молекулярного уровня субстрата реактивности, носит сквозной характер и присутствует на всех уровнях организации живого.
Механизмы реактивности, свойственные ее субклеточному и клеточному уровням, подробно охарактеризованы в последующих разделах. (Патология сигнализации, Патология рецепции и т.д.). В данном общем разделе хотелось бы только подчеркнуть, что молекулярные, субклеточные и клеточные механизмы реактивности несут на себе печать индивидуальности в той же мере, в какой и ее высшие интегральные проявления. Реактивность различна для тех или иных молекул, органоидов и клеток, взятых от различных индивидов и из различных тканей. Фетальный гемоглобин и гемоглобин А по-разному связывают кислород. Митохондрии скелетных мышц предпочитают в качестве энергетических субстратов активные одноуглеродные фрагменты, полученные из глюкозы, а митохондрии кардиомиоцитов — фрагменты, полученные из жирных кислот. Тканевой и кровяной тромбопластин различаются по составу и механизмам образования. Макрофаги разных тканей, например, печеночные клетки Купфера и остеокласты, несмотря на общность происхождения, выглядят по- разному и т.п.
Следующими иерархическими уровнями субстрата реактивности
Рекомендовано к покупке и изучению сайтом МедУнивер - https://meduniver.com/
! |
105 |
являются тканевой и органный. |
|
Помередифференцировки,клеткиостав |
ляютвак ивно |
используепрограаппаратетолькомчастьмномгенетически унаследовпр, грстархивируютсяммльныенных.Поэтомуони отвечаютнараздрре ,свойсжениякциямиданнойтвека. ными Примеромпроявлтканевыхханизмовнияреактивностиможет служитьмесоваскуляризованныхветныйтканейнаповреждение
— воспаление.
Сразвитиеморганогенезаможносвязатьначалоформирования системныхответов,таккакаждыйоргансостоитизразличных тканей.
Важнойсостканевогоавляющейиорганногосубстрата реактивноявляетсяти структурно-функциональныйэлемент органа (ткани).
Несмотрянаразличияназванийнефрон(почек,печеночная |
— вмышцеит.д.) |
|
долька — впечени,двигательнаяединица |
||
структурно-функцедиоргановонитканцыимльныеютй |
М. Чернуху,микроциркуляторные |
|
общиечерты |
строения.ПоА. |
|
сосудистыеединицытипичногостроенияслужат |
[59] структурной |
|
осью,вокругкоторойгруппируютсясоединительно |
-тканные |
|
элементыстроморгана,вып полняющие,трофическуюную |
рганнойпаренхимы. |
|
защитнуюфункциюдляэлементов |
||
Структурно-функциэлементсоединительнойнальтканиый |
||
Чернухназвал« |
гистионом»При.любомповрежденииоргана |
|
специализировзащитфункциивовачаыеленныекаются |
|
|
элементыстроми,вполневозм,чтопринебольшихжно |
|
|
масштабахпроц |
ессэтимогра,невызываяичивается |
|
существенногорасстройствафункцийоргп ннойренхимы.Именно |
М. Чернуха,мобъяснитьжно |
|
этим,сточзренияконцепцииА. |
||
наличиебезжг патлтушных,клкоторыхиникатовпредставлена, |
ойзащитреакции.Знойачение |
|
основном,симптостромальнами |
||
этогооригинальногоподходамывидимто,чтогистионы |
- |
|
дейслужаттвительпервичаренойзащитной |
||
приспособреакц,такиепатойпроцессыьныхлогические,как |
— развертываются |
|
воспаление,гипер,иш, емия,тазромбоз |
||
! |
106 |
именно в гистионах.
Дублирование деятельности множества структурно- функциональных единиц, составляющих орган, определяет надежность системы, даже если при гибели элементы и не могут регенерировать.
Вздоровом организме, как указывают Адо и Новицкий, используется 20-25% нефронов, 12-15% паренхиматозных элементов печени и т.д.
Всвязи с этим, функциональные пробы, оценивающие парциальные функции почек, порой не могут выявить нарушений у больных, значительный процент почечной паренхимы которых уже поражен нефросклерозом.
На уровне систем органов создается качественно иная интеграция механизмов реактивности — вокруг технологической задачи той или иной системы, при участии взаимодействий, описанных П. К. Анохиным (см. предыдущий раздел).
В основе деятельности каждой из систем, интегрирующих механизмы реактивности, будь то нервная, эндокринная или иммунная, лежит, опять-таки, комплементарное взаимодействие регулятора с рецепторно-дискриминаторной системой. Его сквозной характер не случаен: однозначное соответствие одной молекулы другой служит формой структурного отражения. А отражение — добиологическая основа реактивности. Комплементарные взаимодействия, вероятно, возникли раньше чем клетки. По крайней мере, известные нам доклеточные формы жизни — вирусы и прионы — способны к комплементарному взаимодействию с клетками. Прионы ведут себя, как некие комплементарные белковые сигналы, способные при попадании в клетку активировать древнейшие убиквитарные генетические программы, обеспечивающие воспроизводство этих агентов.
Если жизнь началась с прионов, что вполне вероятно, это означало бы, что комплементарное взаимодействие было решающим шагом в ее появлении.
Рекомендовано к покупке и изучению сайтом МедУнивер - https://meduniver.com/
! |
107 |
Понекоторымпредставозниклиниям,первымирецепторные белки,скореевсего,выполнявшиепоначалурольрепресс орных регуклеяпроцто.Необходимостьчныхровссовагента, снимающегоограничение,вызваладавлениеотборазакрепление мутаций,приводящихкобесродспечениюпротогормоновва рецепторов.Появилисьбелкби выеразнообразие, егуляторы которыхвдальн ейшемнарасталоприотносительномконсерватизме рецепторов.Возм,наобжноордестлястоероидныхло гормонов:тоестьразнообразрецепторовнарасталопри эволюционнойстабильностирегуляторов.
Иммуноглобулины,возникшиенаосновегомологичных |
|
|
распознающихбелков,представленныхуорганизмов,не |
|
|
обладающихиммуннойсистем,оказалисьвсостоянииобеспечитьй |
|
|
уникальноеразнообразиерегуляторныхсигн.Какужелов |
|
|
отмечалосьвыше,реактивностьможетрассматриватьсякак |
). |
|
дискнаборретныйактонов( |
функциональныхфенов |
|
Функционкаждтакедобеспечиваетийрованницыинд виду |
|
|
преивсмывоущеспроизводствале, жевремяимеет |
|
|
определецену.Прималыхиннуютенсивностяхиспользованиятого |
го,нопри |
|
илииногореактонавыигрышрастетбысплзнеарееты |
|
|
высоких — ростплатыопережаетприросвы,этогрышаделает |
|
|
присспомданнсореактоблениещьюпасгов( нпоаымним |
|
|
определеГеккелюболезпо !)нипатогеннымподробнее( см. |
|
|
А. Н. Горбань,Р. |
Г. Хлебопрос, 1988). |
|
Нореактивностьвключае тнетолькореаи, ктоны функциусткомбийчивыенальсоздаваемыереактонов, ации интеграаппаратомпо( ерминологииивнымК. Люкаса(1909) — включаетспособифункциональноеповедениеости,использующее ихпутемкомбинации).
Новыереактоныэволюции возникаютнамногореже,чемновые функциональныеомбинацстарыхмозаблоковчных (А. М. Уголев, Р.1985)Гийменназвалэтооппортунизмом.
эволюционпроцессаилиприиспн«ципомогот,чтользуйд рукой» (1984).
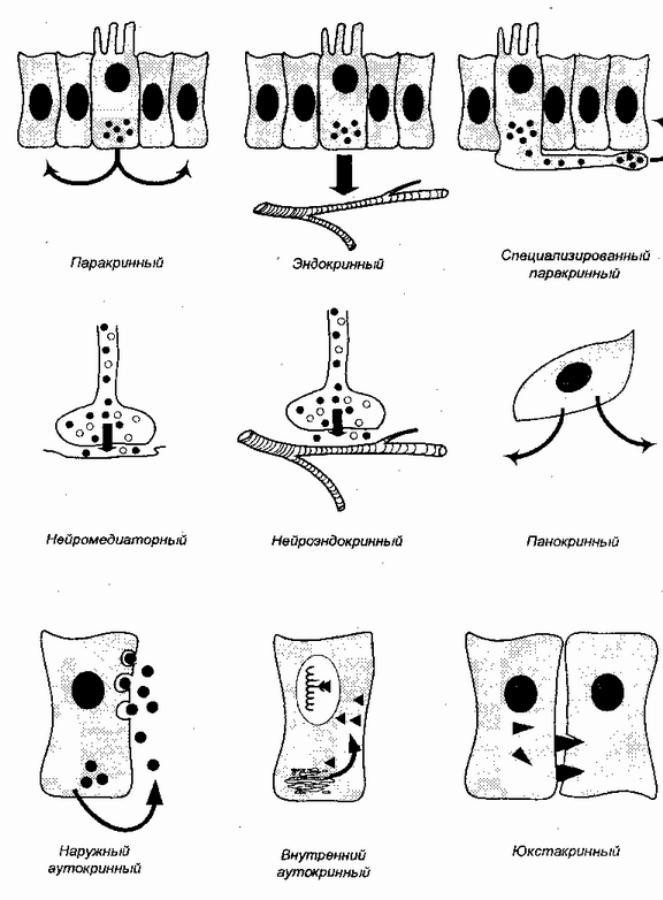
! |
108 |
Рис. 8. Различные типы химических регуляторных взаимодействий между клетками (по Г. Дж. Докрею и К. Р. Хопкинсу, 1982)
Рекомендовано к покупке и изучению сайтом МедУнивер - https://meduniver.com/
! |
|
|
|
|
109 |
|
|
|
Тиротропгормонобнаруженбактерий,ыйинсулинего |
|
— упростейших,а |
||||||
рецептор — удрозофил,тропныегормоны |
|
|||||||
нейропептиды — угубок,неимеющихЦНС.Морфинизмакаимее |
|
|
т |
|||||
кодируемыесходгеаналогиымиамивдиффузнойэндокринной |
|
|
|
|||||
системеживотных |
|
— эндорфэнкефалины, орфины |
Трабукки, 1978)Это . |
|
||||
(Дж. Хьюз, X. |
Костерлиц, 1975,Э. |
Коста,М. |
Снайдер, |
|||||
жеотникопсирецепторамтсяатнымЛ.( |
|
Терениус,С. |
||||||
Э. Саймон, 197 |
4). |
|
|
|
||||
Вазопрессинумлекопидейснаобитающихвуетр,ательныеубки |
|
Ю. Йейтс, 1982). |
||||||
ауамфибий |
|
|
|
— наклеткикожФ. ( |
|
|||
Биохимическиеэлементырегулясисэвтемлюционируютрных |
|
|
|
|||||
какцелоеВ(. |
|
|
Ле Руа, 1987). |
|
|
ьнового |
||
Следовательно,эволюцияреактивностииспользуетпут |
|
|
||||||
поведенческогоиинтеграткомбстарыхивногонирования |
эволюционныеизобретения |
|
||||||
эволюционныхнаходок,реактоныка |
|
|
||||||
непропадают,хотяивостребуютсяпоройдляиныхфункций. |
|
|
|
|||||
Эволюциявыступаеткакмножественныевариациинаогранич нное |
|
|
|
|||||
числотем |
|
-архетипов. |
|
|
|
|||
Ф. Ницшеоснэтомовалпринципесвойизвесафоризм,такный |
|
|
|
|||||
раздражающийнекоторыхистоп ическихогрессистов |
|
|
|
|||||
оптимиЛюди(«,проделавстпучервяйткьчел, овеку |
|
|
|
|||||
многомосталисьчервями»). |
|
|
|
|
регуляции |
|||
Филогдревнейшийнетспособхимическойчески |
|
|
||||||
клеточнойреактивности |
— аутокринный.Онможетбыть |
|
||||||
внутре,когданнимоклеточныйорганизмвырабатывает |
|
|
|
|||||
химическиерегуляторы,действующие,нвыходяизклетки.Есл |
|
|
|
|||||
биорегувыдедейлянасторетсяамтввырабатывающуюетего |
|
|
|
|||||
клетку,говорят |
|
|
|
онаружнойтокриннойрегуляции.Аутокринные |
|
|
||
воздействияинтерлейкина |
-2 — важныйэлементкооперацииклеток |
|
||||||
виммунномответе.Когдабиорсегуляторыкретируютсявовне |
|
|
|
|||||
действуютнадругиеклетки,проявляетсямеханокринныйизм |
Р. |
Хопкинсу, При1982). |
|
|||||
(см. |
рис. |
8,поГ. |
Дж. ДокреюиК. |
|
||||
|
|
|
|
|
|
|
|
|
панокриннойрегуляцклеткавыделяб иорегулятор окружающеепространство,жидкуюсредуоб литания. чву Такспсуществуетойсободноклеточныхипростых многоклеточных,напримерлесневыегрибыводоросливыделяют антиби,подавляярбактерийсттики.Ж антибиотиквотный лизоцим,присутствующийпрактическивосехо етахганизма,
! |
110 |
может расцениваться, как наследие панокринной регуляции в реактивности организма. Юкстакринное воздействие предполагает, что липофильный биорегулятор остается в мембране клетки- источника и активируется контактом с клеткой-мишенью. (М. Спорн и соавт. 1992).
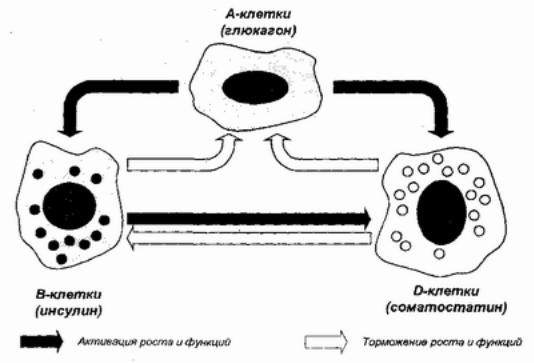
Следующим эволюционным шагом могло быть возникновение паракринной (термин Ф. Фейртера, 1938) регуляции у многоклеточных. При паракринном воздействии биорегулятор выделяется в области тесных клеточных контактов и влияет лишь на реакции ближайших соседей клетки, связанных с нею этими цитологическими структурами. В организме человека древняя паракринная регуляция сохраняется. Примером могут служить взаимоотношения основных клеточных элементов островка Лангерганса в поджелудочной железе. Три основных типа клеток — α, β, и δ образуют гетероклеточные зоны и, помимо выделения своих биорегуляторов — глюкагона, инсулина и соматостатина — в системный кровоток, в качестве гормонов, используют их для локального воздействия друг на друга через тесные клеточные контакты как паракринные сигналы. [62] Как паракринные клеточные сигналы, инсулин и соматостатин
ингибируют секрецию и пролиферацию α-клеток (рис. 9).
Рис. 9. Паракринное взаимодействие клеток островка Лангерганса (по Унгару-Орчи)
Рекомендовано к покупке и изучению сайтом МедУнивер - https://meduniver.com/