

Расогене́з — процесс возникновения и становления человеческих рас[1].
Находки черепов позднепалеолитических людей говорят о том, что основные особенности главных расовых делений человечества, существующих в настоящее время, уже были выражены в эпоху позднего палеолита достаточно отчётливо, хотя, по-видимому, всё же меньше, чем в настоящее время. Они более или менее точно совпадали с границами материков. Европеоидная раса сформировалась преимущественно в Европе, монголоидная — в Азии, представители негроидной расынаселяли Африку, австралоиды Австралию. Исключение составляли пограничные зоны — Средиземноморье, где на европейском побережье встречались представители негроидной расы, а на африканском — европеоидные группы; Кавказ и Средняя Азия, заселённые преимущественно представителями европеоидной расы; Южная и Юго-Восточная Азия, где австралоиды смешивались с монголоидами и европеоидами. Таким образом, в образовании морфологических различий между тремя большими расами человечества (расовых различий первого порядка) основная роль принадлежала, вероятно, двум факторам: приспособлению к среде, различавшейся на разных материках, и изоляции в результате естественных границ между материками.
Среда обитания негроидов характеризуется высокой температурой и большой влажностью, что могло привести к появлению тёмной кожи, курчавых волос, широкого носа, толстых губ. Так, тёмный цвет кожи объясняется наличием в покровных слоях кожи большего, чем у других рас, количества меланина — пигмента, предохраняющего кожу от ожогов. Курчавые волосы создают вокруг головы воздухоносную прослойку, которая предохраняет от перегрева. Широкий нос с крупными ноздрями и толстые губы с обширной поверхностью слизистой оболочки усиливают теплоотдачу, как и большее количество потовых желёз на единицу поверхности тела. Большая высота и удлинённая форма черепной коробки также способствует защите от перегрева.
Монголоидная раса сложилась в областях с жарким, но сухим континентальным климатом в условиях полупустынного и степного ландшафта, где ветер гонит тучи песка. В результате лицо представителей монголоидной расы покрыто слоем жира, значительно превосходящим слой жира на лице представителей других рас. Глаза характеризуются узким разрезом и наличием особой складки во внутреннем углу глаза — эпикантуса. Аналогичные особенности сформировались у бушменов иготтентотов Южной Африки, также живущих в условиях полупустынного и степного ландшафта.
Наиболее характерная особенность европеоидной расы — выступающий нос — может быть объяснена сравнительно суровым климатом Европы в конце четвертичного периода, когда необходимо было предохранить организм человека от переохлаждения. Сильное выступание носовой полости удлиняло путь воздуха до дыхательных путей и способствовало его согреванию.
Кроме того, все расы характеризуются комплексом нейтральных признаков. Формирование многих деталей строения тела — результат случайной концентрации этих признаков в замкнутых ареалах первоначального распространения основных расовых делений человечества, а также корреляционной изменчивости, то есть изменения одного из двух признаков в том случае, если изменился другой. Например, при изменении ширины черепа происходит изменение ширины лица, имеется связь между окраской волос и глаз.
Выделение крупных ветвей в рамках больших рас относится к эпохе мезолита. Внутри европеоидной расы выделяются северная и южная ветви, внутри монголоидной — азиатская и американская, негроидный ствол разбивается на африканскую и австралийскую. К этому же времени относится формирование метисныхтипов в смешанных зонах.
Подавляющая часть современных расовых типов сформировалась на протяжении последних двух-трёх тысячелетий.
↑ Ð Ð°Ñ Ñ‹ и их Ð¿Ñ€Ð¾Ð¸Ñ Ñ…Ð¾Ð¶Ð´ÐµÐ½Ð¸Ðµ — Гипермаркет знаний
Согласно одной полярной версии, рас в верхнем палеолите и ранее просто не было, они сформировались уже в голоцене. Широкоизвестна много раз цитировавшаяся концепция В.В. Бунака о существовании в верхнем палеолите "краниологического полиморфизма", выражающегося в наличии разнородных индивидов в пределах каждой отдельной популяции (Бунак, 1959, 1966, 1980). Противоположная версия – расы существовали во все времена, хотя не обязательно были похожи на современные, сейчас мы наблюдаем просто один из вариантов расовой картины. Как ни странно, эта версия может быть связана тоже с именем В.В. Бунака, поскольку именно он обосновал историческую концепцию расы, согласно которой "раса есть историческая категория" (Бунак, 1938).
Сложение рас в более-менее поздние времена на современном уровне может быть обосновано двояко. Во-первых, почти всеми признаваемое африканское моноцентрическое происхождение вида Homo sapiensпредполагает возникновение вида на сравнительно ограниченной территории, хотя бы это и была вся Африка. Таким образом, первоначальные популяцииHomo sapiens наверняка были в расовом отношении довольно однородными, тем более, если учитывать построения генетиков, расчитывающих наличие в прошлом периодов резкого сокращения численности и вариабельности – "бутылочных горлышек". Выход же из Африки, привёдший к появлению всех прочих рас, кроме негроидной, по всем данным, был первоначально осуществлён очень немногочисленной группой людей из Восточной Африки, скорее всего – древними негроидами или предками восточноафриканской расы. Выход доныне точно не датирован, генетики в последнее время обычно называют дату 80 тысяч лет назад, палеоантропологические и археологические данные говорят о достоверном появлении сапиенсов за пределами Африки только ко времени от силы 45 тысяч лет назад или чуть раньше. В любом случае, все внеафриканские расы должны были появиться за эти четыре-пять десятков тысяч лет. Проблема, правда, в том, что даже древнейшие внеафриканские сапиенсы уже заметно различаются и далеко не всегда похожи на негроидов. Может, это следствие расовообразовательных процессов, шедших от 80 до 45 тысяч лет назад – в промежуток, гипотетически предсказанный, но не обоснованный реальными находками. Таким образом, моноцентрическое происхождение вида не даёт строгого обоснования появления рас лишь позже верхнего палеолита, ибо расселение произошло, возможно, до его начала.
Другим образом позднее сложение рас можно обосновать, исходя из представления о популяционной обусловленности рас. Согласно общепризнанному определению, раса – это популяция или (чаще) группа популяций, имеющая общую историю возникновения на определённом ареале и отличающаяся набором биологических наследуемых признаков (или просто – генетическим своеобразием). Из этого можно сделать вывод, что современные так называемые "большие" расы – просто сильно размножившиеся популяции, которым повезло с местом жизни – климатом, ресурсами, соседями (мысль не моя). В глубокой же древности, когда процессы изоляции преобладали, своеобразие малочисленных групп могло сравнительно быстро доходить (хотя бы только из-за генетико-автоматических процессов типа генного дрейфа и эффекта основателя) до таксономического уровня "больших" рас. Впрочем, хронологическая стабильность таких рас должна была бы быть невеликой из-за малой численности. И этих резко различных рас могло быть великое множество даже на довольно ограниченных территориях. Когда мы находим такие различающиеся черепа, мы воспринимаем это как "верхнепалеолитический краниологический полиморфизм". Сложение же современных "больших" рас, имеющих огромный ареал и численность, должно быть связано с получением какими-то из первоначальных мелких популяций неких явных преимуществ. Такие преимущества вряд ли были биологическими, скорее они должны быть социальными и технологическими.
Первое и очевидное, что приходит в голову – появление производящего хозяйства, земледелие, скотоводство, государственное централизованное устройство с хотя бы как-то организованной армией, желательно – металлургия, жизнь в крупных долговременных поселениях уровня селища и выше. А все эти "достижения цивилизации" появляются лишь в голоцене. Группы, получившие перечисленные усовершенствования, могли успешно вытеснять и истреблять много мелких разрозненных популяций с примитивным типом организации, что мы и знаем из истории. Количество рас сокращалось, а расовые признаки "продвинутых" популяций быстро распространялись в пространстве и численности, создавая современную расовую картину. Такой сценарий объясняет позднее – голоценовое – сложение рас и выглядит вроде бы достоверным, но есть НО. Сложность в том, что среди современных рас есть примеры значительной расовой однородности, сложившейся без каких-либо из перечисленных "вершин цивилизации".
Конечно, на первом месте тут австралийские аборигены. До контактов с европейцами они имели самую примитивную культуру на планете (за исключением родственных им тасманийцев) и были в то же время одной из самых однородных рас. Об их однородности лучше всего говорит тот факт, что до сих пор никому не удалось чётко и объективно выделить в австралоидной расе какие-то варианты. Описанные мюррейский и карпентарийский типы, даже при признании их реальности, отличаются друг от друга весьма слабо; лишь третий – барриноидный – тип имеет более явную специфику; однако провести внятную географическую границу между этими типами практически невозможно. Великая расовая однородность австралийских аборигенов, что замечательно, обеспечивалась, кроме отсутствия резких географических барьеров, как раз низким уровнем социальной стратификации. У аборигенов даже не было племён в общепризнанном значении. Брачные связи ограничивались ближайшими соседями, но в целом система брачных контактов охватывала фактически весь континент, способствуя сглаживанию расовых отличий. Барьеры между группами были фактически лишь лингвистическими, а таковые никогда и никого на планете не удерживали от заключения браков (я не хочу сказать, что аборигены никак не отделяли себя от соседних групп, но у них не было племён, каст, богатых и бедных, "высших" и "низших").
Другими примерами широкого распространения довольно расово-однородных популяций, имеющих низкий цивилизационный уровень, могут служить бушмены и готтентоты Южной Африки, меланезийцы, а также индейцы обеих Америк. Конечно, можно возразить, что в Австралии и Африке нет значительных географических барьеров, кои могли бы способствовать изоляции, в Америку люди попали сравнительно недавно и потому расовая дифференциация не зашла далеко, а меланезийцы, как-никак, достигли уровня производящего хозяйства. Однако, существуют примеры обратного рода. В Индии высокая древняя культура и цивилизация скорее способствовали ВОЗНИКНОВЕНИЮ массы расовых вариантов вследствие одной только кастовой изоляции. В отсутствие географических препятствий (более того – в пределах одного населённого пункта!) и потенциально равном социокультурном уровне расы существуют тысячи лет в практически полной изоляции. Таким образом, предположение о связи сложения современной расовой картины с социокультурными, хозяйственными и цивилизационными аспектами демографии, как минимум, сталкивается с массой сложностей и исключений.
Станислав Владимирович Дробышевский
Научный редактор АНТРОПОГЕНЕЗ.РУ, к.б.н., доцент кафедры антропологии биологического факультета МГУ им. Ломоносова
Фрагмент из книги:

ДОСТАЮЩЕЕ ЗВЕНО
Специально для портала "Антропогенез.РУ". Авторский проект С.Дробышевского. Электронная книга даст читателям базовую информацию о том, что известно современной науке о древней родословной человека.
Мнение о том, что расы в том или ином виде существовали всегда, с одной стороны, подтверждается значительными различиями ископаемых черепов, с другой – тоже имеет недостатки. Во-первых, уже несколько раз упоминалось, что зачастую в одном древнем местонахождении мы имеем черепа, которые формально можно отнести к разным расам. Конечно, можно объяснять это индивидуальной изменчивостью, смешанными браками или переходом индивидов из группы в группу, но в таком случае неясно, как же тогда, при такой лёгкости смены группы, сохранялась значительная расовая разница между этими группами?


Черепа представителей разных рас: пигмей Конго, негр, европеец, эскимос, индеец. Государственный Дарвиновский Музей, Москва Фото: А. Соколов
Во-вторых, принимая за действительность "великий африканский исход" 80-40 тысяч лет назад, логично предположить начало формирования большинства рас современного человека лишь с этого времени. Более ранние расы палеоантропов и архантропов, хотя и наверняка существовали, к нашему виду не имеют отношения (конечно, если они не дали вклад в формирование локальных рас в результате метисации, а по новейшим данным генетики, такие метисации вроде бы имели место как минимум дважды – снеандертальцами в Европе и загадочными "денисовцами" где-то по пути в Меланезию).
Отдельный и очень важный вопрос: насколько расы устойчивы во времени? Как быстро они меняются? Ведь, если они меняются очень быстро, мы почти ничего не сможем сказать о сложении рас, исходя из единичных находок, разделённых огромными временными промежутками. Удивительно, но все вероятные варианты – расы весьма устойчивы во времени, расы заметно меняются со временем, расы могут меняться ОЧЕНЬ быстро – встречаются в реальности.
Первый пример – австралийские аборигены. При почти полной изоляции на протяжении около 42-45 (O'Connell et Allen, 1998) или, максимум, 48 тысяч лет (Gillespie, 2002) австралийская раса не претерпела принципиальных изменений. За это грандиозное время лишь несколько в среднем уменьшилась массивность, а может, даже и не уменьшилась. Как ни странно, подобная стабильность может наблюдаться и в условиях, когда такой исключительной изоляции нет. Примерами могут служить дальневосточная и эфиопская расы. На территории центрального Китая и Египта расовый облик населения принципиально не поменялся с неолита – не менее 5 тысяч лет, при том что достоверно известны значительные перемещения населения, неоднократные чужеземные завоевания, резкие изменения численности и прочие неурядицы. Несмотря на все эти исторические сложности, современные китайцы и египтяне отличаются от своих неолитических предков в основном уровнем массивности – меньшим развитием надбровья, меньшими зубами и тому подобными мелочами. Все же ведущие расовые признаки остались, как ни удивительно, прежними.


Айн. Лекционный плакат кафедры антропологии биофака МГУ
Замечательный пример плавного изменения расовых признаков даёт нам история сложения айнской расы, поскольку она протекала в большой изоляции без существенного притока новых групп населения. Судя по краниологическим данным, айны являются прямыми потомками создателей культуры дзёмон (Hanihara, 1998). В свою очередь, дзёмонцы с наибольшей вероятностью восходят к верхнепалеолитическому населению Японских островов (Seguchi, 2004; The Minatogawa man, 1982). Несмотря на существующее разнообразие мнений, можно констатировать, что верхнепалеолитические люди в антропологическом отношении отличались от неолитических дзёмонцев, те, в свою очередь, несколько разнились от эпи-дзёмонцев, а последние заметно отличались от современных айнов, которые в более-менее современном виде известны примерно с XIII века нашей эры. Таким образом, сложение курильской расы заняло в целом 15-20 тысяч лет, в течении которых признаки менялись плавно и постепенно.
Из исторического времени известно несколько примеров быстрого изменения рас, как правило, в результате метисации. Лучше всего изучено становление южносибирской расы: в бронзовом веке степи юга Западной Сибири были населены европеоидами, а в позднем средневековье тут уже жили представители вполне сложившейся южносибирской расы. Глобальные перемещения монголоидного населения с востока на запад начались во II веке нашей эры переселениями хунну, а закончились татаро-монгольским нашествием XIII века нашей эры. Понятно, что и до гуннов монголоиды проникали на запад, что чётко прослеживается по останкам людей пазырыкской и близких ей культур Алтая – с VII-VI веков до нашей эры. После же татаро-монголов метисация шла ещё несколько столетий, так что окончательно сложение южносибирской расы относится к XIV-XVI векам нашей эры. Таким образом, при весьма интенсивном смешении расообразование заняло примерно от 1500 до 2200 лет.
С другой стороны, в современной Латинской Америке интенсивное смешение всех рас планеты, происходившее с момента открытия Америки Колумбом, доныне не привело к уравниванию признаков до состояния монолитной расы, то есть 500 лет для этого оказалось недостаточно. Впрочем, за всё это время шёл постоянный приток новых мигрантов, которые не давали "устаканиться" расовым комплексам. В более локальном масштабе, например, Мексики и Пуэрто-Рико новые расовые типы фактически успели сложиться, хотя доныне они не нашли адекватного места в расовых схемах.
Таким образом, заранее невозможно определить, как могли вести себя расовые комплексы той или иной территории, тем более, что причины стабильности или нестабильности конкретных рас достоверно неизвестны.
Очевидно, что решить все указанные сложности и противоречия можно, лишь тщательно изучая реальные краниологические материалы, а не упражняясь в теоретических построениях. А данные на настоящий момент примерно таковы...
1. Понятие «расы человека»
Расы человека (франц., ед. ч. race) – систематические подразделения внутри вида Человек разумный (Homo Sapiens Sapiens). В основе понятия «раса» лежит биологическое, прежде всего физическое сходство людей и общность населяемой ими территории (ареала) в прошлом или настоящем. Раса характеризуется комплексом наследуемых признаков, к которым относятся цвет кожи, волос, глаз, форма волос, мягких частей лица, черепа, отчасти рост, пропорции тела и др. Но поскольку большинство этих признаков у человека подвержено изменчивости, а между расами происходили и происходят смешения (метисация), конкретный индивид редко обладает всем набором типичных расовых признаков.
2. Большие расы человека
Начиная с 17 века предложено немало различных классификаций человеческих рас. Наиболее часто выделяют три основные, или большие, расы: европеоидная (евразийская, кавказоидная), монголоидная (азиатско-американская) и экваториальная (негро-австралоидная). Европеоидная раса характеризуется светлой кожей (с вариациями от очень светлой, главным образом в Северной Европе, до относительно смуглой в Южной Европе и на Ближнем Востоке), мягкими прямыми или волнистыми волосами, горизонтальным разрезом глаз, умеренно или сильно развитым волосяным покровом на лице и груди у мужчин, заметно выступающим носом, прямым или несколько наклонным лбом. У представителей монголоидной расы цвет кожи варьируется от смуглого до светлого (в основном у североазиатских групп), волосы, как правило, темные, часто жесткие и прямые, выступание носа обычно небольшое, глазная щель имеет косой разрез, значительно развита складка верхнего века и, кроме того, есть складка (эпикантус), прикрывающая внутренний угол глаза; волосяной покров слабый. Экваториальная, или негро-австралоидная раса отличается темной пигментацией кожи, волос и глаз, курчавыми или широковолнистыми (австралийцы) волосами; нос обычно широкий, маловыступающий, нижняя часть лица выдается. Генетически все расы представлены различными аутосомными компонентами, а в тех случаях когда раса смешанного происхождения, то таких компонент обычно выделяется несколько, каждая из которых различного происхождения.
3. Малые расы и их географическое распространение
Каждая большая раса подразделяется на малые расы, или антропологические типы. Внутри европеоидной расы различают атланто-балтийскую, беломорско-балтийскую, среднеевропейскую, балкано-кавказскую и индо-средиземноморскую малые расы. Ныне европеоиды населяют фактически всю обитаемую сушу, но до середины 15 века — начала великих географических открытий — основной их ареал включал Европу и частично Северную Африку, Переднюю и Среднюю Азию и Северную Индию. В современной Европе представлены все малые расы, но численно преобладает среднеевропейский вариант (часто встречается у австрийцев, немцев, чехов, словаков, поляков, русских, украинцев); в целом же ее население очень смешанно, особенно в городах, вследствие переселений, метисации и притока мигрантов из других регионов Земли. Внутри монголоидной расы обычно выделяют дальневосточную, южноазиатскую, североазиатскую, арктическую и американскую малые расы, причем последняя иногда рассматривается и как отдельная большая раса. Монголоиды заселили все климатогеографические зоны (Северная, Центральная, Восточная и Юго-Восточная Азия, острова Тихого океана, Мадагаскар, Северная и Южная Америка). Для современной Азии характерно большое разнообразие антропологических типов, но преобладают по численности различные монголоидные и европеоидные группы. Среди монголоидов наиболее распространены дальневосточная (китайцы, японцы, корейцы) и южноазиатская (малайцы, яванцы, зондцы) малые расы, среди европеоидов — индо-средиземноморская. В Америке коренное население (индейцы) составляет меньшинство, по сравнению с различными европеоидными антропологическими типами и группами населения представителей всех трех больших рас.
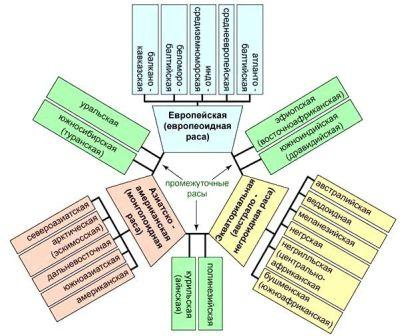 Рис.
Схема антропологического состава
народов мира (малые расы, выделяемые
внутри больших, отличаются друг от
друга не столь существенными
признаками).
Экваториальная,
или негро-австралоидная, раса включает
три малые расы африканских негроидов
(негрская, или негроидная, бушменская
и негрилльская) и столько же океанийских
австралоидов (австралийская, или
австралоидная, раса, которую в некоторых
классификациях выделяют в самостоятельную
большую расу, а также меланезийская и
веддоидная). Ареал экваториальной расы
не сплошной: он охватывает большую
часть Африки, Австралию, Меланезию,
Новую Гвинею, отчасти Индонезию. В
Африке численно преобладает негрская
малая раса, на севере и юге континента
значителен удельный вес европеоидного
населения.
В
Австралии коренное население составляет
меньшинство по отношению к мигрантам
из Европы и Индии, довольно многочисленны
и представители дальневосточной расы
(японцы, китайцы). В Индонезии преобладает
южноазиатская раса.
Наряду
с перечисленным выше существуют расы
с менее определенным положением,
образовавшиеся в результате длительного
смешения населения отдельных регионов,
например, лапаноидная и уральская расы,
совмещающие черты европеоидов и
монголоидов в той или иной степени, а
также эфиопская раса — промежуточная
между экваториальной и европеоидной
расами.
Рис.
Схема антропологического состава
народов мира (малые расы, выделяемые
внутри больших, отличаются друг от
друга не столь существенными
признаками).
Экваториальная,
или негро-австралоидная, раса включает
три малые расы африканских негроидов
(негрская, или негроидная, бушменская
и негрилльская) и столько же океанийских
австралоидов (австралийская, или
австралоидная, раса, которую в некоторых
классификациях выделяют в самостоятельную
большую расу, а также меланезийская и
веддоидная). Ареал экваториальной расы
не сплошной: он охватывает большую
часть Африки, Австралию, Меланезию,
Новую Гвинею, отчасти Индонезию. В
Африке численно преобладает негрская
малая раса, на севере и юге континента
значителен удельный вес европеоидного
населения.
В
Австралии коренное население составляет
меньшинство по отношению к мигрантам
из Европы и Индии, довольно многочисленны
и представители дальневосточной расы
(японцы, китайцы). В Индонезии преобладает
южноазиатская раса.
Наряду
с перечисленным выше существуют расы
с менее определенным положением,
образовавшиеся в результате длительного
смешения населения отдельных регионов,
например, лапаноидная и уральская расы,
совмещающие черты европеоидов и
монголоидов в той или иной степени, а
также эфиопская раса — промежуточная
между экваториальной и европеоидной
расами.
4. Происхождение человеческих рас
Расы человека, по-видимому, появились сравнительно недавно. Согласно одной из схем, основанной на данных молекулярной биологии и генетики, разделение на два больших расовых ствола — негроидный и европеоидно-монголоидный — произошло скорее всего около 80 тысяч лет назад, а первичная дифференциация прото-европеоидов и прото-монголоидов — около 40-45 тысяч лет назад. Большие расы в основном формировались под влиянием природных и социально-экономических условий в ходе внутривидовой дифференциации уже сложившегося человека разумного, начиная с эпохи палеолита и мезолита, но распространились главным образом уже в неолите и позже. Европеоидный тип установился массово с неолита, хотя многие отдельные его черты прослеживаются в позднем или даже среднем палеолите. Фактически отсутствуют достоверные свидетельства присутствия сложившихся монголоидов в Восточной Азии в донеолитическую эпоху, хотя в Северной Азии они, возможно, существовали уже в позднем палеолите. В Америке предки индейцев не были окончательно сложившимися монголоидами. Также и Австралия заселялась еще «нейтральными» в расовом отношении неоантропами. Существуют две основные гипотезы происхождения человеческих рас — полицентризма и моноцентризма. Согласно теории полицентризма, современные расы человека возникли в результате длительной параллельной эволюции нескольких филетических линий на разных материках: европеоидная в Европе, негроидная в Африке, монголоидная в Центральной и Восточной Азии, австралоидная в Австралии. Однако если эволюция расовых комплексов и шла параллельно на разных континентах, она не могла быть полностью независимой, поскольку древние проторасы должны были скрещиваться на границах своих ареалов и обмениваться генетической информацией. В ряде областей сформировались промежуточные малые расы, характеризующиеся смешением признаков разных больших рас уже в древности. Так, промежуточное положение между европеоидной и монголоидной расами занимают южносибирская и уральская малые расы, между европеоидной и негроидной — эфиопская и т. д. С позиций моноцентризма современные человеческие расы сформировались относительно поздно, 30-35 тыс. лет назад, в процессе расселения неоантропов из области их возникновения. При этом также допускается возможность скрещивания (хотя бы ограниченного) неоантропов во время их экспансии с вытесняемыми популяциями палеоантропов (как процесса интрогрессивной межвидовой гибридизации) с проникновением аллелей последних в генофонды популяций неоантропов. Это также могло способствовать расовой дифференциации и устойчивости некоторых фенотипических признаков (подобных лопатообразным резцам монголоидов) в центрах расообразования. Существуют и компромиссные между моно- и полицентризмом концепции, допускающие расхождение филетических линий, ведущих к разным большим расам, на различных уровнях (стадиях) антропогенеза: например, более близких друг к другу европеоидов и негроидов уже на стадии неоантропов с первоначальным развитием их предкового ствола в западной части Старого Света, тогда как еще на стадии палеоантропов могла обособиться восточная ветвь — монголоиды и, может быть, австралоиды, хотя по некоторым отдельным признакам европеоиды имеют общие признаки и с австролоидами. Большие расы человека занимают обширные территории, охватывающие народы, которые различаются по уровню экономического развития, культуре, языку. Нет сколько-нибудь четких совпадений понятий «раса» и «этнос» (народ, нация, национальность). В то же время есть примеры антропологических типов (малые, а иногда и большие расы), которые соответствуют одному или нескольким близким этносам, например, лапаноидная раса и саамы. Гораздо чаще, однако, наблюдается обратное: один антропологический тип широко распространен среди многих этносов, как, например, в коренном населении Америки или у народов Северной Европы. Вообще все крупные народы, как правило, неоднородны в антропологическом отношении. Нет также совпадения между расами и языковыми группами — последние возникали позже, чем расы. Так, среди тюркоязычных народов имеются представители как европеоидов (азербайджанцы), так и монголоидов (якуты). Термин «расы» неприменим к языковым семьям — например, нужно говорить не о «славянской расе», а о группе родственных народов, говорящих на славянских языках.
5. Расы и расизм
Многие расовые признаки имеют приспособительное значение. Например, у представителей экваториальной расы темная пигментация кожи защищает от обжигающего действия ультрафиолетовых лучей, а вытянутые пропорции тела увеличивают отношение поверхности тела к его объему и тем самым облегчают терморегуляцию в условиях жаркого климата. Однако расовые признаки не являются определяющими для существования человека, поэтому они ни в коей мере не свидетельствуют о каком-либо биологическом или интеллектуальном превосходстве или, напротив, неполноценности той или иной расы. Все расы находятся на одном и том же уровне эволюционного развития и характеризуются одинаковыми видовыми особенностями. Поэтому концепции о якобы неравноценности человеческих рас в физическом и психическом отношениях (расизм), выдвигаемые с середины 19 века, научно несостоятельны. Расизм имеет отчетливые социальные корни и всегда использовался как оправдание для насильственного захвата земель и дискриминации коренных народов. Расисты обычно игнорируют тот факт, что различия между достижениями разных народов полностью объясняются историей их культур, зависящей от внешних факторов, от их исторически меняющейся роли. Достаточно сравнить уровень культурного развития населения Северной Европы в наши дни и в эпоху великих цивилизаций прошлого в Месопотамии, Египте, долине Инда.
Заключение
Расы человека – систематические подразделения внутри вида Человек разумный (Homo sapiens). В основе понятия «раса» лежит биологическое, прежде всего физическое сходство людей и общность населяемой ими территории (ареала) в прошлом или настоящем. Наиболее часто по признакам выделяют три основные, или большие, расы: европеоидная (евразийская, кавказоидная), монголоидная (азиатско-американская) и экваториальная (негро-австралоидная). Каждая большая раса подразделяется на малые расы, или антропологические типы. Существуют две основные гипотезы происхождения человеческих рас — полицентризма и моноцентризма. Согласно теории полицентризма, современные расы человека возникли в результате длительной параллельной эволюции нескольких филетических линий на разных материках: европеоидная в Европе, негроидная в Африке, монголоидная в Центральной и Восточной Азии, австралоидная в Австралии. С позиций моноцентризма современные человеческие расы сформировались относительно поздно, 20—35 тыс. лет назад, в процессе расселения неоантропов из области их возникновения. Существуют и компромиссные между моно- и полицентризмом концепции, допускающие расхождение филетических линий, ведущих к разным большим расам, на различных уровнях (стадиях) антропогенеза. Большие расы человека занимают обширные территории, охватывающие народы, которые различаются по уровню экономического развития, культуре, языку. Нет сколько-нибудь четких совпадений понятий «раса» и «этнос» (народ, нация, национальность). Вообще все крупные народы, как правило, неоднородны в антропологическом отношении. Нет также совпадения между расами и языковыми группами — последние возникали позже, чем расы. Многие расовые признаки имеют приспособительное значение и не являются определяющими для существования человека, поэтому они ни в коей мере не свидетельствуют о каком-либо биологическом или интеллектуальном превосходстве или, напротив, неполноценности той или иной расы. Все расы находятся на одном и том же уровне эволюционного развития и характеризуются одинаковыми видовыми особенностями. Поэтому концепции о якобы неравноценности человеческих рас в физическом и психическом отношениях (расизм), выдвигаемые с середины 19 века, научно несостоятельны. Расизм имеет отчетливые социальные корни и всегда использовался как оправдание для насильственного захвата земель и дискриминации коренных народов. Расисты обычно игнорируют тот факт, что различия между достижениями разных народов полностью объясняются историей их культур, зависящей от внешних факторов, от их исторически меняющейся роли. На генетическом уровне также прослеживаются четкие корреляции между аутосомными компонентами и расовой принадлежностью. Скачать подробное филогенетическое дерево Y гаплогрупп Скачать карту путей миграции Y-гаплогрупп