

Генетика
Предмет, задачи и методы генетики
Генетика—- наука о наследственности и изменчивости живых организмов и методах управления ими. В ее основу легли закономерности наследственности, установленные выдающимся чешским ученым Грегором Менделем (1822—1884) при скрещивании различных сортов гороха.
Наследственность — это неотъемлемое свойство всех живых существ сохранять и передавать в ряду поколений характерные для вида или популяции особенности строения, функционирования и развития. Наследственность обеспечивает постоянство и многообразие форм жизни и лежит в основе передачи наследственных задатков, ответственных за формирование признаков и свойств организма. Благодаря наследственности некоторые виды (например, кистеперая рыба латимерия, жившая в девонском периоде) оставались почти неизменными на протяжении сотен миллионов лет, воспроизводя за это время огромное количество поколений.
В то же время в природе существуют различия между особями как разных видов, так и одного и того же вида, сорта, породы и т. д. Это свидетельствует о том, что наследственность неразрывно связана с изменчивостью.
Изменчивость — способность организмов в процессе онтогенеза приобретать новые признаки и терять старые. Изменчивость выражается в том, что в любом поколении отдельные
особи чем-то отличаются и друг от друга, и от своих родителей. Причиной этого является то, что признаки и свойства любого организма есть результат взаимодействия двух факторов: наследственной информации, полученной от родителей, и конкретных условий внешней среды, в которых шло индивидуальное развитие каждой особи. Поскольку условия среды никогда не бывают одинаковыми даже для особей одного вида или сорта (породы), становится понятным, почему организмы, имеющие одинаковые генотипы, часто заметно отличаются друг от друга по фенотипу, т. е. по внешним признакам.
Таким образом, наследственность, будучи консервативной, обеспечивает сохранение признаков и свойств организмов на протяжении многих поколений, а изменчивость обусловливает формирование новых признаков в результате изменения генетической информации или условий внешней среды.
Задачи генетики вытекают из установленных общих закономерностей наследственности и изменчивости. К этим задачам относятся исследования: 1) механизмов хранения и передачи генетической информации от родительских форм к дочерним; 2) механизма реализации этой информации в виде признаков и свойств организмов в процессе их индивидуального развития под контролем генов и влиянием условий внешней среды; 3) типов, причин и механизмов изменчивости всех живых существ; 4) взаимосвязи процессов наследственности, изменчивости и отбора как движущих факторов эволюции органического мира.
Генетика является также основой для решения ряда важнейших практических задач. К ним относятся: 1) выбор наиболее эффективных типов гибридизации и способов отбора; 2) управление развитием наследственных признаков с целью получения наиболее значимых длячеловека результатов; 3) искусственное получение наследственно измененных форм живых организмов; 4) разработка мероприятий по защите живой природы от вредных мутагенных воздействий различных факторов внешней среды и методов борьбы с наследственными болезнямичеловека, вредителями сельскохозяйственных растений и животных; 5) разработка методов генетической инженерии с целью получения высокоэффективных продуцентов биологически активных соединений, а также для создания принципиально новых технологий в селекции микроорганизмов, растений и животных.
При изучении наследственности и изменчивости на разных уровнях организации живой материи (молекулярный, клеточный,
организменный, популяционный) в генетике используют разнообразные методы современной биологии: гибридологический, цитогенетический, биохимический, генеалогический, близнецовый, мутационный и др. Однако среди множества методов изучения закономерностей наследственности центральное место принадлежит гибридологическому методу. Суть его заключается в гибридизации (скрещивании) организмов, отличающихся друг от друга по одному или нескольким признакам, с последующим анализом потомства. Этот метод позволяет анализировать закономерности наследования и изменчивости отдельных признаков и свойств организма при половом размножении, а также изменчивость генов и их комбинирование.
При микроскопическом анализе хромосом прежде всего видны различия их по форме и величине. Строение каждой хромосомы сугубо индивидуальное. Можно заметить также, что хромосомы обладают общимиморфологическими признаками. Они состоят из двух нитей — хроматид, расположенных параллельно и соединенных между собой в одной точке, названной центромерой или первичной перетяжкой. На некоторыххромосомах можно видеть и вторичную перетяжку. Она является характерным признаком, позволяющим идентифицировать отдельные хромосомы в клетке. Если вторичная перетяжка расположена близко к концухромосомы, то дистальный участок, ограниченный ею, называют спутником. Хромосомы, содержащие спутник, обозначаются как АТ-хромосомы. На некоторых из них в тело-фазе происходит образование ядрышек. Концевые участки хромосом имеют особую структуру и называются теломерами. Теломерные районы обладают определенной полярностью, препятствующей их соединению друг с другом при разрывах или со свободными концами хромосом.
Участок хрома-тиды (хромосомы) от теломеры до центромеры называют плечом хромосомы. Каждаяхромосома имеет два плеча. В зависимости от соотношения длин плеч выделяют три типа хромосом: 1) мета-центрические (равноплечие); 2) субметацентрические (неравноплечие); 3) акроцентрические, у которых одно плечо очень короткое и не всегда четко различимо. На Парижской конференции по стандартизации кариотипов вместо морфологических терминов «метацентрики» или «акро-центрики» в связи с разработкой новых методов получения «полосатых»хромосом предложена символика, в которой всем хромосомам набора присваивается ранг (порядковый номер) по порядку убывания величины и в обоих плечах каждой хромосомы (р — короткое плечо, q — длинное плечо) нумеруются участки плеч и полосы в каждом участке по направлению от центромеры. Такая система обозначений позволяет детально описывать аномалии хромосом. Наряду с расположением центромеры, наличием вторичной перетяжки и спутника важное значение для определения отдельных хромосом имеет их длина.
Для каждой хромосомы определенного набора длина ее остается относительно постоянной. Измерениехромосом необходимо для изучения их изменчивости в онтогенезе в связи с болезнями, аномалиями, нарушением воспроизводительной функции. Тонкое строение хромосом. Химический анализ структуры хромосом показал наличие в них двух основных компонентов: дезоксирибонуклеиновой кислоты (ДНК) и белков типа гистонов и протомите (в половых клетках). Исследования тонкой субмолекулярной структуры хромосом привели ученых к выводу, что каждая хроматида содержит одну нить — хромонему. Каждая хро-монема состоит из одной молекулы ДНК. Структурной основой хроматиды является тяж белковой природы. Хромонема уложена в хроматиде в форму, близкую к спирали. Доказательства этого предположения были получены, в частности, при изучении мельчайших обменных частиц сестринских хроматид, которые располагались поперек хромосомы.
Уровни компактизации хроматина
Нуклеосомиая нить. Этот уровень организации хроматина обеспечивается четырьмя видами нуклеосомных гистонов: Н2А, Н2В, НЗ, Н4. Они образуют напоминающие по форме шайбу белковые тела — коры, состоящие из восьми молекул (по две молекулы каждого вида гистонов) (рис. 3.46).
Молекула ДНК комплектируется с белковыми корами, спирально накручиваясь на них. При этом в контакте с каждым кором оказывается участок ДНК, состоящий из 146 пар нуклеотидов (п.н.). Свободные от контакта с белковыми телами участки ДНК называютсвязующими или линкерными. Они включают от 15 до 100 п.н. (в среднем 60 п.н.) в зависимости от типа клетки.
Отрезок молекулы ДНК длиной около 200 п. н. вместе с белковым кором составляетнуклеосому. Благодаря такой организации в основе структуры хроматина лежит нить, представляющая собой цепочку повторяющихся единиц — нуклеосом (рис. 3.46, Б). В связи с этим геном человека, состоящий из 3 · 109 п. н., представлен двойной спиралью ДНК, упакованной в 1,5 · 107 нуклеосом.
Вдоль нуклеосомной нити, напоминающей цепочку бус, имеются области ДНК, свободные от белковых тел. Эти области, расположенные с интервалами в несколько тысяч пар нуклеотидов, играют важную роль в дальнейшей упаковке хроматина, так как содержат нуклеотидные последовательности, специфически узнаваемые различными негистоновыми белками.
В результате нуклеосомной организации хроматина двойная спираль ДНК диаметром 2 нм приобретает диаметр 10—11 нм.
Хроматиновая фибрилла. Дальнейшая компактизация нуклеосомной нити обеспечивается пистоном HI, который, соединяясь с линкерной ДНК и двумя соседними белковыми телами, сближает их друг с другом. В результате образуется более компактная структура, построенная, возможно, по типу соленоида. Такая Хроматиновая фибрилла, называемая также элементарной, имеет диаметр 20—30 нм (рис. 3.47).
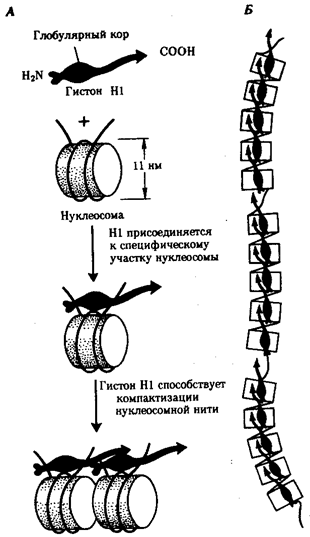

Рис. 3.47. Хроматиновая фибрилла диаметром 20—30 нм.А — соединение соседних нуклеосом с помощью гистона HI; Б — цепочка, образуемая нуклеосомами разделенными участками ДНК, свободными от белковых тел; В — возможная модель упаковки ДНК в хроматиновой фибрилле в виде соленоида
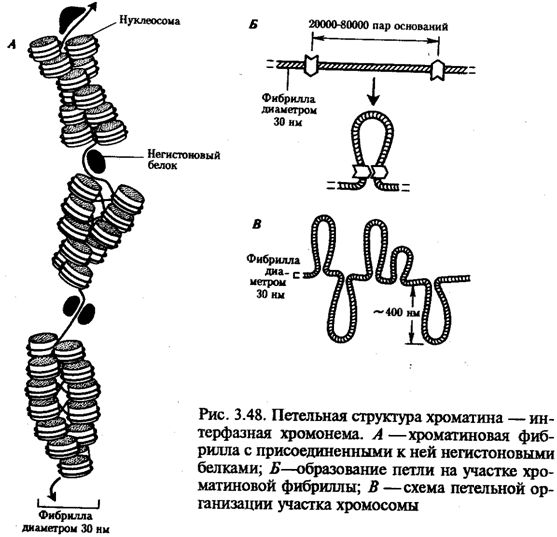
Интерфазная хромонема. Следующий уровень структурной организации генетического материала обусловлен укладкой хроматиновой фибриллы в петли. В их образовании, по-видимому, принимают участие негистоновые белки, которые способны узнавать специфические нуклеотидные последовательности вненуклеосомной ДНК, отдаленные друг от друга на расстояние в несколько тысяч пар нуклеотидов. Эти белки сближают указанные участки с образованием петель из расположенных между ними фрагментов хроматиновой фибриллы (рис. 3.48). Участок ДНК, соответствующий одной петле, содержит от 20 000 до 80 000 п. н. Возможно, каждая петля является функциональной единицей генома. В результате такой упаковки Хроматиновая фибрилла диаметром 20—30 нм преобразуется в структуру диаметром 100—200 нм, называемую интерфазной хромонемой.
Отдельные участки интерфазной хромонемы подвергаются дальнейшей компактизации, образуя структурные блоки, объединяющие соседние петли с одинаковой организацией (рис. 3.49). Они выявляются в интерфазном ядре в виде глыбок хроматина. Возможно, существование таких структурных блоков обусловливает картину неравномерного распределения некоторых красителей в метафазных хромосомах, что используют в цитогенетических исследованиях (см. разд. 3.5.2.3 и 6.4.3.6).
Неодинаковая степень компактизации разных участков интерфазных хромосом имеет большое функциональное значение. В зависимости от состояния хроматина выделяютэухроматиновые участки хромосом, отличающиеся меньшей плотностью упаковки в неделящихся клетках и потенциально транскрибируемые, и гетерохроматиновые участки, характеризующиеся компактной организацией и генетической инертностью. В их пределах транскрипции биологической информации не происходит.
гетерохроматин и эухроматин
эухроматин, активный хроматин — участки хроматина, сохраняющие деспирализованное состояние элементарных дезоксирибонуклеопротеидных нитей (ДНП) в покоящемся ядре, т. е. в интерфазе (в отличие от других участков, сохраняющих спирализованное состояние — гетерохроматина).
Эухроматин отличается от гетерохроматина также способностью к интенсивному синтезу рибонуклеиновой кислоты (РНК) и большим содержанием негистоновых белков. В нём, помимо ДНП, имеются рибонуклеопротеидные частицы (РНП-гранулы) диаметром 200—500, которые служат для завершения соГетерохроматин — участки хроматина, находящиеся в течение клеточного цикла в конденсированном (компактном) состоянии. Особенностью гетерохроматиновой ДНК является крайне низкая транскрибируемость.зревания РНК и переноса ее в цитоплазму. Эухроматин содержит большинство структурных генов организма.
понятия о геноме и кариотипе.морфология хромосом.использование кариологии и селекции
Гено́м — совокупность наследственного материала, заключенного в клетке организма[1]. Геном содержит биологическую информацию, необходимую для построения и поддержания организма. Большинство геномов, в том числе геном человека и геномы всех остальных клеточных форм жизни, построены из ДНК, однако некоторые вирусы имеют геномы из РНК.Существует также и другое определение термина «геном», в котором под геномом понимают совокупность генетического материала гаплоидного набора хромосом данного вида.У человека (Homo sapiens) геном состоит из 23 пар хромосом, находящихся в ядре, а также митохондриальной ДНК. Двадцать две аутосомы, две половые хромосомы Х и Y, а также митохондриальная ДНК человека содержат вместе примерно 3,1 млрдпар оснований. Кариоти́п — совокупность признаков (число, размеры, форма и т. д.) полного набора хромосом, присущая клеткам данного биологического вида (видовой кариотип), данного организма (индивидуальный кариотип) или линии (клона) клеток. Кариотипом иногда также называют и визуальное представление полного хромосомного набора (кариограммы)
Морфология хромосом
Митотическая суперкомпактизация хроматина делает возможным изучение внешнего вида хромосом с помощью световой микроскопии. В первой половине митоза они состоят из двух хроматид, соединенных между собой в области первичной перетяжки (центромеры иликинетохора) особым образом организованного участка хромосомы, общего для обеих сестринских хроматид. Во второй половине митоза происходит отделение хроматид друг от друга. Из них образуются однонитчатые дочерние хромосомы, распределяющиеся между дочерними клетками.
В зависимости от места положения центромеры и длины плеч, расположенных по обе стороны от нее, различают несколько форм хромосом: равноплечие, или метацентрические (с центромерой посередине), неравноплечие, или субметацентрические (с центромерой, сдвинутой к одному из концов), палочковидные, или акроцентрические (с центромерой, расположенной практически на конце хромосомы), и точковые —очень небольшие, форму которых трудно определить (рис. 3.52). При рутинных методах окраски хромосом они различаются по форме и соотносительным размерам. При использовании методик дифференциальной окраски выявляется неодинаковая флуоресценция или распределение красителя по длине хромосомы, строго специфические для каждой отдельной хромосомы и ее гомолога (рис. 3.53).
Таким образом, каждая хромосома индивидуальна не только по заключенному в ней набору генов, но и по морфологии и характеру дифференциального окрашивания.
Кариология (от лат. «карио» и «логия») — раздел цитологии, занимающийся изучением строения и функций ядра клетки, как в целом, так и его структур:
хромосом,
ядерной оболочки
ядрышка
и т. д.
Селе́кция (лат. selectio - выбирать) — наука о создании новых и улучшении существующих пород животных, сортов растений,штаммов микроорганизмов. Селекцией называют также отрасль сельского хозяйства, занимающуюся выведением новыхсортов и гибридов сельскохозяйственных культур и пород животных.
Передача наследственной информации при бесполом размножении.Митотический цикл. Фазы митоза.Амитоз.Эндомитоз
Первый тип - это деление. Делением размножаются одноклеточные организмы: каждая особь при этом делится на две или большее число клеток, которые называются дочерними, они идентичны родительской клетке. Перед делением клетки происходит репликация ДНК, а у эукариот - деление и ядра. В большинстве случаев происходит бинарное деление, при котором образуются две идентичные клетки. Таким образом делятся бактерии, многие простейшие, например амеба, и некоторые одноклеточные водоросли, например эвглена. При подходящих условиях это приводит к быстрому росту популяции. Множественное деление, при котором следом за рядом повторных делений клеточного ядра происходит деление самой клетки на огромное множество дочерних клеток, можно наблюдать у споровиков - это группа простейших, и к ним относится, например, возбудитель малярии Plasmodium. Стадия, на которой происходит множественное деление, называется шизонтом, а сам этот процесс - шизогонией. У Plasmodium шизогония следует за заражением хозяина, когда паразит проникает в печень. В результате этого получается сразу около тысячи дочерних клеток, каждая из которых способна инвазировать эритроцит и произвести путем шизогонии еще 24 дочерние клетки. Такая высокая плодовитость компенсирует большие потери из-за трудностей успешной передачи паразита отдельного хозяина другому, а именно от человека организму-переносчику, т. е. малярийному комару, и в обратном направлении
Второй тип бесполого размножения - это образование спор, или споруляция. Спора - это одноклеточная репродуктивная единица обычно микроскопических размеров, состоящая из небольшого количества цитоплазмы и ядра. Образование спор можно наблюдать у бактерий, простейших, у представителей всех групп зеленых растений и всех групп грибов. Споры могут быть различными по своему типу и функции и часто образуются в специальных структурах. Например, у Rhiropus и Dryoptenis споры образуются в спорангиях; микроспоры (или пальцевые зерна) или мегаспоры (или зародышевые мешки) семенных растений образуются в особых спораниях, которые носят названия «пыльцевой мешок» и «семязачаток». Очень часто споры образуются в больших количествах, но они имеют ничтожный вес, и это облегчает их распространение ветром, а также животными, но главным образом - насекомыми. Из-за своих маленьких размеров спора обычно содержит лишь минимальные запасы питательных веществ; из-за того, что многие споры не попадают в подходящее место для прорастания, их потери очень велики. Главное достоинство таких спор заключается в возможности быстрого размножения и расселения видов, в особенности это касается грибов. Споры бактерий служат не для размножения, а для того, чтобы выжить при неблагоприятных условиях, потому что каждая бактерия образует только одну спору. Бактериальные споры относятся к числу наиболее устойчивых спор. Так, например, они очень часто выдерживают обработку сильными дезинфицирующими средствами и кипячением в воде. Важно отметить, что один организм может производить споры более чем одного типа; например, Rhiropus образует половые и бесполовые споры, а высшие растения производят бесполым путем микроспоры и мегаспоры Третий тип бесполого размножения - это почкование. Почкованием называют одну из форм бесполого размножения, при которой новая особь образуется в виде выроста (или почки) на теле родительской особи, а затем отделяется от нее, при этом превращаясь в самостоятельный организм, который совершенно идентичен родительскому организму. Почкование встречается у различных групп организмов, особенно у кишечнополостных, например у гидры, и у одноклеточных грибов, таких, как дрожжи. В последнем случае почкование отличается от делений (которые тоже наблюдаются у дрожжей) тем, что две образующиеся части имеют разные размеры. Необычная форма почкования описана у суккулентного растения Bryophyllum - ксерофита, часто выращиваемого в качестве декоративного комнатного растения: по краям его листьев развиваются маленькие, миниатюрные растеньица, снабженные крошечными корешками; эти так называемые «почки» в конце концов отпадают и начинают существовать как самостоятельные растения. Четвертый тип бесполого размножения - размножение фрагментами, или фрагментация. Фрагментацией называют разделение особи на две или несколько частей, каждая из которых растет и образует новую особь. Фрагментация происходит, например, у нитчатых водорослей, таких, как спирогира. Нить спирогиры может разорваться на две части в любом месте. Фрагментация наблюдается также у некоторых низших животных, которые, в отличие от более организованных форм, сохраняют значительную способность к регенерации из относительно слабо регенерированных клеток. Например, тело намертин (это группа примитивных червей, главным образом - морских) особенно легко разрывается на много частей, каждая из которых может дать в результате регенерации новую особь. В этом случае регенерация - это процесс нормальный и регулируемый; но, несмотря на это, у некоторых животных (например, у морских звезд) восстановление из отдельных частей происходит только после случайной фрагментации. Животные, способные к регенерации, служат объектами для экспериментального изучения этого процесса; часто при этом используют свободно живущего червя планарию. пятый тип бесполого размножения - вегетативное. Вегетативное размножение представляет собой одну из форм бесполого размножения, при котором от растения отделяется относительно большая, обычно дифференцированная, часть и развивается в самостоятельное растение. По существу, вегетативное размножение сходно с почкованием. Нередко растения образуют структуры, специально предназначенные для этой цели: луковицы, клубне-луковицы, корневища, столоны и клубни. Некоторые из этих структур служат также для запасания питательных веществ, что позволяет растению переживать зиму и давать в следующем году цветки и плоды (это двулетние растения) или выживать в течение ряда лет (это многолетние растения). К таким органам, они называются зимующие, относятся луковицы, клубне-луковицы, корневища и клубни. Зимующими органами могут быть также стебли, корни или целые побеги (почки): во всех случаях содержащиеся в них питательные вещества создаются главным образом в процессе фотосинтеза, происходящего в листьях текущего года. Образовавшиеся питательные вещества переносят в запасающий орган, а затем обычно превращаются в резервный материал, например в крахмал. При наступлении неблагоприятных условий подземные части растений отмирают, а подземный зимующий орган переходит в состояние покоя. В начале следующего вегетативного периода запасы питательных веществ мобилизуются с помощью ферментов: почки пробуждаются и в них начинаются процессы активного роста и развития за счет запасенных питательных веществ. Если прорастает больше одной почки, то можно считать, что размножение осуществилось. Последовательность этих событий очень тесно связана со сменой времен года, потому что она регулируется такими внешними факторами, как длина светового дня (это фотопериод) и температура. Глубокое влияние этих факторов на рост и развитие было давно доказано.
Митотический цикл
совокупность процессов, в результате которых из одной клетки образуются две новые. М. ц. охватывает период Митоза и часть интерфазы (См. Интерфаза). — периода между делениями, когда происходит подготовка к следующему митозу. М. ц. — часть жизненного цикла клетки; в быстро делящихся клеточных популяциях (например, у бластомеров дробящегося яйца) М. ц. почти совпадает с жизненным циклом клетки.
Различают следующие четыре фазы митоза: профаза, метафаза, анафаза и телофаза. . Впрофазе хорошо видны центриоли — образования, находящиеся в клеточном центре и играющие роль в делении дочерних хромосом животных. (Напомним, что у высших растений нет центриолей в клеточном центре, который организует деление хромосом). Мы же рассмотрим митоз на примере животной клетки, поскольку присутствие центриоли делает процесс деления хромосом более наглядным. Центриоли делятся и расходятся к разным полюсам клетки. От центриолей протягиваются микротрубочки, образующие нити веретена деления, которое регулирует расхождение хромосом к полюсам делящейся клетки. В конце профазы ядерная оболочка распадается, ядрышко постепенно исчезает, хромосомы спирализуются и в результате этого укорачиваются и утолщаются, и их уже можно наблюдать в световой микроскоп. Еще лучше они видны на следующей стадии митоза — метафазе. В метафазе хромосомы располагаются в экваториальной плоскости клетки. При этом хорошо видно, что каждая хромосома, состоящая из двух хроматид, имеет перетяжку — центромеру. Хромосомы своими центромерами прикрепляются у нити веретена деления. После деления центромеры каждая хроматида становится самостоятельной дочерней хромосомой. Затем наступает следующая стадия митоза — анафаза, во время которой дочерние хромосомы (хроматиды одной хромосомы) расходятся к разным полюсам клетки. Следующая стадия деления клетки — телофаза. Она начинается после того, как дочерние хромосомы, состоящие из одной хроматиды, достигли полюсов клетки. На этой стадии хромосомы вновь деспирализуются и приобретают такой же вид, какой они имели до начала деления клетки в интерфазе (длинные тонкие нити). Вокруг них возникает ядерная оболочка, а в ядре формируется ядрышко, в котором синтезируются рибосомы. В процессе деления цитоплазмы все органоиды (митохондрии, комплекс Гольджи, рибосомы и др.) распределяются между дочерними клетками более или менее равномерно. Таким образом, в результате митоза из одной клетки получаются две, каждая из которых имеет характерное для данного вида организма число и форму хромосом, а следовательно, постоянное количество ДНК. Весь процесс митоза занимает в среднем 1-2 ч. Продолжительность его несколько различна для разных видов клеток. Зависит он также от условий внешней среды (температуры, светового режима и других показателей). Биологическое значение митоза заключается в том, что он обеспечивает постоянство числа хромосом во всех клетках организма. Все соматические клетки образуются в результате митотического деления, что обеспечивает рост организма. В процессе митоза происходит распределение веществ хромосом материнской клетки строго поровну между возникающими из нее двумя дочерними клетками. В результате митоза все клетки организма получают одну и ту же генетическую информацию. Амитоз
прямое деление ядра, один из способов деления ядра у простейших, в растительных и животных клетках. А. впервые был описан немецким биологом Р. Ремаком (1841); термин предложен гистологом В. Флеммингом (1882). При А., в отличие от Митоза, или непрямого деления ядра, ядерная оболочка и ядрышки не разрушаются, веретено деления в ядре не образуется, хромосомы остаются в рабочем (деспирализованном) состоянии, ядро или перешнуровывается или в нём, внешне неизменном, появляется перегородка; деления тела клетки — цитотомии (См. Цитотомия), как правило, не происходит (рис.); обычно А. не обеспечивает равномерного деления ядра и отдельных его компонентов.
ЭНДОМИТОЗ
(от эндо... и митоз), удвоение числа хромосом внутри ядерной оболочки без разрушения ядрышка и без образования веретена деления клетки. Ранее Э. считали способом образования полиплоидных и политенных ядер, однако позднее было установлено, что полиплоидия — результат неоконченных обычных митозов, а политения — повторяющихся циклов синтеза ДНК без митоза.
Мейоз {греч. meiosis — уменьшение) — способ деления клеток, приводящий к уменьшению в них числа хромосом вдвое. Мейоз служит ключевым звеном гаметогенеза у животных и спорогенеза у растений, в результате которого из диплоидных клеток образуются гаплоидные клетки. Мейоз протекает сходно почти у всех организмов. Он состоит из двух последовательных клеточных делений — мейоза I (первое деление) и мейоза II (второе деление), разделенных непродолжительным периодом интеркинеза. При этом репликация ДНК предшествует лишь первому делению. В каждом из делений мейоза различают те же четыре стадии, что и при митозе: профазу, метафазу, анафазу и телофазу, которые, однако, имеют некоторые особенности Первое мейотическое деление (мейоз I) приводит к уменьшению вдвое количества хромосом и называется редукционным. В результате из одной диплоидной клетки (2пАС) образуются две гаплоидные клетки (л2С).
Отличительной особенностью первого деления мейоза является сложная и продолжительная профаза I, в начале которой хромосомы спирализуются и становятся видимыми в световой микроскоп. Затем гомологичные хромосомы сближаются и объединяются друг с другом. Их конъюгация (лат. conjugatio — соединение) происходит сначала в отдельных точках, а затем и по всей длине хромосомы, вследствие чего образуются биваленты. Поскольку каждая из гомологичных хромосом состоит издвуххроматид, бивалент, включающий четыре хроматиды, называют также тетрадой. В диплоидной клетке образуется п бивалентов, и, таким образом, после конъюгации формула клетки приобретает вид n4C. В некоторых местах конъюгированных хромосом хроматиды остаются соединенными, перекрещиваются друг с другом, рвутся и обмениваются своими участками. Процесс обмена участками несестринских хроматид гомологичных хромосом называется кроссинговером (англ. crossingover — перекрест). К концу профазы связь между гомологами ослабевает, и целостность бивалента в это время сохраняется лишь благодаря соединению между собой хроматид в местах кроссинговера, называемых хиазмами. В это время ядрышки и ядерная оболочка распадаются, центриоли клеточного центра расходятся к полюсам клетки и образуется веретено деления. Хромосомы еще больше спирализуются, и биваленты начинают двигаться к плоскости экватора клетки. Обычно профаза занимает около 90% времени, необходимого для завершения мейоза.В метафазе I мейоза завершается формирование веретена деления. Гомологичные хромосомы, объединенные в биваленты, выстраиваются в экваториальной плоскости клетки, образуя метафазную пластинку. При этом центромерные районы каждой хромосомы бивалента, в отличие от митоза, взаимодействуют с нитями веретена деления только от одного полюса. В результате центромерные районы хромосом, составляющих бивалент, оказываются соединенными с разными полюсами.В анафазе I взаимодействие сестринских хроматид прекращается по всей длине хромосомы, за исключением центромерного района. Под действием нитей веретена гомологичные хромосомы бивалентов, каждая из которых состоит из двух сестринских хроматид, отходят к противоположным полюсам клетки. В результате у каждого полюса оказывается по одной гомологичной двухроматидной хромосоме из каждой их пары. Напомним, что такое поведение хромосом при мейозе отличается от митоза, при котором к полюсам клетки расходятся отдельные их хроматиды.В телофазе I хромосомы деспирализуются; формируется ядерная мембрана; разделяется цитоплазма. Клетки, образующиеся в результате первого мейотического деления, содержат гаплоидный набор хромосом и удвоенное количество ДНК и имеют генетическую формулу п2С. После короткой интерфазы, во время которой репликации ДНК не происходит, они приступают к следующему делению.Второе деление мейоза (мейоз II) по своему механизму сходно с митозом и включает такие же стадии: профазу И, метафазу II, анафазу II и телофазу П. В каждой из двух делящихся клеток ядрышки и ядерные мембраны разрушаются, хроматиды укорачиваются и утолщаются. Центриоли расходятся к противоположным полюсам; формируется веретено деления. Двухроматидные хромосомы размещаются таким образом, что их центромеры выстраиваются по экватору веретена. Затем центромеры разделяются, и нити веретена растаскивают сестринские хроматиды (которые после отделения друг от друга называются хромосомами), к противоположным полюсам клетки. Хромосомы деспирализируются и становятся плохо различимыми; вокруг них формируются ядерные оболочки. Разделением цитоплазмы завершается образование двух новых клеток, каждая из которых содержит гаплоидное число однохроматидных (нереплицированных) хромосом и имеет формулу пС. Второе деление мейоза называют также эквационным, или уравнительным. В итоге, в результате двух последовательных мейотических делений из одной клетки с диплоидным набором двухроматидных хромосом (2п4С) образуется четыре клетки с гаплоидным набором однохроматидных хромосом (пС).
Биологическое значение мейоза состоит в сохранении постоянства количества хромосом в ряду поколений организмов, размножающихся половым путем. Если бы в процессе мейоза не наблюдалась редукция набора хромосом, то в каждом следующем поколении при слиянии яйцеклеток и сперматозоидов количество хромосом увеличивалось бы в два раза. Мейоз служит также основой комбинативной изменчивости.
макроспорогаметогенез у цветковых растений
правила проведения гибридологического метода.законы мендаля
Сущность гибридологического метода заключается в следующем: 1) для скрещивания выбирают родительские формы, четко различающиеся по одной, двум или трем парам контрастных, альтернативных признаков. Например, у одного растения окраска семядолей зрелых семян желтая, у другого — зеленая, форма семян — круглая или морщинистая и т. д. Скрещивание, в котором родители отличаются друг от друга одним признаком, в последующем получило название моногибридного, двумя — дигиб-ридного, многими признаками — полигибридного; 2) выбранные для скрещивания родительские формы должны быть генетически чистыми. После двухлетнего предварительного испытания Мендель отобрал 22 сорта гороха, которые за время опытов ежегодно высевали и все без исключения сохраняли свою константность; 3) Мендель ввел точный математический учет наследования каждого отдельного признака. Наблюдению подвергают все без исключения растения в каждом отдельном поколении. Как правило, для определения наследования признака используют гибриды первого, второго и иногда третьего поколений; 4) гибриды и их потомки в каждом из следующих друг за другом поколений не должны обнаруживать заметных нарушений в плодовитости; 5) Мендель ввел буквенное обозначение наследственных задатков (генов) различных признаков. Например, А — ген доминантного признака, а — ген рецессивного признака. При гибридологическом анализе довольно часто используют реципрокное скрещивание. Реципрокным называют два скрещивания, в одном из которых доминантным признаком отличается отцовская форма, в другом — материнская: например, в одном скрещивании отец черной масти, мать красной, в другом, наоборот, мать черной масти, отец красной. Одна из главных причин, обеспечивших успех в работе Менделя, — удачный выбор объекта исследования. Горох — однолетнее растение, имеет много сортов с четко различающимися признаками, легко культивируется, строгий самоопылитель, строение его цветка таково, что почти невозможен занос чужой пыльцы, но при необходимости можно производить искусственное опыление. При анализе закономерностей наследования признаков пользуются некоторыми терминами и понятиями, введенными уже после переоткрытия законов Менделя. Датский ученый В. Ио-ганнсен в 1909 г. ввел понятия «ген», «генотип» и «фенотип». Ген — наследственный задаток. Генотип — совокупность наследственных задатков (генов) организма. Фенотип — совокупность всех признаков и свойств организма, доступных наблюдению и анализу. Английский зоолог У. Бетсон ввел понятия «гомозигота» и «гетерозигота». Гомозиготными называют особей, получивших от отца и матери одинаковые наследственные задатки (гены) по какому-то конкретному признаку. Гетерозиготными называют особей, получивших от отца и матери разные гены. Таким образом, по генотипу особи могут быть гомозиготными (АА или аа) или гетерозиготными (Аа). Фенотип формируется под влиянием генотипа и условий среды. Фенотип можно выразить по определенному признаку такими словами, как рогатый или комолый, высокий или низкий, черный или красный. Наследственные задатки (гены) альтернативных признаков были названы аллелями. Аллели (аллельные гены) расположены в одинаковых точках (локусах) парных гомологичных хромосом.Один аллельный ген (А или а) зигота получает с яйцеклеткой от матери, другой (А или а) — со сперматозоидом от отца. При изучении наследования признаков составляются схемы скрещивания; скрещивание обозначают знаком умножения (х), который ставится между родителями. При написании схем родительские формы обозначают буквой Р (от слова parentes — родители), женский пол обозначают знаком ? (символ планеты Венеры), мужской — (/(символ планеты Марс). Рядом со знаками ? и j проставляют генотип родителя, а ниже записывают типы производимых ими гамет (половых клеток). Затем в результате соединения гамет родителей определяют генотип потомства. Полученное в результате скрещивания потомство называют гибридами и обозначают буквой F (от слова filii — дети), внизу буквы ставят цифру, указывающую, к какому поколению оно относится, например, Fi — гибрид первого поколения, F2 — второго, F3 — третьего поколения и т. д. Мендель вел учет наследования отдельно по каждой паре альтернативных признаков, отвлекаясь от других различий между родительскими формами. На основании опытов Мендель установил три закона: единообразия гибридов первого поколения, расщепления, независимого наследования признаков, а также правило чистоты гамет. Закон единообразия гибридов первого поколения, или первый закон Менделя, утверждает, что потомство первого поколения от скрещивания устойчивых форм, различающихся по одному признаку, имеет одинаковый фенотип по этому признаку. При этом все гибриды могут иметь фенотип одного из родителей (полное доминирование), как это имело место в опытах Менделя, или, как было обнаружено позднее, промежуточный фенотип (неполное доминирование). В дальнейшем выяснилось, что гибриды первого поколения могут проявить признаки обоих родителей (кодоминировапие). Этот закон основан на том, что при скрещивании двух гомозиготных по разным аллелям форм (АА и аа) все их потомки одинаковы по генотипу (гетерозиготны — Аа), а значит, и по фенотипу.
Закон расщепления, или второй закон Менделя, гласит, что при скрещивании гибридов первого поколения между собой среди гибридов второго поколения в определ. соотношениях появляются особи с фенотипами исходных родительских форм и гибридов первого поколения. Так, в случае полного доминирования выявляются 75% особей с доминантным и 25% с рецессивным признаком, т. е. два фенотипа в отношении 3:1 (рис. 1). При неполном доминировании и кодомииировании 50% гибридов второго поколения имеют фенотип гибридов первого поколения и по 25% — фенотипы исходных родительских форм, т. е. наблюдают расщепление 1:2:1. В основе второго закона лежит закономерное поведение пары гомологичных хромосом (с аллелями А и а), к-рое обеспечивает образование у гибридов первого поколения гамет двух типов, в результате чего среди гибридов второго поколения выявляются особи трёх возможных генотипов в соотношении 1АА:2Аа:1аа. Конкретные типы взаимодействия аллелей и дают расшепления по фенотипу в соответствии со вторым законом Менделя.
Закон независимого комбинирования (наследования) признаков, или третий закон Менделя, утверждает, что каждая пара альтернативных признаков ведёт себя в ряду поколений независимо друг от друга, в результате чего среди потомков второго поколения в определ. соотношении появляются особи с новыми (по отношению к родительским) комбинациями признаков. Напр., при скрещивании исходных форм, различающихся по двум признакам, во втором поколении выявляются особи с четырьмя фенотипами в соотношении 9:3:3:1 (случай полного доминирования). При этом два фенотипа имеют «родительские» сочетания признаков, а оставшиеся два — новые. Этот закон основан на независимом поведении (расщеплении) неск. пар гомологичных хромосом (рис. 2). Напр., при дигибридном скрещивании это приводит к образованию у гибридов первого поколения 4 типов гамет (АВ, Ab, aB, ab) и после образования зигот — закономерному расщеплению по генотипу и соответственно по фенотипу.
Как один из М. з. в генетич. лит-ре часто упоминают закон чистоты гамет. Однако, несмотря на фундаментальность этого закона (что подтверждают результаты тетрадного анализа), он не касается наследования признаков и, кроме того, сформулирован не Менделем, а У. Бэтсоном (в 1902).
10) закон единообразия гибридов первого поколения.
Закон единообразия первого поколения гибридов, иди первый закон Менделя. Для иллюстрации первого закона Менделя — закона единообразия первого поколения — воспроизведем его опыты по монтлгибридному скрещиванию растений гороха. Скрещивание двух организмов называется гибридизацией, потомство от скрещивания двух особей с разной наследственностью называют гибридным, а отдельную особь — гибридом.Моногибридным называется скрещивание двух организмов, отличающихся друг от друга по одной паре альтернативных (взаимоисключающих) признаков. Следовательно, при таком скрещивании прослеживаются закономерности наследования только двух признаков, развитие которых обусловлено парой аллельных генов. Все остальные признаки, свойственные данным организмам, во внимание не принимаются.
Если скрестить растения гороха с желтыми и зелеными семенами, то у всех полученных в результате этого скрещивания гибридов семена будут желтыми. Такая же картина наблюдается при скрещивании растений, обладающих гладкой и морщинистой формой семян; все потомство первого поколения будет иметь гладкую форму семян. Следовательно, у гибрида, первого поколения из каждой пары альтернативных признаков развивается только один. Второй признак как бы исчезает, не проявляется. Явление преобладания у гибрида признака одного из родителей Г. Мендель назвал доминированием. Признак, проявляющийся у.гибрида первого поколения и подавляющий развитие другого признака, был назван доминантным, а противоположный, т, е. подавляемый, признак — рецессивным. Если в генотипе организма (зиготы) два одинаковых аллельных гена — оба доминантные или оба рецессивные (АА или аа), такой организм называется гомозиготным. Если же из пары аллельных генов один доминантный, а другой рецессивный (Аа), то такой организм носит названиегетерозиготного.
Закон доминирования — первый закон Менделя — называют также законом единообразия гибридов первого поколения, так как у всех особей первого поколения проявляется один признак.