

II.II. Частная физиология сенсорных систем
1. Зрительная сенсорная система
Зрительная сенсорная система (зрительный анализатор) представляет собой совокупность защитных оптических, рецепторных и нервных структур, воспринимающих и анализирующих световые раздражители. Зрительная система состоит из периферического отдела – глаза, промежуточных звеньев – подкорковых зрительных центров (наружное коленчатое тело таламуса и переднее двухолмие) и конечного звена – зрительной коры. Все уровни зрительной системы соединены друг с другом проводящими путями.
Строение глаза
Орган зрения человека – глаз (рис. 1) имеет шарообразную (или близкую к таковой) форму. Он включает в себя ядро, покрытое тремя оболочками.
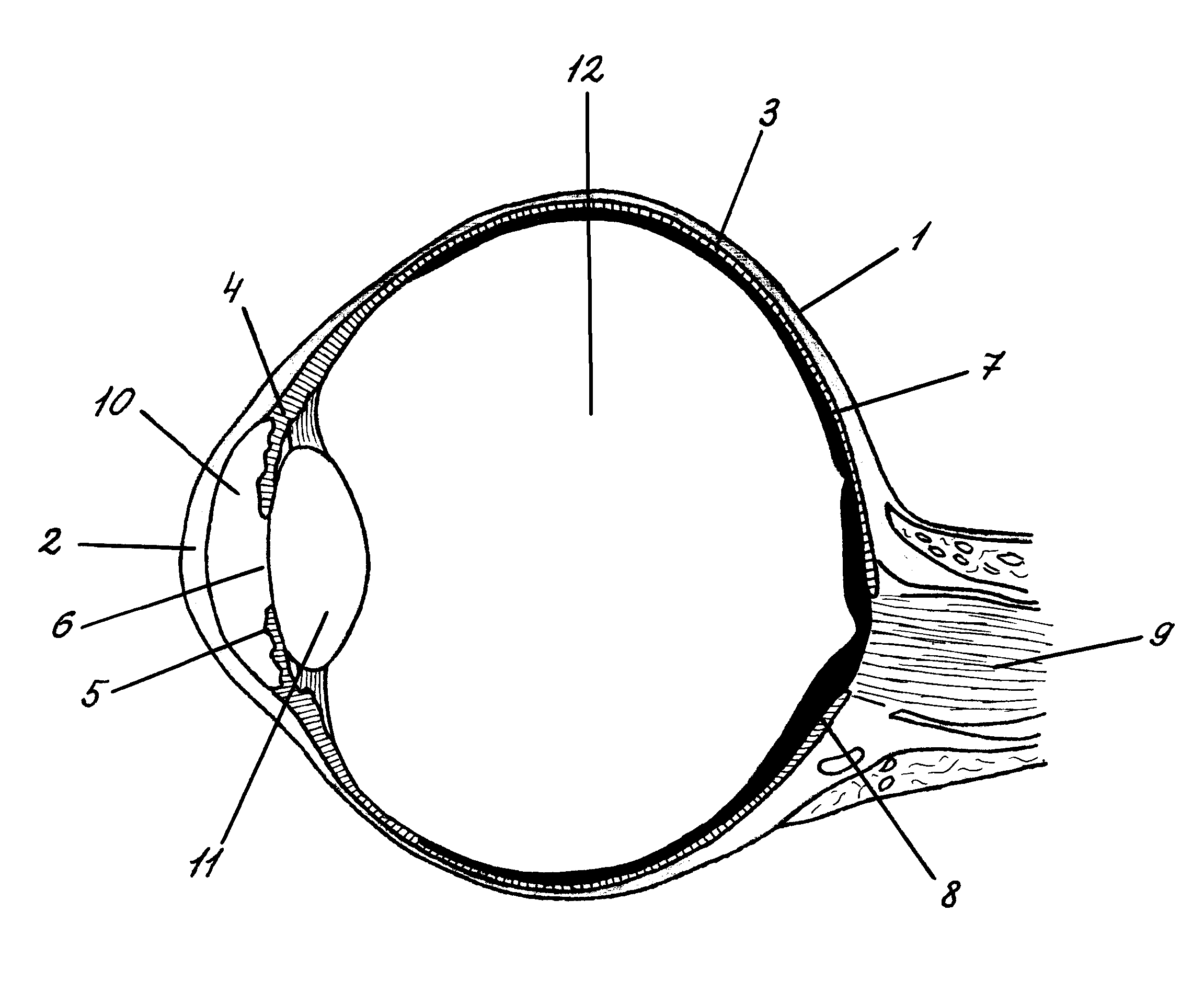
Горизонтальный разрез правого глаза: 1 – склера; 2 – роговая оболочка (роговица); 3 – сосудистая оболочка; 4 – ресничное тело; 5 – радужная оболочка; 6 – зрачок; 7 – пигментный эпителий; 8 – сетчатка; 9 – зрительный нерв; 10 – передняя камера глаза; 11 – хрусталик; 12 – стекловидное тело.
Наружная плотная непрозрачная оболочка – склера - выполняет главным образом защитную, механическую функцию. В передней части глазного яблока склера переходит в прозрачную роговую оболочку, или роговицу. Кривизна поверхности роговицы определяет особенности преломления света. Роговица обладает наибольшей преломляющей способностью. Под склерой лежит сосудистая оболочка, которая образована сетью кровеносных сосудов. Ее основное назначение – питание глазного яблока. Спереди сосудистая оболочка утолщается и переходит сначала в ресничное тело (мышца, изменяющая кривизну хрусталика) и далее – в радужную оболочку, которые состоят из гладких мышечных волокон, кровеносных сосудов и пигментных клеток. Цвет радужной оболочки зависит от пигментации составляющих ее клеток и их распределения. Между роговицей и радужной оболочкой находится передняя камера глаза, наполненная жидкостью – «водянистой влагой». В центре радужной оболочки имеется отверстие – зрачок, играющий роль диафрагмы и регулирующий величину светового потока, проникающего внутрь глаза. Размер зрачка зависит от освещенности. Контроль за изменениями размера зрачка осуществляется автоматически нервными волокнами, заканчивающимися в мускулатуре радужной оболочки. Круговая мышца, суживающая зрачок – сфинктер – иннервируется парасимпатическими волокнами, мышца, расширяющая зрачок – дилататор – иннервируется симпатическими волокнами. Реакция расширения зрачка до максимального диаметра – 7,5 мм – очень медленная: она длится около 5 минут. Максимальное сокращение диаметра зрачка до 1,8 мм достигается быстрее – всего за 5 секунд.
Позади радужной оболочки расположен хрусталик. Он представляет собой двояковыпуклую линзу, расположенную в сумке, волокна которой соединены с ресничными мышцами. С помощью этих мышц хрусталик способен изменять свою кривизну. Такая способность хрусталика называется аккомодацией. Аккомодации обеспечивает ясное видение различно удаленных предметов. При рассматривании близко расположенных предметов кривизна хрусталика увеличивается, если же предмет находится далеко, кривизна уменьшается. Аккомодация хрусталика иногда оказывается недостаточной, чтобы спроецировать изображение точно на сетчатку. Если расстояние между хрусталиком и сетчаткой больше, чем фокусное расстояние хрусталика, то возникает близорукость (миопия). Если сетчатка расположена слишком близко к хрусталику и фокусировка хороша только при рассматривании далеко расположенных предметов, возникает дальнозоркость (гиперметропия).
Внутри глаза, позади хрусталика, находится стекловидное тело. Оно представляет собой коллоидный раствор гиалуроновой кислоты во внеклеточной жидкости. Поскольку и хрусталик, и стекловидное тело являются белковыми структурами, то обменные процессы в них могут нарушаться. Например, с возрастом снижается эластичность хрусталика, поэтому ухудшается способность видения близко расположенных предметов (старческая дальнозоркость), постепенно он теряет свою прозрачность, возникает заболевание, получившее название катаракты. В стекловидном теле могут появляться плотные вкрапления, что субъективно ощущается как темные точки, пылинки в поле зрения. Эти изменения в конечном итоге снижают четкость изображения и могут привести к потере зрения. Стекловидное тело и хрусталик называют оптической системой глаза, которая обеспечивает фокусировку изображения на рецепторной поверхности сетчатки. Изображение на сетчатке оказывается четким, но уменьшенным и перевернутым. Мозг исправляет эту «ошибку», руководствуясь не только поступающей зрительной информацией, но и информацией от других сенсорных систем (вестибулярной, проприоцептивной, кожной).
Строение сетчатки
Сетчатка – с нейроанатомической точки зрения – высокоорганизованная слоистая структура, объединяющая рецепторы и нейроны. Она состоит из нескольких слоев клеток, выполняющих разные функции. Несколько упрощенно строение светочувствительного и проводящего аппарата сетчатки можно представить в виде следующей схемы (рис. 2).
Наружный слой сетчатки, плотно примыкающий непосредственно к сосудистой оболочке, образован пигментными клетками, содержащими пигмент фусцин. Этот пигмент поглощает свет, препятствуя его отражению и рассеиванию, что способствует четкости зрительного восприятия. К пигментному слою изнутри примыкает слой фоторецепторов – колбочек и палочек, которые повернуты от пучка падающего света таким образом, что их светочувствительные сегменты спрятаны в промежутках между клетками пигментного слоя. Каждый фоторецептор состоит из чувствительного к действию света наружного сегмента, содержащего зрительный пигмент, и внутреннего сегмента, содержащего ядро и митохондрии, обеспечивающие энергетические процессы в фоторецепторной клетке.
|
|
Рис. 2. Строение сетчатки: 1 – палочка; 2 – колбочка; 3 – карликовый биполяр; 4 – палочковый биполяр; 5 – плоский биполяр; 6 – горизонтальная клетка; 7 – амакриновая клетка; 8 – ганглиозная клетка; 9 – пигментный эпителий; 10 – мюллерова клетка (глия).
|
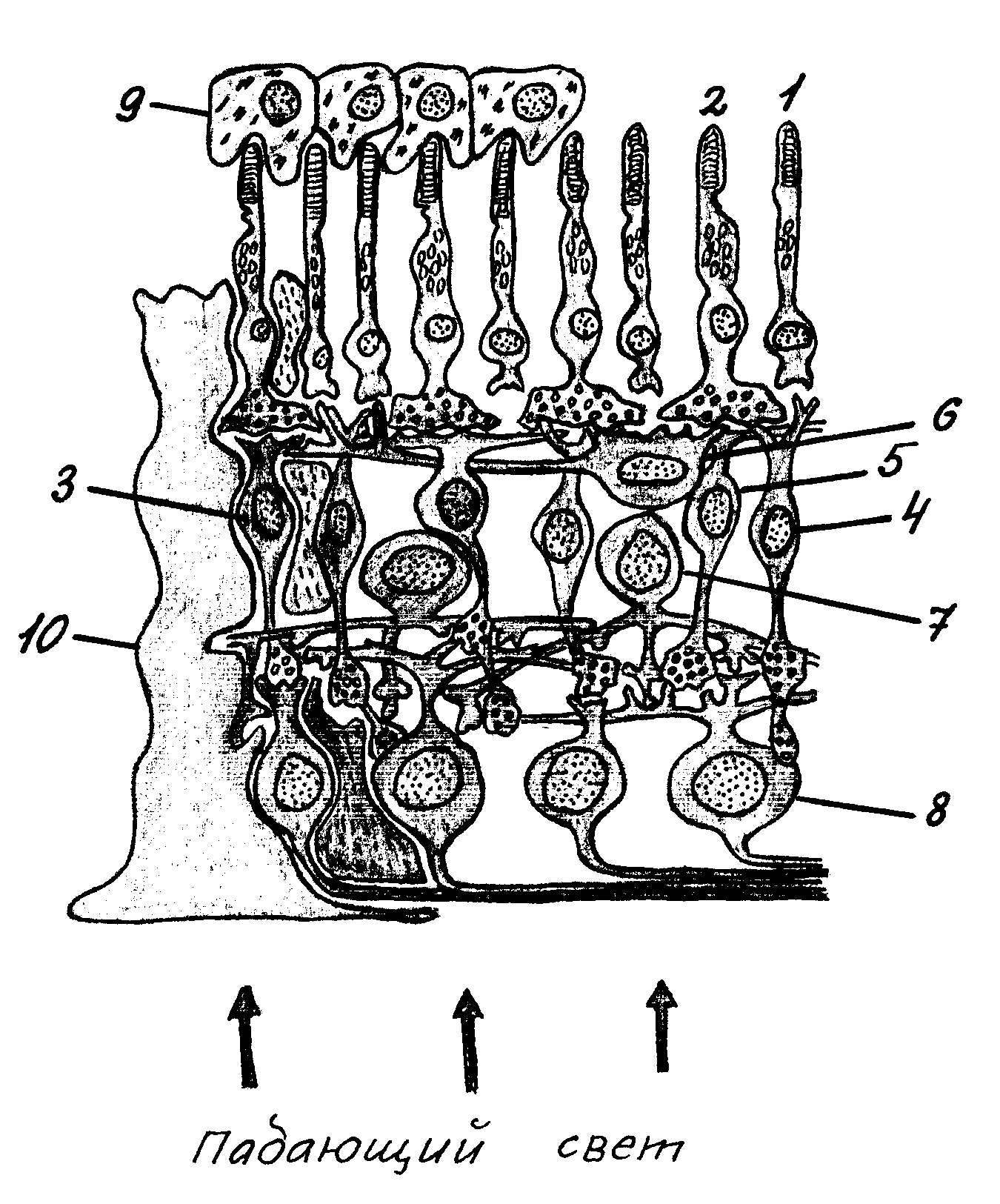
Палочки и колбочки отличаются функционально: палочки реагируют на свет и обеспечивают зрительное восприятие при слабой освещенности, а колбочки функционируют при ярком свете и обеспечивают восприятие цвета. Фоторецепторы содержат зрительные пигменты, которые по своей природе являются белками. В палочках содержится пигмент родопсин, в колбочках – пигменты иодопсин, хлоролаб и эритлаб, необходимые для цветового зрения. Свет, попадая на сетчатку, вызывает разложение пигмента. Эти химические преобразования сопровождаются изменением потенциала на мембране рецептора, т.е. возникновением рецепторного потенциала. Таким образом, функция рецепторов сводится к преобразованию энергии квантов света в электрическую энергию ответа клетки.
На сетчатке каждого глаза около 6 млн. колбочек и 120 млн. палочек – всего около 130 млн. фоторецепторов. Они распределены по сетчатке неравномерно: чем ближе к периферии, тем больше палочек, чем ближе к центру, тем больше колбочек, наконец, в самом центре сетчатки напротив зрачка располагаются только колбочки. Эта область называется желтым пятном или центральной ямкой. Здесь плотность колбочек составляет 150 тысяч на 1 квадратный миллиметр, поэтому в области желтого пятна острота зрения максимальна.
Центральная часть сетчатки представлена биполярными клетками, имеющими по два относительно длинных отростка, одним из которых они контактируют с фоторецепторами, другим – с ганглиозными клетками сетчатки, которые, в свою очередь, составляют ее внутреннюю часть. Ганглиозные клетки обладают круглыми рецептивными полями с четко выраженными центром и периферией. Размеры центральной части и периферической каймы могут изменяться в зависимости от освещенности. Если центр возбуждается при попадании света на сетчатку, то периферия при этом тормозится. Может быть и обратное соотношение. Ганглиозные клетки имеют как палочковые, так и колбочковые рецептивные поля. В последнем случае центр и периферия рецептивного поля возбуждается (или тормозится) определенным цветом. Например, если в ответ на предъявление красного цвета центр возбуждается, то периферия будет тормозиться. Такие комбинации могут быть самыми разнообразными. Ганглиозные клетки в отличие от других элементов сетчатки способны генерировать потенциалы действия, направляющиеся по нервным волокнам к центральным структурам мозга.
Ганглиозные клетки являются выходными элементами сетчатки. Их аксоны формируют зрительный нерв, который пронизывает сетчатку в противоположном направлении и входит в полость черепа. В месте вхождения в сетчатку волокон зрительного нерва фоторецепторы отсутствуют; эта область получила название слепого пятна.
Таким образом, фоторецепторы, биполяры и ганглиозные клетки представляют собой три последовательных звена переработки зрительной информации.
На уровне между рецепторами и биполярами имеются специализированные клетки с горизонтальным расположением отростков, которые регулируют передачу возбуждения от рецепторов к биполярам и носят название горизонтальных. Между биполярами и ганглиозными клетками, располагаясь как бы симметрично горизонтальным, находятся амакриновые клетки, которые «управляют» передачей электрических сигналов от биполяров к ганглиозным клеткам. На теле амакриновых клеток заканчиваются центробежные волокна, несущие возбуждение из ЦНС. Горизонтальные и амакриновые клетки обеспечивают латеральное торможение между соседними клеточными элементами сетчатки, ограничивая распространение зрительного возбуждения внутри нее.
В заключение следует отметить, что сетчатка как система позволяет выделять такие характеристики светового сигнала, как его интенсивность (яркость), пространственные параметры (размер, конфигурация). Рецептивные поля, построенные по принципу антагонистических отношений центра и периферии, позволяют оценивать контрастность и контуры изображения, а также оптимальным образом выделять полезный сигнал из шума.
Центральные структуры зрительной системы
Наружное коленчатое тело (НКТ) – основной подкорковый центр зрительного анализатора. Большая часть зрительных волокон (аксонов ганглиозных клеток) в составе зрительного тракта оканчивается в этой структуре. Основные пути от НКТ идут в 17-е, в меньшей степени - в 18-е и 19-е зрительные поля (по Бродману). Другие волокна направляются к верхнему двухолмию, подушке зрительного бугра и другим структурам.
Рецептивные поля нейронов НКТ имеют различную форму – от круглой до вытянутой; существуют поля с возбуждающимся центром и тормозной периферией и наоборот. В НКТ кодируется информация о пространственных характеристиках (размере) зрительного изображения, об уровне освещенности, о цвете. Ввиду многочисленных связей НКТ с различными таламическими ядрами (в первую очередь ассоциативными) можно предположить, что на этом уровне происходит перераспределение потока информации по различным каналам и начинается процесс анализа наиболее сложных параметров стимула, в частности анализа информации о биологической значимости данного раздражителя.
Переднее двухолмие. Хотя к передним холмам среднего мозга направляется не более 10% зрительных волокон, эта структура играет важную роль в организации ориентировочного поведения.
Переднее двухолмие имеет слоистую структуру. В верхних слоях заканчиваются волокна, идущие от сетчатки, коры мозга (затылочной, лобной и височной областей), из спинного мозга, от задних холмов четверохолмия, НКТ, мозжечка и черной субстанции. Нижние слои называют эфферентным центром, дающим начало наиболее длинным нисходящим путям. Они направляются в спинной мозг, к ядрам черепно-мозговых нервов, в ретикулярную формацию и другим структурам, обеспечивающим зрительные ориентировочные рефлексы.
Большая часть нейронов не отвечает или слабо отвечает на действие диффузного света или на неподвижные объекты, но дает сильную реакцию на движение, поэтому их называют детекторами движения. При этом более 75% нейронов реагируют только на определенное направление движения (преимущественно на движение в горизонтальной плоскости), и сила реакции зависит от скорости движения. Удаление или разрушение переднего двухолмия у животных сопровождается потерей способности следить за движущимся объектом. В связи с этим считают, что переднее двухолмие осуществляет координацию движений глазных яблок с поступлением зрительной информации.
Зрительная кора. Зрительная кора имеет слоистую структуру. В зависимости от выраженности тех или иных слоев ней выделяют первичную область – 17-е поле, вторичную – 18-е поле и третичную – 19-е поле по Бродману. Поле 17 является центральным полем коркового ядра анализатора, 18-е и 19-е поля – периферическими.
Функциональное значение зрительной коры чрезвычайно велико. Это доказывается наличием многочисленных связей не только со специфическими зрительными подкорковыми образованиями, но и с ассоциативными и неспецифическими ядрами таламуса, с ретикулярной формацией, теменной ассоциативной областью и т.д.
Реакции одиночных нейронов зрительной коры впервые были зарегистрированы Р. Юнгом в начале 50-х г.г. Было показано, что на диффузный засвет сетчатки реагирует лишь около половины нейронов. Большая же часть нейронов отвечает лишь на стимулы, ориентированные определенным образом (лучше всего – на светлые полосы на темном фоне или пространственные решетки, состоящие из чередующихся светлых и темных полос).
В 60-х гг. ХХ в. американские нейрофизиологи Д. Хьюбел и Т. Визель, исследуя свойства нейронов зрительной коры, выделили три типа рецептивных полей – простые, сложные и сверхсложные. Рецептивные поля простого типа имеют прямоугольную форму, состоят из центра и периферии, границы которых приблизительно параллельны друг другу. Лучше всего они отвечают на движение светлой полосы по темному фону или наоборот. Как правило, у нейронов с простым типом рецептивного поля существует предпочитаемое направление движения, реакция на которое выражена сильнее всего.
Нейроны с рецептивным полем сложного типа лучше отвечают на полоску или решетку, оптимальным образом ориентированную относительно сетчатки (вертикальное, горизонтальное или наклонное положение).
Нейроны сверхсложного типа могут отвечать на несколько положений полосы (линии), ее повороты на определенный угол, на угол, образуемый двумя линиями, на кривизну контура или более сложные пространственные характеристики зрительного изображения. Предполагается, что существует конвергенция нейронов с простыми рецептивными полями на нейроны более высокого порядка. В 17-м поле коры встречается больше нейронов с простыми, а в 18-м и 19-м – со сложными и сверхсложными рецептивными полями.
На основании этого Д. Хьюбел и Т. Визель сформулировали детекторную теорию переработки зрительной информации. Суть ее состоит в том, что нейроны с простыми рецептивными полями, являясь детекторами элементарных признаков зрительного изображения (например, ориентации линий), конвергируют с нейронами более высокого уровня, которые в результате этой конвергенции приобретают более сложные свойства. Таким образом, существует иерархия нейронов-детекторов, на верхних ступенях которой находятся детекторы наиболее сложных признаков зрительного изображения. Однако, как было показано в дальнейшем, такого рода нейроны, ответственные за опознание целостных зрительных образов, расположены за пределами собственно зрительной коры – в первую очередь, в нижневисочной области. Таким образом, процесс зрительного восприятия не заканчивается в проекционных областях, а продолжается на более сложных уровнях ассоциативных корковых зон.
Альтернативой детекторной теории является пространственно-частотная гипотеза переработки зрительной информации, предложенная английским исследователем Ф. Кемпбеллом и отечественным физиологом В.Д. Глезером. Согласно этой гипотезе, нейроны зрительной коры определяют две основные характеристики зрительного изображения – ориентацию стимула (полосы, решетки) и его пространственную частоту. При этом нейроны разных участков коры «настроены» на стимулы разной пространственной ориентации и пространственной частоты. Таким образом, в 17-м поле зрительной коры создается «мозаика» из возбужденных и невозбужденных нейронов, изоморфно отображающая пространственное распределение возбужденных и заторможенных рецепторов сетчатки глаза. Нейроны вторичной и третичной зрительных областей (18-е и 19-е поля) используют информацию, поставляемую из первичной коры (17-е поле), для формирования более крупных подобразов зрительного изображения.
Таким образом, на уровне зрительной коры осуществляется тонкий, дифференцированный анализ наиболее сложных признаков зрительного сигнала (выделение контуров, очертаний, формы объекта, локализации, перемещений в пространстве и т.д.). На уровне вторичной и третичной областей, по-видимому, осуществляется наиболее сложный интегративный процесс, подготавливающий организм к опознанию зрительных образов и формированию целостной сенсорно-перцептивной картины мира. Формирование же целостных зрительных образов, их опознание и оценка биологической значимости осуществляется в ассоциативных областях в первую очередь, заднетеменной и нижневисочной.
Ассоциативные зоны коры. В нейрофизиологических исследованиях было показано, что нейроны нижневисочной коры (НВК) лучше всего отвечают на целостные образы (например, на геометрические фигуры). При этом можно выделить клетки, отвечающие только на одну фигуру (например, круг), или реагирующие на несколько различных изображений (круг, треугольник, крест и квадрат). Ответы нейронов, как правило, инвариантны к преобразованиям фигур, т.е. не зависят от размера, поворота, цвета изображений, освещенности и т.д.
В целом считают, что нейроны НВК отвечают на сенсорное значение зрительного стимула независимо от его значимости для моторного поведения. При этом для НВК важны не отдельные характеристики стимула, а их определенные сочетания. Очевидно, НВК осуществляет классификацию изображений в соответствии с конкретной задачей, стоящей перед животными и человеком. При повреждении этой области у человека нарушаются процессы опознания объектов и памяти.
Заднетеменная кора (ЗТК) создает нейронную конструкцию (модель) окружающего пространства, описывая расположение и перемещение объектов в этом пространстве по отношению к телу, а также положение и движение тела по отношению к окружающему пространству. Другими словами, в ЗТК происходит переработка информации, описывающей соотношения между внутренними и наружными координатными системами. Имеются также данные о связи нейронов ЗТК с произвольным вниманием к тому или иному зрительно воспринимаемому объекту.
При двусторонних повреждениях теменной области у человека возникают нарушения зрительного восприятия пространства. Такие больные не могут оценить пространственные преобразования фигур, у них нарушена топографическая ориентировка и т. д. Это свидетельствует о важной роли ЗТК в восприятии пространства и пространственных соотношений между объектами, находящимися в поле зрения.
Опознание образов осуществляется содружественной работой НВК и ЗТК. Если первая осуществляет опознание отдельных элементов (фрагментов) целостной зрительной ситуации, инвариантно к их пространственным преобразованиям, то вторая формирует целостную картину окружающего мира.
Лобная кора благодаря своим многочисленным связям со структурами памяти и структурами лимбической системы осуществляет оценку значимости стимула для организма и планирование соответствующего поведенческого акта.
Световая чувствительность и адаптация
Световая чувствительность характеризует способность зрительной системы воспринимать излучение света. Наибольшую световую чувствительность глаз имеет в темноте. Минимальное количество световой энергии, необходимое в этих условиях для возникновения ощущения света, называют абсолютным порогом. Фоторецептор способен возбуждаться при действии одного-двух квантов света, однако, для возникновения светового ощущения необходима суммация возбуждения от нескольких рецепторов. В естественных условиях зрительная система крайне редко работает на пределе, т.е. в околопороговой области, и основное значение для зрения имеет контрастная чувствительность, т.е. чувствительность в условиях световой адаптации. Если тестовое пятно находится на освещенном фоне, то минимальную разность яркости пятна Вc и фона Вf, которая воспринимается наблюдателем как едва заметное различие, называют разностным, или дифференциальным, порогом (∆ В): ∆В = │Вc – Вf│. Отношение разностного порога к освещенности фона называют пороговым контрастом или относительным дифференциальным порогом. Величина относительного дифференциального порога безразмерная и показывает, насколько нужно изменить величину тестового стимула по отношению к фону, чтобы уловить едва заметную разницу между ними. Например, если относительный дифференциальный порог равен 0,03, то это означает, что тестовый стимул должен отличаться от фона на 3%. Согласно закону Бугера-Вебера, ∆В/Вf = const, или ∆В = k∙Вf (разностный порог растет пропорционально освещенности). Однако этот закон справедлив лишь для среднего диапазона интенсивностей и нарушается при малых и больших значениях стимула.
Большое значение для световой чувствительности зрительной системы имеет ее способность к адаптации, т.е. к функциональной перестройке, позволяющей работать в оптимальном режиме при данном уровне освещенности. Различают темновую и световую адаптацию. Темновая адаптация характеризуется максимальным повышением световой чувствительности (снижением абсолютных порогов) зрительной системы для восприятия светового раздражителя в абсолютной темноте. Световая адаптация характеризует чувствительность системы при разных уровнях освещенности.
Темновая адаптации включает в себя изменение палочковой и колбочковой чувствительности. Палочковая адаптация завершается через 7–8 минут, изменения палочковой чувствительности происходят примерно в течение 30 мин. Механизм темновой адаптации с одной стороны состоит в постепенном восстановлении зрительного пигмента в темноте, с другой – в перестройка рецептивных полей в передающей системе рецептор – биполяр – ганглиозная клетка. Так, обнаружено, что в процессе темновой адаптации уменьшается вплоть до полного исчезновения тормозная «кайма» на периферии рецептивного поля ганглиозной клетки, а следовательно, увеличивается ее световая чувствительность.
Световая чувствительность при световой адаптации понижается при переходе от меньшей освещенности к большей. Она протекает быстрее, чем темновая адаптация, и составляет примерно 1–3 минуты.
Острота зрения
Острота зрения характеризует предельную пространственную разрешающую способность зрительной системы, т.е. способность глаза различать две близко расположенные точки как раздельные. Острота зрения определяется как оптикой глаза, так и его нейронными механизмами.
При измерении остроты зрения чаще всего пользуются методом обнаружения, когда предъявляют светлый тест-объект на темном фоне или темный на светлом. Так, наблюдатель должен опознать буквы разного углового размера, определить наклон решетки, состоящей из параллельных полос, и т.д. Наибольшее распространение получили кольца Ландольта, в которых требуется определить положение разрыва на кольце. За количественную меру остроты зрения принимается величина, обратная углу зрения, минимального по размеру, но еще воспринимаемого объекта.
Острота зрения зависит от ряда факторов: освещенности, контрастности фона и текста, состояния и адаптации рецепторного аппарата, оптического аппарата глаза. Она обусловлена также перестройкой рецептивных полей ганглиозных клеток сетчатки. При повышении уровня освещенности размер центра рецептивного поля уменьшается, а влияние тормозной периферии нарастает. Можно предположить, что когда изображение двух точек попадает в два смежных рецептивных поля, разделенных тормозной периферией, вероятность их различения выше, чем в том случае, когда такая периферия отсутствует.
Острота зрения является также функцией положения тест-объекта на сетчатке (или удаления последнего от центральной ямки). Чем дальше от центра производится измерение, тем острота зрения меньше.
Движения глаз и их роль в зрении
Движения глаз играют весьма важную роль в зрительном восприятии. Даже в том случае, когда наблюдатель фиксирует взглядом неподвижную точку, глаз не находится в покое, а все время совершает небольшие движения, которые являются непроизвольными. Движения глаз выполняют функцию дезадаптации при рассматривании неподвижных объектов. Другая функция мелких движений глаза – удерживание изображения в зоне ясного видения.
В реальных условиях работы зрительной системы глаза все время перемещаются, обследуя наиболее информативные участки поля зрения. При этом одни движения глаз позволяют рассматривать предметы, расположенные на одном удалении от наблюдателя, например, при чтении или рассматривании картины, другие – при рассматривании объектов, находящихся на разном удалении от него. Первый тип движений – это однонаправленные движения обоих глаз, в то время как второй осуществляет сведение или разведение зрительных осей, т.е. движения направлены в противоположные стороны.
Показано, что перевод глаз с одних объектов на другие определяется их информативностью. Взор не задерживается на тех участках, которые содержат мало информации, и в то же время длительно фиксирует наиболее информативные участки (например, контуры объекта). Эта функция нарушается при поражении лобных долей. Движение глаз обеспечивает восприятие отдельных признаков предметов, их соотношение, на основе чего формируется целостный образ, хранящийся в долговременной памяти.
Цветовое зрение
Восприятие цвета обусловлено работой двух механизмов. Первичным является фоторецепторный механизм, основанный на существовании рецепторов, избирательно реагирующих на разные участки спектра. В сетчатке были обнаружены три типа колбочек с максимумами поглощения в различных областях спектра (синий, зеленый и красный).
В то же время в психологии и физиологии описан ряд фактов, которые невозможно объяснить, основываясь на фоторецепторном механизме. Такими примерами могут служить феномены одновременного и последовательного контраста. Одновременный контраст заключается в изменении цветового тона в зависимости от фона, на котором предъявляется тот или иной тестовый стимул. Например, серое пятно на красном фоне приобретает зеленоватый оттенок, на желтом – синеватый и т.д. Феномен последовательного контраста состоит в том, что если достаточно долго смотреть на окрашенную в определенный цвет поверхность (например, красную), а затем перевести взор на белую, то она приобретает оттенок оппонентного цвета (в данном случае – зеленоватый). Здесь вступает в работу центральный механизм. Суть его заключается в том, что нейроны ганглиозных клеток, НКТ и зрительной коры обладают цветооппонентными рецептивными полями, т.е. центр рецептивного поля активируется одним цветом, а его периферия – другим, противоположным (оппонентным). Это обусловлено особенностями их рецептивных полей, включающих возбуждающие и тормозные связи с разными типами колбочек. Описаны две цветооппонентные системы: красно-зеленая, желто-синяя.
Таким образом, восприятие цвета обусловлено работой двух разных механизмов, работающих на разных уровнях зрительной системы.
Стереоскопическое зрение
Стереоскопическое зрение позволяет оценить глубину пространства, т.е. относительную удаленность объектов в поле зрения. Оно обусловлено неодинаковым изображением одного и того же объекта на сетчатках обоих глаз. Поскольку глаза расположены на определенном расстоянии друг от друга, то они воспринимают объект под разным углом (так называемый бинокулярный параллакс), поэтому изображения на правой и левой сетчатке будут несколько отличаться друг от друга. В этом легко убедиться, по очереди закрывая то один глаз, то другой. Глазные оси были бы строго параллельны друг другу только в том случае, если бы фиксируемый объект находился на бесконечном удалении от наблюдателя. По мере приближения объекта он будет восприниматься как объемный, а глазные оси будут сходиться. Наконец, на очень близком расстоянии возникает двоение изображения. Иначе говоря, существует определенная зона видения, в пределах которой объект кажется объемным. Она выражается в угловых минутах. Ее нижняя граница составляет около 2 угл. мин. Это угол зрения, при котором две точки для наблюдателя сливаются в одну, т.е. явление глубины (или стереопсиса) исчезает. На практике эту границу достаточно легко определить: это расстояние, при котором глаза другого человека воспринимаются как одно изображение, что в среднем составляет около 6 км. Верхней границей стереопсиса является угол зрения, составляющий около 10 угл. град., за пределами этой границы изображение начинает двоиться.
Нейрофизиологические механизмы стереоскопического зрения на сегодняшний день изучены не до конца. Показано, однако, что основную роль в возникновении стереопсиса играет характер передачи изображения от сетчатки в высшие центры зрительной системы (рис 3.).
|
|
Рис. 3 Поступление зрительной информации в большие полушария мозга у животных с бинокулярным зрением: 1 – поле зрения; 2 – глазное яблоко; 3 – хиазма; 4 – НКТ; 5 – кора.
|
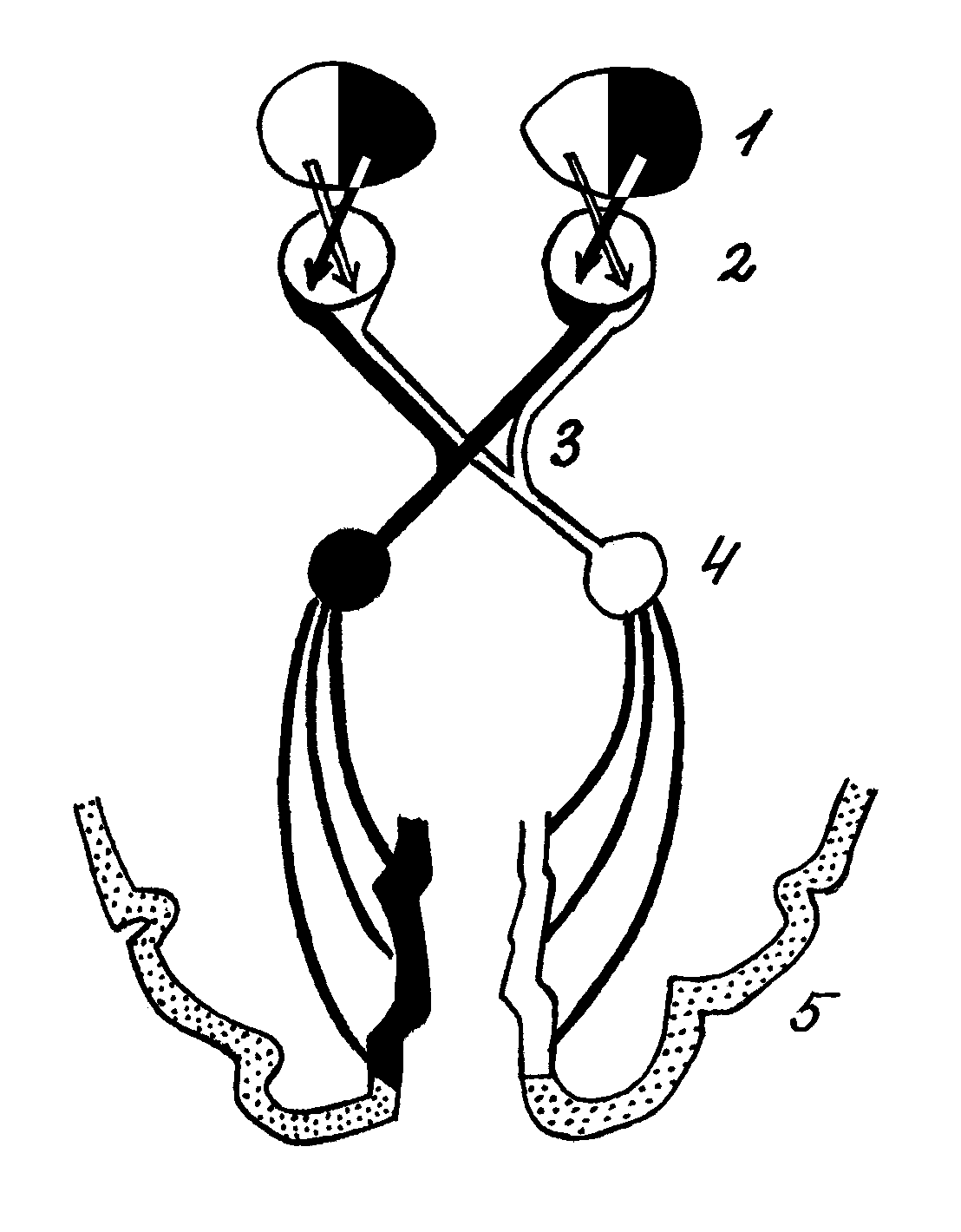
Как известно, у человека в области хиазмы осуществляется неполный перекрест волокон зрительного нерва – волокна от внутренних половин сетчаток перекрещиваются и идут в НКТ и зрительную кору противоположного полушария. Волокна от наружных половин сетчаток идут без перекреста. Таким образом, в каждое полушарие приходит информация от противоположной половины поля зрения. Это и является физиологической основой стереоскопического зрения.
Вопросы и задания для самоконтроля
1. Какие структуры глаза относятся к его оптической системе, какова их роль в зрительном восприятии?
2, Рассмотрите строение сетчатки. Какие из элементов сетчатки способны генерировать потенциал действия?
3. Назовите функциональные отличия фоторецепторов.
4. Какую роль выполняют горизонтaльные и амакриновые клетки?
5. Что является причиной возникновения рецепторного потенциала в фоторецепторах?
6. Kaкaя информация кодируется нейронами наружного коленчатого тела?
7. Какова функция передних холмов в переработке зрительной информации?
8. Чем детекторная теория зрительного восприятия отличается от пространственно-частотной теории?
9. Какую функцию выполняют нижневисочнaя и заднетеменнaя кора?
10. Как изменяется соотношение центра и периферии рецептивного поля ганглиозной клетки при темновой и световой адаптации?
11. От каких факторов зависит острота зрения?
12. Какую роль в зрительном восприятии играют движения глаз?
13. Oпишите основные механизмы цветового зрения.
14. Что лежит в основе стереопсиса?