

gistologia
.pdfТаким образом, из приведенного материала следует, что у хор довых имеется определенная зависимость между количеством желтка в яйцеклетках и характером дробления. Оно изменяется от полного (голобластического) к частичному (меробластическому), а бластула — от целобластулы к дискобластуле.
Общими свойствами развивающихся зародышей всех классов животных на стадии дробления являются постепенное увеличение числа клеток, а следовательно, и ДНК, так как дочерние клетки всегда диплоидные; увеличение площади клеточных поверхностей; возрастание региональных различий клеточных популяций.
ГАСТРУЛЯЦИЯ. ЗАКЛАДКА ОСЕВЫХ ОРГАНОВ
При дальнейшем развитии бластулы в процессе деления, роста, дифференцировки клеток и их перемещений, перегруппировок, взаимодействий и влияний клеток друг на друга формируется сна чала двух-, а затем трехслойный зародыш. Его слоями являются эктодерма (ektos — наружний) — наружный зародышевый лис ток, эндодерма (entos — внутренний) — внутренний зародышевый листок, мезодерма (mesos — средний) — средний зародышевый листок. Из этих листков образуются осевые органы: зачаток нерв ной системы (нервная трубка), хордомезодермальный зачаток и кишечная трубка.
Следует иметь в виду, что закладка этих эмбриональных за чатков начинается задолго до гаструляции. Они входят в состав сформированной бластулы и называются презумптивными (ргаеsumptio — предположение, вероятность) зачатками. Так крыша бластулы построена из презумптивных зачатков эктодермы и нервной трубки, дно бластулы — из презумптивного зачатка ки шечной трубки, а краевая зона — из хордальной пластинки и ме зодермы.
Способы перестройки бластулы в гаструлу сложны и различ ны. Они обусловлены типом дробления и строением бластулы. Для простоты понимания разные способы этого процесса можно све сти к четырем основным типам.
1. Инвагинация, или впячиваные. Часть клеток прогибается в бластоцель, образуя при этом второй — внутренний зародыше вый листок, или энтодерму. Данный способ возможен, если блас тоцель обширен и зародыш напоминает форму шара.
2. Эпиболия, или обрастание. Мелкие, интенсивно делящиеся клетки обрастают зону более крупных бластомеров, обладающих низкой митотической активностью. В этом случае бластоцель всегда незначителен, сдвинут к анимальному полюсу, бластодер ма многослойна.
3.Иммиграция, или вселение. Отдельные клетки или их попу ляции перемещаются из бластодермы и образуют внутренний за родышевый листок — энтодерму.
4.Деляминация, или расслоение. Клетки бластодермы делятся, образуется внутренний клеточный пласт — энтодерма и наруж-
62
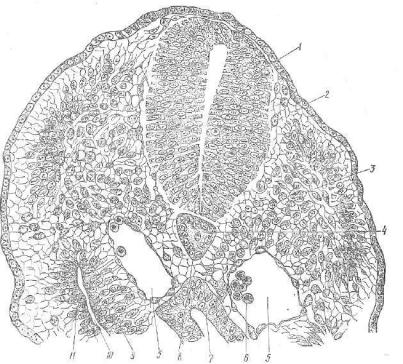
Рис. 46. Способы гаструляции:
1 ~ инвагинация; 2 — эпиболия; з — им миграция; 4 — деляминация.
ный — эктодерма. Если дно бла стулы состоит из желтка, а бласто цель имеет вид узкой щели и сдви нут к зародышевому диску, гаструляция протекает путем делямипации (рис. 46).
У позвоночных гаструляция осуществляется при сочетании не скольких типов, которые могут сменять друг друга или протекать одновременно. Обычно один или два из них являются основными, другие — дополнительными.
А. При изолецитальной, олиголецитальной яйцеклетке, полном, равномерном, синхронном дробле нии и однослойной бластуле, у ко торой бластоцель занимает цент ральную часть, гаструляция проте кает путем инвагинации. В бласто цель начинают впячиваться интен
сивно делящиеся клетки краевой зоны, увлекая за собой крупные бластомеры дна бластулы, содержавшие весь желток развивающе гося организма. Инвагинирующий материал постепенно замещает бластоцель, образуя стенку новой внутренней полости первичной кишки, или гастроцеля.
Гастроцель сообщается с внешней средой с помощью первич ного рта, или бластопора (poms — отверстие). Края бластопора— это его губы. Их четыре: спинная, или дорсальная, губа (dorsum— спина), брюшная, или вентральная (ventres — брюхо), и две боко вые, или латеральные (lateralis — бок) (рис. 47). Через губы бластопора инвагинирует материал разных эмбриональных зачат ков. Так через дорсальную губу перемещается материал будущей хорды, через боковые губы — будущие мезодермальные зачатки, а через вентральную губу — материал кишечной трубки.
Гаструляция путем инвагинации протекает так быстро, что названные презумптивные органы входят в состав энтодермы, ко торая именуется поэтому первичной энтодермой. В состав первич ной эктодермы включены материал нервной трубки и кожная эктодерма.
Таким образом, после инвагинации дна, краевой зоны и вытес нения бластоцеля бластулы зародыш приобретает форму двух слойной чаши, состоящей из наружного листка — первичной экто дермы и внутреннего листка — первичной энтодермы.
63

1 — материал эктодермы; г — бластоцель; з — материал энтодермы: 4 — матери ал мезодермы; 5 — хорда; 6 — гастроцель; 7 — бластопор; 8 — дорсальная губаJ «бластопора.
Зародыш продолжает расти в длину. На его спинной поверх ности в эктодерме от дорсальной губы бластопора обособляется клеточный тяж интенсивно делящихся клеток столбчатой формы.
"Сначала этот тяж имеет форму пластинки, называемой нерв н о й п л а с т и н к о й (рис. 48).
Клетки эктодермы, лежащие вблизи, интенсивно делятся и с •боков наползают на нервную пластинку. Последняя оказывается покрытой эктодермой. Первичная эктодерма после выделения из •ее состава материала нервной пластинки именуется в т о р и ч н о й к о ж н о й э к т о д е р м о й .
Клетки нервной пластинки имеют столбчатую форму и интен- •сивно делятся, в связи с чем вскоре нервная пластинка прогибает ся и образуется желобок, или нервная бороздка. Края нервной пластинки смыкаются и формируется нервная трубка с централь но расположенным каналом. Стадию дифференцировки нервной трубки принято называть н е й р у л я ц и е й.
^Одновременно с закладкой нервной трубки происходит даль нейшее развитие первичной энтодермы. В ее спинной части под нервной трубкой обособляется клеточный тяж — хордальная пла стинка. После смыкания ее концов она приобретает форму труб ки, называемой хордой.
С боков от хорды формируются два мешкообразных выроста, -состоящих из мелких клеток с высокой митотической активностью. -Это зачатки третьего зародышевого листка — м е з о д е р м ы . В ней различают спинные сегменты (сомиты), сегментные ножки и несегментированную часть, состоящую из двух листков: наруж-
ного, или париентального, и внутреннего, или висцерального. Полость, ограниченная этими листками, носит название вторич н о й п о л о с т и т е л а .
Первичная энтодерма после выделения из ее состава материа лов будущей хорды и мезодермы называется в т о р и ч н о й , или
ки ш е ч н о й э н т о д е р м о й .
Б.Если яйцеклетка, как это бывает у амфибий, содержит сред нее количество желтка и телолецитальна, при полном неравномер ном дроблении, с многослойной бластулой, у которой бластоцель сдвинут к анимальному полюсу, гаструляция начинается с инва гинации материала бластулы, лежащего на границе между дном и краевой зоной. При этом образуется серповидная бороздка, или дорсальная губа бластопора. Другие губы бластопора загружены желтком и очень инертны. Микромеры бластулы, интенсивно делясь, обрастают макромеры снаружи. Зародыш становится двух слойным, состоящим из эктодермы и энтодермы.
Постепенно длина серповидной бороздки увеличивается, ее концы смыкаются, образуется округлый бластопор, в центре кото рого находится желточная пробка. Она состоит из бластомеров дна бластулы, не инвагинировавших в бластоцель (рис. 49).
Клетки, расположенные на поверхности зародыша, постепенно стягиваются к бластопору, мигрируют в зоне его губ и продвига ются вглубь (рис. 50). Через дорсальную и боковые губы пере мещается презумптивный материал мезодермальных зачатков.
Гаструляция протекает медленнее, чем у ланцетника, поэтому мезодерма не входит в состав первичной энтодермы, а, обособив-*
Рис. 48. Гаструляция и образование осевых органов у ланцетника:
-<64

•А—В — появление серповидной бороздки (а); В, Г, Д — постепенное смыкание? концов серповидной бороздки и образование бластопора; Е — образование жел точной пробки.
шись в виде самостоятельного зачатка, растет между эктодермой и энтодермой. В связи с этим кишечная энтодерма в начале гаструляции имеет вид незамкнутой трубки (рис. 51). Таким образом, при данном типе гаструляции первичная эктодерма отсутствует.
В. Если яйцеклетка полилецитальная, телолецитальпая, а дробление меробластическое (рыбы, рептилии) и образуется дискоидальная бластула, гаструляция протекает путем инвагинации
иделяминации.
Воднослойном овальном зародышевом диске наблюдается массовая миграция клеток в каудальном направлении, поэтому задний край зародышевого диска утолщается и приподнимается над желтком. В этой зоне начинается подворачивание материала внутрь — образуется краевая зарубка. Она соответствует бласто-
пору. Его средняя часть —это дорсальная губа; латеральнее от нее лежат боковые губы, вентральная губа бластопора отсут ствует.
Из материала, инвагинировавшего через край зародышевого диска, образуется энтодерма, прилегающая к неразделенному на бластомеры желтку. Энтодерма отщепляется от зародышевого диска и между энтодермой и желтком формируется полость пер вичной кишки.
Хордомезодермальпый зачаток вначале входит в состав энто дермы, а затем после миграции располагается между эктодермой и энтодермой. Средняя часть мезодермального зачатка — это ма териал будущей хорды, с боков от которой находится материал мезодермы.
66
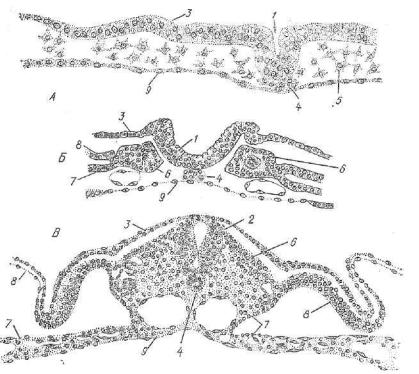
!Рис. 50. Схема гаструляции у амфи бии:
Л—Г — вид со стороны бластопора; I— |
||||
IV — сагиттальные разрезы; 1 — пре< |
||||
кумптивный эпидермис; 2 — вентраль |
||||
ная мезодерма; 3 — презумптивная |
нер- |
|||
ппая система; а — ранняя гаструла; б— |
||||
средняя гаструла; в |
— |
поздняя гастру- |
||
л а (стадия желточной пробки); з — ко |
||||
нец |
гаструляции; |
4— презумптивная |
||
хорда; |
5 — прехордальная пластинка; |
|||
> — |
презумптивная |
|
энтодерма; |
7 — |
бластоцель; 8 — дорсальная губа бла |
||||
стопора; 9 — вентральная губа бласто |
||||
пора; |
ю — желточная |
пробка; |
и — |
|
fi л а стопор; 12 — гастроцель; 13 — сер повидная бороздка. Крестом обозначено положение вегетативного полюса.
Г. У животных с резко полилецитальными и телолецитальными яйцеклетками (птицы) га струляция начинается с делями нации, то есть расщепления за родышевого диска, распластан ного на желтке, на два слоя: по верхностный — эктодерма и внутренний — энтодерма. Даль нейшая интенсивная миграция клеток, расположенных на пери ферии диска, в направлении зад него конца зародыша заверша ется формированием первичной
полоски. На переднем ее конце образуется головной (первичный) узелок. Последний соответствует дорсальной губе бластопора. Бо ковыми губами бластопора является первичная полоска. В этой зоне путем инвагинации клеток обособляется хордомезодермаль пый зачаток, который затем растет в виде клина между эктодермой п энтодермой.
Процесс гаструляции у млекопитающих сходен с гаструляцией птиц.
Таким образом, сравнительный анализ процесса гаструляции свидетельствует о том, что у всех животных он завершается фор мированием зародышевых листков: эктодермы и энтодермы (пер вая фаза) и мезодермы (вторая фаза). Это показатель онтогене тического сходства и филогенетического родства животных. Об этом свидетельствуют и другие процессы: у всех хордовых начало гаструляции связано с дорсальной губой бластопора: инвагина ция у ланцетника, серповидная бороздка у амфибий, краевая за рубка у рыб, рептилий, гензеновский узелок у пресмыкающихся, птиц, млекопитающих (рис. 52). Бластомеры этих зон у всех животных очень активны. По сравнению с другими клетками они вдвое больше содержат гликогена, в них интенсивнее протекают все цитохимические реакции. Дорсальная губа бластопора оказы-
5* |
67 |

I — амфибластула; II—III — гаструляция; IV — нейрула; 1 — эктодерма; S —• энтодерма; 8 — хорда; 4 — мезодерма; б — нервная пластинка; в — верхняя и 7— нижняя губа бластопора; 8 — бластопор; о — гастроцель; 10 — нервная трубка; II — нервный канал; 12 — сегментированная мезодерма; 13 — несегментированная мезодерма; 14 — желточная энтодерма (желточная пробка).
вает влияние, то есть индуцирует развитие других частей зароды ша. Бластомеры, мигрировавшие через дорсальную губу бласто пора, — это материал будущей хорды. Последняя индуцирует раз витие соседних эмбриональных зачатков, а развитие эктодермальной закладки обусловливает развитие нервной трубки.
Много сходного у хордовых в развитии хордомезодермального зачатка, в двухфазности гаструляции-, дифференцировке зароды шевых листков. В процессе развития мезодермы у всех позвоноч ных образуется хорда, сегментированная мезодерма, или сомиты (спинные сегменты), и несегментированная мезодерма, или спланхнотом. Последний состоит из двух листков: наружного — париетального (пристеночного) и внутреннего — висцерального. Пространство между этими листками называется вторичной поло стью тела (рис. 53).
Спинные сегменты могут быть связаны с несегментированной мезодермой сегментными ножками, или нефрогонадотомом. В со-
68
Рис. 52. Сравнение гаструлы амфибий
(Л) с бластодермой куриного зародыша (/>). Стрелки указывают положение и направление миграции клеток хорды и мезодермы:
1 — хорда; г— мезодерма; 8 — бластопор;
4 — первичная полоска.
митах различают три зачатка: дерматом, миотом, спланхнотом.. Из них соответственно развиваются глубокие слои кожи, мышеч ная ткань скелета, костная и хрящевая ткани. Нефрогонадотом — ото зачаток мочеполовой системы, а из спланхнотома образуются внутренняя выстилка серозных оболочек, корковое вещество над почечников, ткани сердца.
Из эктодермы развиваются у всех позвоночных нервная труб ка и кожная, а также внезародышевая эктодерма. Процесс дифференцировки нервной пластинки описан выше.
Краниальная часть нервной трубки расширена, из нее разви ваются нервные пузыри, а затем головной мозг. Нервная трубка— это зачаток всей нервной системы животного.
Клетки эктодермы, граничащие с нервной пластинкой, харак теризуются высокой митотической активностью. Делясь, они по крывают зачаток нервной трубки, в связи с чем последняя оказы вается лежащей под эктодермой. Из кожной эктодермы развива ется самый поверхностный (эпителиальный) слой кожного покро ва и его производные.
Из кишечной энтодермы развивается внутренняя ткань (эпи-
69
ВНЕЗАРОДЫШЕВЫЕ ОРГАНЫ
Внезародышевые, или временные (провизорные), органы получи ли свое развитие в связи с адаптацией животных к новым усло виям обитания в процессе эволюции позвоночных. Первоначально их значение сводилось к запасу и хранению питательного мате риала (желтка). Затем их роль расширилась. Образовались вре менные органы, которые стали выполнять защитную, дыхатель
ную и трофическую функции. Провизорные органы создают вокруг зародыша, развивающегося на суше, водную среду — самую благоприятную для нового организма.
В настоящей главе освещаются только основные закономерно сти строения и функции этих органов. Более подробно они будут изложены в главах, посвященных развитию птиц и млекопитаю щих.
К временным органам позвоночных относят желточный мешок, амнион, аллантоис, серозу, хорион, плаценту. Они развиваются из внезародышевых участков эктодермы, мезодермы, энтодермы. Эти зоны зародышевых листков не принимают участия в закладке ди-
финитивных (окончательных) органов тела |
зародыша. |
У хордовых животных первый временный орган появился у |
|
рыб. Им является ж е л т о ч н ы й м е ш о к . |
В формировании его |
стенки участвуют все зародышевые листки: эктодерма, несегментированная мезодерма и энтодерма. У птиц и млекопитающих в
состав желточного |
мешка |
входят внезародышевые |
энтодерма и |
||
висцеральный листок мезодермы, а эктодерма |
и |
париетальный |
|||
листок мезодермы |
участвуют в формировании |
амниона, |
серозы, |
||
хориона. |
|
|
|
|
|
А м н и о н построен из |
внезародышевых эктодермы и |
парие |
|||
тального листка мезодермы. Он хорошо развит у птиц и млекопи тающих. Создавая вокруг зародыша водную среду, амнион защи щает его от механических повреждений, высыхания, участвует в питании плода у птиц.
С ер о за, или серозная оболочка (ее нельзя путать с сероз1- ными оболочками, покрывающими внутренние органы, лежащие в грудной и брюшной полостях), развита у пресмыкающихся и птиц. В ее состав входят эктодерма и париетальный листок мезо дермы. Она выполняет защитную и дыхательную функции.
Х о р и о н и п л а ц е н т а — это внезародышевые органы мле копитающих. Они возникли в связи с переходом животных к внут риутробному развитию, поэтому основной их функцией является связь зародыша с материнским организмом и его питание. Эти оболочки построены из эктодермы и париетального листка мезо* дермы (см. Развитие млекопитающих).
А л л а н т о и с состоит из внезародышевых энтодермы и вис церального листка мезодермы. Он участвует в газообмене и вы полняет роль мочевого пузыря, так как является местом скопле нии продуктов обмена веществ.
Тшшм образом, в процессе эмбриогенеза выход позвоночных па сушу обусловил развитие временных или внезародышевых ор ганов (плодных оболочек).
ЭМБРИОНАЛЬНОЕ РАЗВИТИЕ ПТИЦ
Половые клетки. Оплодотворение. Дробление. С п е р м и н птиц так же, как и у других представителей позвоночных, имеют жгу тиковую форму и состоят из головки, шейки, хвостового отдела*
71

У разных видов сельскохозяйственных птиц они отличаются раз мером, формой головки и хвостика. Например, верхняя часть головки спермия петуха лопастной формы, а у гусака она вытя нута и заострена на конце.
Продолжительность жизни спермиев в половых путях самки более 30 дней. В течение данного периода без повторного спари вания несушки откладывают оплодотворенные яйца. В первые две недели после спаривания число оплодотворенных яиц наиболь шее.
По количеству и локализации желтка я й ц е к л е т к и |
птиц |
относят к полилецитальным, телолецитальным. В яйце |
птиц |
содержатся: желток, то есть яйцеклетка, белок, подскорлуповая оболочка и скорлупа (рис. 55). Все компоненты яйца, кроме желт ка, относятся к третичным оболочкам. Их материал продуцирует ся железами яйцевода. При продвижении яйцеклетки по яйцево- д^рна покрывается третичными оболочками.
Яйцеклетка, как и другие клетки, имеет ядро и цитоплазму. Ядро, расположенное в анимальном полюсе клетки, окружено
незначительным слоем цитоплазмы, содержащей органеллы. Вся остальная часть яйцеклетки заполнена желтком — клеточным включением. В состав желтка входят протеиды и липиды. В ре зультате концентрации из растворенного состояния они осажда ются и приобретают вид шариков. В начальной стадии желточ ные шарики богаты фосфолипидами, а затем насыщенными жир ными кислотами. У кур в желточных шариках много свободного холестерина.
Желток яйца неоднороден, он бывает светлый и темный. В центре яйцеклетки находится светлый желток. Эта зона назы вается латеброй и по форме напоминает колбу. В остальной части яйцеклетки темный и светлый желток расположен слоями. Обыч но светлый желток откладывается в ночное время, а темный — в другое время суток.
Периферический слой цитоплазмы яйцеклетки называется кор тикальным, а оболочка яйцеклетки, или плазмолемма, — первич ной оболочкой. В белке яйцеклетка подвешена на халавах, направ ленных от желтка к тупому и острому концу яйца. Халазы — это пучки тонких скрученных волоконец, состоящих из плотного бел- ково-углеводного комплекса.
Халазы удерживают желток в центре яйца таким образом, что зародыш всегда оказывается сверху.
Снаружи от желтка располагается белок. По месту его лока лизации различают наружный и внутренний слои, каждый из которых состоит из жидкого и плотного белка. В яичный белок входят вода (87%), протеины, гликопротеиды, свободные угле воды, незначительное количество липидов, золы и других веществ. Для развивающегося зародыша белок — это основной источник питательных веществ и воды среднего периода эмбриогенеза. Белок является поставщиком минеральных веществ, протеинов. Он содержит бактерицидные вещества, убивающие микроорга низмы.
Белок покрыт подскорлуповой оболочкой, в которой различают два слоя: паружный и внутренний. Наружный слой толщиной около 56 мкм состоит из волокон диаметром 2—10 мкм. Этот слой плотно соединен со скорлупой. Толщина внутреннего слоя равна 16 мкм, а диаметр ее волокон 2—3 мкм. По химическому соста ву волокна сходны с кератиноподобным (роговым) веществом. Внутренний слой подскорлуповой оболочки прилегает к белку.
В зоне тупого конца яйца слои подскорлуповой оболочки рас ходятся и образуется воздушная камера, содержащая воздух.
Подскорлуповая оболочка очень плотна, эластична и прони цаема для газов, воды, растворимых соединений. Свойства обо лочки в значительной степени зависят от ее влажности. Влажная подскорлуповая оболочка набухает, в ней увеличиваются поры, роль которых выполняют пространства между волокнами.
Содержимое яйца заключено в скорлупу. Последняя защища ет яйцо от повреждений, ее материал используется зародышем в качестве минеральных веществ при построении скелета. Скорлупа
72 |
73 |
|

имеет поры, через которые происходят газообмен и испарение влаги при развитии зародыша.
В состав скорлупы входят органические и минеральные вещест ва. Из органических веществ построены плотно сплетенные волок на и гранулы белка. Снаружи скорлупа покрыта топкой надскорлуповой пленкой — кутикулой. Построенная из муцина, она пре пятствует проникновению через поры скорлупы микроорганизмов, спор грибов и проницаема для газов.
После выхода из яичника яйцеклетка попадает в яйцевод, где она о п л о д о т в о р я е т с я . При спаривании самца с самкой в яйцевод попадает одновременно несколько миллионов спермиев. Особенностью оплодотворения яйцеклеток сельскохозяйственных птиц является полиспермия — в яйцеклетку может проникать до 300 спермиев, с ядром женской половой клетки соединяется толь ко один спермий. После оплодотворения, продвигаясь по яйцеводу и покрываясь третичными оболочками (белком, подскорлуповой оболочкой, скорлупой), яйцеклетка интенсивно делится митозом. Следовательно, снести яйцо — это зародыш на ранней стадии эмбриогенеза.
Д р о б л е н и е у птиц меробластическое (частичпое) или дискоидальное. В нем принимает участие "й^ъко анимальпый полюс зиготы, где расположены ядро и цитоплазма, лишенная желтка. Вегетативный полюс не участвует в дроблении, так как он загру жен желтком, тормозящим дробление.
Первые две борозды дробления имеют меридиональное направ ление и расположены перпендикулярно друг другу. Меридиональ ные борозды сменяются широтными (рис. 56). Образовавшиеся при дроблении клетки (бластомеры) лежат на желтке в виде од ного слоя — диска, поэтому дробление получило название дискоидального, а образовавшаяся при этом бластула — дискобластулы. Ее крыша и краевая зона состоят из бластомеров диска, а дно — из неразделенного на клетки желтка.
Бластоцель имеет вид узкой щели и сдвинут к анимальному полюсу бластулы (рис. 57).
После оплодотворения яйцо в яйцеводе может находиться в те чение 4—27 ч, поэтому и дроблениел и гаструляция протекают в этом органе.
В снесенном яйце эмбриогенез приостанавливается и вновь возобновляется с момента начала инкубации или насиживания» Гаструляция. Закладка осевых органов. Развитие зародышевых листков. Через 12 ч инкубации в центре зародышевого диска образуется зародышевый щиток. Из этой части зародышевого диска развивается тело зародыша. Остальная часть диска называется незародышевой, так как она используется при построении плод ных оболочек (временных, или провизорных, органов), обеспечи вающих нормальное развитие цыпленка. Зародышевый щиток окружен светлым полем. Его клетки приподняты над желтком. Под ними располагается подзародышевая полость, образовавшая ся вследствие использования зародышем желтка. Светлое поле окружено темным полем. Его клетки интенсивно делятся и раз растаются по поверхности желтка. Эту зону называют краем об
растания.
Г а с т р у л я ц и я у птиц, как и у других позвоночных, про текает в две фазы. Первая фаза: путем расслоения, или делямииации, однослойного диска образуются два зародышевых лист ка — эктодерма и энтодерма (рис. 58). Вторая фаза — формирова ние хордомезодермального зачатка: клетки краевой зоны диско бластулы интенсивно делятся и мигрируют в направлении_ааднего края зародышевого щитка двумя потоками, где они встречаются
У4

и начинают продвигаться вперед по средней линии зародышевого щитка, формируя утолщенный клеточный валик — первичную полоску. Она направлена от заднего края зародышевого щитка вперед.
В- средней зоне первичной полоски образуется углубление — первичная бороздка (рис. 59), на переднем конце которой возни кает утолщение. Это первичный, или гензеиовский, узелок. Он имеет углубление, носящее название первичной ямки (гастрального мешочка).
Первичная полоска идентична бластопору гаструлы ланцетни ка и амфибий. Первичная ямка гензеновского узелка — это гомо лог дорсальной губы бластопора, остальные зоны первичной по лоски гомологичны боковым и вентральной губам.
Взоне первичной ямки зародышевый материал инвагинирует
ив виде клеточного тяжа перемещается между эктодермой и энто дермой к переднему концу зародыша. Этот тяж называется голов ным, или хордальным, отростком, из него развивается хорда (рис.60).
Через первичную бороздку в направлении переднего конца зародыша мигрируют два крылообразных зачатка. Они растут с боков от хорды между эктодермой и энтодермой. Дифференциров-
Рис. 59. Гаструляция у птиц:
А — миграция клеток в зародышевом щитке; В — обра зование хордомезодермального зачатка; 1 — эктодер ма; 2 — материал будущей нервной пластинки; з — ма териал хордальной пластин ки; 4 — первичный (голов ной) узелок; 5 — первичная ямка; в — первичная полос ка; 7 — первичная бороздка; 8 — хорда; 9 — мезодерма.
Сплошные стрелки обозначают направление перемещения
материала в составе наруж ного, а пунктирные — в со ставе среднего зародышевого листка (по Кнорре).
ка этих зачатков заканчивается образованием среднего зародыше вого листка — мезодермы.
Таким образом, и у птиц гаструляция завершается з а к л а д к о й з а р о д ы ш е в ы х л и с т к о в . В гаструляции птиц отме чают дальнейшее усложнение в развитии позвоночных: зачаток хорды и мезодермы полностью утратили связь с первичной киш кой. Последняя очень рано отделяется от других эмбриональных зачатков и входит в состав энтодермы зародышевого щитка.
Дальнейшее р а з в и т и е , зародышевых листков у птиц про текает, как и у других позвоночных (см. рис. 53). Из эктодермы выделяется нервная пластинка, а после соединения ее краев фор мируется нервная трубка. Эктодерма наползает снаружи на нерв ную трубку, в связи с чем последняя оказывается погруженной под эктодерму. Нервная трубка — это источник развития всей нервной системы, а эктодерма — зачаток поверхностного слоя кож ного покрова (эпидермиса).
Мезодерма подразделяется на сомиты (сегментированную ме зодерму), сегментные ножки, несегментированную мезодерму (спланхнотом). В состав сомитов входят дермотом, миотом, склеротом. Из дермотома развиваются глубокие слои кожного покрова, из миотома — мышечная ткань скелета, из склеротома — скелет. Сегментные ножки — источник развития мочевыделительной си стемы.
В спланхнотоме различают париетальный (наружный) и вис-
77
А, В, В — три последовательные стадии; 1 — нервный желобок; 2 — нервная труб ка; з — эктодерма; 4 — хорда; 5 — мезодерма; в — сомиты; 7 — висцеральный лис ток спланхнотома; 8 — париетальный листок спланхнотома; 9 — кишечная энто дерма.
церальный (внутренний) листки, между которыми находится це лом — вторичная полость тела. Из спланхнотома развивается эпи телиальная ткань, покрывающая серозные оболочки внутренних органов, грудную и брюшную полости (рис. 61).
Образование внезародышевых органов. Развитие тела зароды ша сопровождается образованием временных, или внезародышевых (провизорных), органов, способствующих созданию необходи мых условий для нормального развития зародыша. К провизорным органам у птиц относят: желточный мешок, амнион, серозу, аллантоис. Они построены также из зародышевых листков — эктодер мы, энтодермы, листков несегментированной мезодермы. Источни ком развития этих листков является внезародышевая зона зароды шевого диска (цв. табл. 1).
Темпы развития временных органов часто могут опережать раз витие зародыша, у птиц по сравнению с другими животными этот процесс протекает более интенсивно.
Образование провизорных органов начинается с разрастания по поверхности желтка периферической зоны внезародышевого 78
материала бластодиска. Эта зона называется зоной обрастания, после завершения гаструляции она состоит из внезародышевых эктодермы, энтодермы и листков несегментированной мезодермы.
В дальнейшем, при развитии плодных оболочек, эктодерма все гда связана с париетальным листком, а энтодерма — с висцераль ным листком мезодермы.
Ранее других плодных оболочек образуется желточный мешок, формирующийся в процессе обрастания желтка зародышевыми листками. Он выполняет трофическую функцию и построен из внезародышевых энтодермы и висцерального листка мезодермы. Энтодерма желточного мешка участвует в ферментативном рас щеплении желтка и всасывании питательных веществ в кровенос ные сосуды, расположенные в его мезодермальном слое. Так как яйцеклетка птиц содержит большое количество желтка, эта плод ная ободочка развита очень хорошо. Желточный мешок отпгауровывается от тела зародыша туловищной складкой. Ее образование обусловлено интенсивным разрастанием клеток всех зародышевых листков, лежащих на границе зародышевого и незародышевого материала. В туловищной складке участвуют все зародышевые листки этой зоны. Острие складки направлено под туловище заро дыша, в связи с чем зародыш приподнимается над желтком, а его туловищная энтодерма, ранее распластанная на желтке, образует кишечную трубку. Последняя сообщается с желтком посредством узкого желточного протока.
Туловищная складка первоначально развивается в головной части зародыша, а затем распространяется в каудальном направ лении.
Амнион (amnion — чаша), или водная оболочка. Сероза. Одно временно с туловищной складкой, отграничивающей зародышевую от незародышевой части, образуется амниотическая складка. Ее гребень направлен над телом зародыша. Возникая сначала над головой зародыша, она постепенно захватывает туловище. Голов ная часть амниотической складки образуется очень быстро, по этому ее стенка состоит только из эктодермы. После срастания краев головной части амниотической складки возникает проамнион. Туловищная зона амниотической складки отстает в развитии. В ее состав входит не только эктодерма, но и париетальный листок мезодермы.
Над туловищем зародыша края амниотической складки срас таются: эктодерма с эктодермой, париетальный листок с парие тальным листком мезодермы. После срастания краев амниотиче ской складки тело зародыша оказывается окруженным двумя плодными оболочками — амнионом и серозой.
В формировании а м н и о н а принимают участие эктодерма и париетальный листок мезодермы внутренней половины амниоти ческой складки, второй — эктодерма и париетальный листок наг ружной половины амниотической складки. Края хвостовой части •амниотической складки не срастаются, здесь образуется канал, пшзывающий полость амниона с остальной частью яйца. Этот
79
канал получил название серо-амниотического канала. Эктодерма амниона продуцирует амниотическую жидкость, которая заполня ет амниотическую полость, создавая для развивающегося зароды ша самую благоприятную водную среду. Амнион выполняет за щитную функцию, он сглаживает удары и создает для эмбриона возможность некоторой подвижности.
С е р о з а — сама поверхностная оболочка зародыша. Приле гая к скорлупе, она покрывает снаружи амнион с расположенным в нем зародышем и желточный мешок с желтком.
Основными функциями серозы является участие в газообмене и переносе ионов кальция из скорлупы в тело зародыша. Элек тронно-микроскопическое строение клеток эктодермы серозы сходно с клетками, продуцирующими хлориды: плазмолемма снаб жена многочисленными микроворсинками, в цитоплазме очень много митохондрий. Эти клетки синтезируют соляную кислоту, действие которой направлено, очевидно, на растворение солей кальция скорлупы, которые затем переводятся в организм плода.
Аллантоис (allantois — колбасообразный) образуется после завершения формирования кишечника на вентральной поверхно сти задней кишки. Он, как и кишечник, состоит из энтодермы и висцерального листка мезодермы. Аллантоис постепенно заполня ет все щели между амнионом, желточным мешком и серозой, срастаясь с мезодермальным листком последней. В этом участке формируется густая сеть кровеносных сосудов, что способствует улучшению снабжения организма кислородом.
Таким образом, прилегая к скорлупе, аллантоис участвует в газообмене. Он выполняет функцию выделительного органа, так как в нем скапливаются продукты обмена веществ.
Все провизорные органы в конце эмбриогенеза редуцируются. Из приведенного материала следует, что развитие птиц также про текает стадийно. Нарушение типичного течения одной из стадий влечет за собой изменения всего хода развития зародыша. Зна ние стадийности развития позволяет выделить наиболее важные периоды в формировании зародыша. Понимание их крайне необ ходимо в связи с задачами практического птицеводства.
Однако в настоящее время периодизация в развитии птиц еще недостаточно разработана. Наиболее детально изучено развитие куриного зародыша.
Н. П. Третьяков и М. Д. Попов, основываясь на характере пи тания зародыша, разработали следующую классификацию стадий развития цыпленка.
1-я с т а д и я — латебральное питание — протекает первые 30—36 ч. Материалом для питания служит желток латебры, со держащий белки, соли, воду. Источником энергии является глико ген, поэтому зародыш почти не нуждается в кислороде. В орга низме отсутствует кровообращение.
2-я с т а д и я — желточное питание и наличие желточного кру га кровообращения — длится с 30—36 ч до 7—8 дней инкубации. К 30-му часу инкубации закладываются сердце и желточный ме*
80
шок, в последнем развиваются кровеносные сосуды. Запас глико гена почти полностью исчезает. В организм поступает кислород,. что облегчает использование белков и жиров. Формируются к другие плодные оболочки, а также органы тела: развивается нерв ная система, начинается биение сердца и сокращение мускулату ры тела. Закладка печени обусловливает начало синтеза мочеви ны, поэтому продукты распада белков становятся менее вредными для зародыша.
3-я с т а д и я — дыхание кислородом воздуха и питание бел ком яйца — продолжается с 7—8-го дня по 18—19-й день. К кон цу ее зародыш становится сформированным плодом. Для этой стадии характерны интенсивное развитие и функционирование аллантоиса, в стенке которого формируется густая сеть кровенос ных сосудов. Прилегая к серозе и вместе с ней к подскорлуповой оболочке яйца, аллантоис обеспечивает снабжение кислородом развивающиеся ткани. Благодаря растворению скорлупы возрас тает интенсивность минерального обмена, усиливается усвоение жиров.
4-я с т а д и я — потребление кислорода воздуха из воздушной камеры яйца. Цыпленок переходит на легочное дыхание, начина ет функционировать малый круг кровообращения, появляется ар териальная кровь, обогащающая ткани кислородом. Продолжается стадия с 18—19-го дня до наклева, К этому моменту аллантоис подвергается обратному развитию, и цыпленок начипает испыты вать недостаток в кислороде. Он проклевывает внутренний листок подскорлуповой оболочки и начипает дышать воздухом, содержав шимся в воздушной камере яйца.
5-я с т а д и я — вылупление — длится с 20-х по 21-е сутки.. Цыпленок питается остатками желтка, которые вместе с желточ ным мешком втягиваются в полость кишки. Другие зародышевыеоболочки также прекращают функционировать и отмирают. Цып ленок проклевывает скорлупу и покидает яйцо.
Существуют и другие классификации. Согласно разработанной Г. А. Шмидтом и М. Н. Рогозиной периодизации, основанной на эмбриогенезе куриного зародыша, различают четыре периода раз вития: зародышевый, предплодный, нлодный, а также период вы ступления.
З а р о д ы ш е в ы й п е р и о д длится первые восемь дней, из которых начальные стадии развития протекают во время продви жения зародыша по яйцеводу. Зародышевый период характеризу ется интенсивно протекающими процессами формообразования. Источником питания зародыша является желток и частично сме шанная с ним жидкая фракция белка. После образования желточ ного мешка транспортировка питательных веществ осуществляется с помощью его кровеносных сосудов. Дыхание зародыша сна чала протекает осмотическим путем, а затем обеспечивается сосу дистой системой желточного мешка. В конце зародышевого перио да в этом процессе принимают участие кровеносные сосуды аллан тоиса.
6 Заказ Ns 908 |
81 |