

Конспект_лекцій_Заг.вірусологія_0
.pdfПісля відкриття вірусів, була відома єдина їх фізико-хімічна особливість – здатність проходити через бактерійні фільтри. По суті, це була оцінка розміру віріонів вірусів, яка ставила їх окремо від мікроорганізмів. У той час визначення інакших властивостей вірусів було неможливим, тому дослідження були спрямовані на вивчення інфекційного процесу і реакції на зараження з боку організму. З цієї причини перші спроби класифікації ґрунтувалися на схожості патогенних властивостей вірусу, наприклад, виділяли «віруси гепатиту», або на їх органотропності, наприклад «респіраторні віруси». У міру проникнення в природу вірусів, з 1950-х років робилися різноманітні спроби їх класифікації, на основі різноманітних властивостей. Були запропоновані ціла низка класифікацій, часто взаємовиключних.
Існує три основні гіпотези щодо походження вірусів:
1)від доклітинних форм життя (автори: вірусологи А.О. Смородінцев, А.С. Кривицький, 1953; В.М. Жданов, 1953);
2)від бактерій (автори: французький мікробіолог, Нобелівський лауреат Ш.Ж.А. Ніколль, 1925; американський вірусолог Р. Грін, 1935; австралійський вірусолог та імунолог, Нобелівський лауреат Ф.М. Бернет, 1943);
3)від компонентів клітини (автори: американські вірусологи, Нобелівський лауреат С. Лурія і Дж. Дарнелл, 1967).
Таксономічна класифікація вірусів
В основу сучасної класифікації вірусів покладено такі основні критерії: 1)тип нуклеїнової кислоти (ДНК або РНК), її структура (кількість
ниток); 2)наявність зовнішньої ліпопротеїнової оболонки;
3)стратегія вірусного геному (механізм реплікації); 4)розмір і морфологія віріона, тип симетрії, кількість капсомерів; 5)форми генетичних взаємодій; 6)спектр сприйнятливих хазяїв;
7)патогенність, у тому числі цитопатичні зміни та утворення тілецьвключень у клітинах;
8)географічне поширення;
9)спосіб передавання;
10) антигенні властивості.
В 1966 році був створений Комітет по номенклатурі вірусів. В 1973 році його перейменували в Міжнародний комітет по таксономії вірусів МКТВ (International Committee on Taxonomy of Viruses ICTV). Завдання даного комітету є підготовка та видавництво праць по удосконаленню таксономії та номенклатурі вірусів. Правила що встановлює ця організація є обов’язковими для всіх наукових публікацій в міжнародних журналах.
Існувало багато варіантів класифікацій вірусів взагалі, наприклад класифікації по Брауну, Метьюзу, Балтімору. На даний час однією із нових являється класифікація по Франкі (1991).
21
Нині відомо понад 3600 видів вірусів хребетних, безхребетних, рослин, грибів, найпростіших і бактерій. Із них 1550 класифіковані в 3 порядки, 56 родин, 9 підродин і 233 роди. З урахуванням штамів і серотипів налічується понад 30 000 вірусів. Віруси хребетних входять у 2 порядки, 28 родин, із яких 10 —ДНК-вмісні і 18 — РНК-вмісні, 7 підродин і 85 родів.
Всі віруси належать до царства – Vira. Це царство розподіляється на РНК та ДНК вмістні віруси. Надалі віруси розподіляються на одно та дволанцюгові по структурі нукленової кислоти, або від заряду таких – негативний геном чи позитивний. Також іде розподілення по морфології – нитковидні, видовжені чи ізометричні. Але також слід замітити, що до одних і тих же груп можуть відноситись віруси, що реплікуються через фермент полімеразу (+РНК або ДНК віруси), або через реверстранскриптазу (-РНК чи ДНК віруси). Більшість вірусів, як відомо, складаються із нуклеїнової кислоти та білкового капсиду, що її вкриває, але є розподілення також на такі, що мають суперкапсид.
Деякі віруси мають свій геном в одній часточці (моногеномні), але існують і такі, що мають свій геном в багатьох частках вірусів (мультигеномні). Він розподіляється на дві, три рідше на чотири часточки.
Інші характеристики в класифікації вірусів включають в себе симетрію спіралі у ізометричних вірусів, розміри вірусів, а також інші фізичні, хімічні та біологічні властивості.
Порядок вірусів є зібрання родин з загальними характеристиками, що відрізняються від інших порядків та родин. Він позначається іменуванням з суфіксом –virales. Віруси рослин об’єднані в три порядка: Caudovirales , що включає родини Myoviridae, Siphoviridae, Podoviridae ; порядок Mononegavirales включає в себе Paramyxoviridae, Phabdoviridae, Bornaviridae і Filoviridаe; порядок Nidovirales який включає роди Coronaviridae і Alteriviridae.
Родини, підродини об’єднують роди вірусів з загальними характеристиками, що відрізняються від властивостей інших родин. Вони позначаються суфіксом –viridae. Більшість родин відрізняються від інших морфологією віріона, структурою геному, або стратегією реплікації.
Роди вірусів представляють собою групу вірусів, що мають загальні характеристики, які мають загальні характеристики і відрізняються від вірусів інших родів. Вони позначаються суфіксом –virus.
Вид вирусів. Видовий таксон є найбільш важливою одиницею в системі класифікації. Визначення виду вірусу «Вірусний вид є політипічною категорією, що складає схожу лінію реплікації і займає особливу екологічну нішу”. Члени цього класу визначаються більше ніж по одному признаку.
Фітовірусологи вважають, що концепція видів для фітовірусів являється не досить досконалою. В основному із-за механізму розмноження – на відміну від багатьох інших груп організмів.
Практично кожного тижня у світі стає відомо про нові і нові віруси, серед яких вагоме місце займають фітовіруси. Такі віруси мусять бути класифіковані і віднесені до певної вірусної групи. Певний час досліджує мі
22
віруси являються не класифікованими і досконально вивчаються. Не слід забувати, про високий рівень штамоспецифічності серед вірусів. Нерідко нові віруси можуть являти собою якийсь окремий ізолят вже відомого вірусу.
В даний час для таксономічних цілей використовується багато характеристик вірусів: морфологія (розмір, форма віріона, наявність оболонки, структура капсиду), фізико-хімічні властивості (молекулярна маса, коефіцієнт седиментації, стійкість до розчинників), тип геномної НК, розмір генома в тисячах пар основ, кількість ниток НК, лінійність або кільцевидність, нуклеотидну послідовність, співвідношення Г+Ц, властивості білків (кількість, розмір та функціональна активність структурних білків, наявність ферментів, амінокислотна послідовність).
Більшість фітовірусів мають назву, що походить від зовнішніх симптом захворювання, що вони викликають. Віруси рослин угруповані на відміну від інших вірусів у групи. Назва вірусологічних груп походить як правило від першого хазяїну, де їх було виявлено. Наприклад, вірус викликаючий мозаїчне захворювання тютюну – названий вірус тютюнової мозаїки (ВТМ) (tobacco mosaic virus), а вірусологічна група таких вірусів названа – Тобамовірус, вірус, що викликає бронзовість томатів – tomato sрotted wilt virus – Тосповірус, і т.д.
Конгресом було вирішено забороняти давати фітовірусам офіційні назви, але віруси досі називають по-різному, нерідко по-місцевому, в основному на англійській мові. Такі назви часто включають імена або основну рослину, на якій його вперше було виявлено.
Як уже зазначалося найбільш направляючими дослідженнями по класифікації вірусів являються сіквенс нуклеотидів. Такі дослідження дають змогу уявити про дійсний еволюційний зв’язок серед царства Vira.
ЛЕКЦІЯ 5. ОСНОВИ РЕПРОДУКЦІЇ ВІРУСІВ
Загальні поняття про репродукцію вірусів.
Біологічні та генетичні особливості механізмів репродукції вірусів. Стадії репродукції вірусів
Пошуки етіологічних агентів інфекційних захворювань увінчалися відкриттям сотень вірусів. Патологічні ефекти при вірусних захворюваннях складаються з взаємодії декількох факторів: а) токсичного впливу продуктів вірусних генів на метаболізм заражених клітин; б) реакції хазяїна на експресію вірусних генів у заражених клітинах; в) модифікації експресії генів хазяїна в результаті їх структурної чи функціональної взаємодії з генетичним матеріалом вірусу. У більшості випадків симптоми гострих вірусних захворювань можуть бути безпосередньо зв'язані з руйнуванням клітин вірусом.
23
Загальні поняття про репродукцію вірусів.
Віруси відтворюють собі подібні частки в такій величезній кількості і настільки своєрідними способами, що це явище стали іменувати репродукцією, тому що тут копіюються молекули нуклеїнових кислот і, відповідно до укладених в них генетичної інформації, синтезуються вірусні білки.
1.Для того щоб вірус міг розмножитися, він повинний спочатку заразити клітину. Спектр хазяїнів визначається як типами клітин, так і видами тварин, яких він може заражати й у який він здатний розмножуватися. Спектр хазяїнів різних вірусів значно варіює. Одні віруси мають широкий спектр хазяїнів, інші заражають лише клітини одного типу визначених видів. Здатність об'єкта заражатися називають сприйнятливістю. Коли людина вступає в контакт із вірусом, у спектр хазяїнів якого він входить, негайно заражаються лише сприйнятливі клітини, що знаходяться у вхідних воріт інфекції. Однак для клінічного прояву інфекції зараження цих клітин може виявитися недостатньо. Як правило, хвороба виникає внаслідок зараження клітин-мішеней (наприклад, клітин центральної нервової системи) вірусом, що розмножився в чуттєвих клітинах у місці проникнення інфекції.
Убагатьох випадках (наприклад, респіраторні інфекції) клітини-мішені розташовуються у вхідних воріт інфекції.
2.На початку інфекції вірус вводить у клітину свій генетичний матеріал — РНК чи ДНК, часто разом з необхідними білками. Розміри, склад і генна організація вірусних геномів дуже сильно варіюють. Віруси, очевидно, еволюціонують різними шляхами, і не існує якого-небудь одного переважного способу їхньої реплікації. Тут варто підкреслити два положення. По-перше, здатність вірусів до розмноження і доля заражених клітин залежать від синтезу і функції продуктів вірусних генів — білків. Ніде кореляція між структурою і функцією — у даному випадку між складом і розташуванням генетичного матеріалу і механізмом експресії вірусних генів
— не є настільки очевидною, як у вірусів. Розходження механізмів, за допомогою яких віруси забезпечують синтез своїх білків, відбито в їхній генетичній структурі, але далеко не завжди може бути виведене з її. Подруге, хоча віруси значно розрізняються по числу генів, що містяться в них, можна сказати, що усі віруси кодують функції трьох типів, які виражаються специфічними для них білками. Вірусні білки забезпечують: а) реплікацію вірусного генома, б) упакування генома у вірусні частки (віріони) і в) зміна структури і (чи) функції заражених клітин.
Стратегія, яка застосовується вірусами для забезпечення цих функцій, варіює. Іноді (паповавіруси) вірусні білки просто допомагають ферментам клітини хазяїна реплікувати вірусний геном. У більшості випадків (пікорнавіруси, герпесвіруси, реовіруси) вірусні білки самі забезпечують реплікацію вірусного генома, але навіть найбільш незалежні віруси використовують при цьому принаймні кілька білків хазяїна. У всіх випадках саме вірусні білки відповідальні за упакування генома у віріони, навіть якщо білки хазяїна і поліаміни зв'язуються з вірусним геномом (наприклад, у
24
паповавірусів) до чи під час формування вірусних часток. Наслідки розмноження вірусу в клітині можуть варіювати від загибелі клітини до незначних, але потенційно дуже важливих змін її функції й антигенної специфічності.
Наші знання щодо циклів репродукції вірусів отримані головним чином з аналізу подій, що відбуваються в синхронно інфікованій культурі клітин. Про віруси, що не вдається виростити в культурі клітин, мало що відомо; у майбутньому, імовірно, вирішенню цього питання допоможе молекулярне клонування. Цикл репродукції усіх вірусів має декілька загальних рис. Незабаром після зараження і протягом декількох годин після нього вдається знайти лише невелику кількість батьківського вірусу. Цей період відомий як екліпс фаза; у цей час геном вірусу взаємодіє з хазяйським чи вірусним апаратом, необхідним для його експресії, але чисельність потомства вірусу ще не перевищує фонового рівня. Потім наступає інтервал, під час якого віріони потомства накопичуються усередині чи поза клітиною з експоненціальною швидкістю. Цей період відомий як фаза дозрівання. Через кілька годин у клітинах, заражених літичними вірусами, знижується метаболічна активність і вони втрачають структурну цілісність. Клітини, заражені іншими вірусами, можуть продовжувати синтез віріонів необмежений час. Цикл репродукції вірусів варіює від 6—8 годин (пікорнавіруси) до 40 годин і більш (деякі герпесвіруси). Врожай вірусу в розрахунку на одну клітину широко варіює й у випадку вірусу поліомієліту, наприклад може перевищувати 100000 часток.
Зараження сприйнятливих клітин не означає, що неминуче буде відбуватися розмноження вірусу і нагромадження його потомства. Ця концепція, одна з найважливіших у вірусології, виникла в останнім часом і її варто розглянути детальніше. Інфекція сприйнятливих клітин може бути продуктивною, обмеженою й абортивною. Продуктивна інфекція відбувається в пермісивних клітинах і характеризується продукцією інфекційного потомства. Абортивна інфекція може наступити в силу двох обставин. Незважаючи на сприйнятливість до зараження, клітини можуть виявитися непермісивними, тому що, найчастіше по невідомих причинах, у них здатні експресуватися лише деякі вірусні гени. Абортивна інфекція може бути також результатом зараження як пермісивних, так і непермісивних клітин дефектними вірусами, у яких відсутній повний набір вірусних генів. Нарешті, клітини можуть бути тільки тимчасово пермісивними, унаслідок чого вірус або зберігається в клітинах до моменту, коли вони стають пермісивними, або в будь-який даний момент вірусне потомство утвориться тільки в деяких клітинах популяції. Цей вид інфекції одними дослідниками був визначений як рестриктивний (restrictive), іншими - як обмежений (restringent). Дана класифікація важлива; її значення обумовлене тим, що цитолітичні віруси, як правило руйнуючі пермісивні клітини під час продуктивної інфекції, можуть просто ушкоджувати, але не руйнувати абортивно заражені пермісивні і непермісивні клітини. Внаслідок ушкодження може відбуватися така експресія хазяйських функцій, у
25
результаті якої клітина перетворюється з нормальної в злоякісну. Додатковим наслідком обмеженої й абортивної інфекцій є персистенція (збереження в клітині) вірусного генома.
Біологічні та генетичні особливості механізмів репродукції вірусів.
Перша і принципова особливість вірусів, що відрізняє їх від інших організмів, це та, що геноми їх представлено молекулами ДНК, чи РНК.
Друга особливість полягає у великій розмаїтості структури і форми їх геномів (одно -, двоспіральні ДНК, одно -, двоспіральні РНК, кільцеві форми нуклеїнових кислот). Геноми РНК-містких вірусів можуть бути представлені як однією молекулою, так і декількома (до 12) різними молекулами РНК. Такі геноми прийнято називати фрагментованими.
Третя особливість вірусів полягає в тому, що майже усі вірусні РНК здатні реплікуватися незалежно від ДНК клітини, тоді як клітинні РНК (рРНК, іРНК, тРНК) синтезуються на матриці клітинної ДНК.
Четверта особливість — диз'юнктивний (роз'єднаний у часі і просторі) біосинтез їх структурних компонентів. Так, наприклад, у складнихвірусів, що відносяться до родин Poxviridae, Paramyxoviridae, Orthomyxoviridae, Rhabdoviridae, синтез суперкапсиду і капсиду нуклеоїду роз'єднаний у часі і проходить у дві стадій. При цьому вірусна нуклеїнова кислота може бути транскрибована безпосередньо в складі нуклеокапсида чи нуклеоїда. Процес транскрипції в цьому випадку здійснюється за допомогою вірусних РНКполімераз, що входять до складу нуклеоїду. Нуклеїнова кислота вірусу може реплікуватися, наприклад, у ядрі чи ядерці, білок може синтезуватися в цитоплазмі, а зборка цілих віріонів чи нуклеокапсидів може відбуватися на внутрішній поверхні цитоплазматичної мембрани. Нарешті, складний ліпопротеїновий суперкапсид може набуватися вірусами в процесі брунькування.
П'ята кардинальна особливість: віруси не мають власних білоксинтезуючих систем, а використовують системи клітини.
Та, нарешті, шоста особливість вірусів полягає у великій розмаїтості самих механізмів реплікації їх вірусних нуклеїнових кислот і репродукції вірусних часток.
Реплікацію нуклеїнових кислот вірусів здійснюють ферменти. Використовуючи нуклеотиди (мономери) клітини, ферменти будують з них полінуклеотидні ланцюги нових молекул нуклеїнових кислот вірусів. Ферменти ці носять загальне найменування — полімерази. У залежності від типу синтезованих нуклеїнових кислот вони називаються ДНК-полімеразами чи РНК-синтетазами. Вірусні полімерази (РНК-реплікази) строго специфічні.
У деяких випадках нуклеїнові кислоти вірусів реплікуються клітинними полімеразами — ферментами, що є присутніми у клітині до її зараження вірусом. Однак частіше реплікацію вірусних нуклеїнових кислот здійснюють полімерази, що з'являються після зараження клітини вірусом. Такі полімерази іменують вірусоспецифічними, тому що біосинтез їх закодований у структурі нуклеїнових кислот самих вірусів. І нарешті,
26
реплікація нуклеїнових кислот деяких вірусів здійснюється за рахунок присутніх у віріоні полімераз. Синтез вірусоспецифічних полімераз, як і вірусних структурних білків, здійснюється на рибосомах клітини відповідно до інформації, що закодована у вірусоспецифічних РНК.
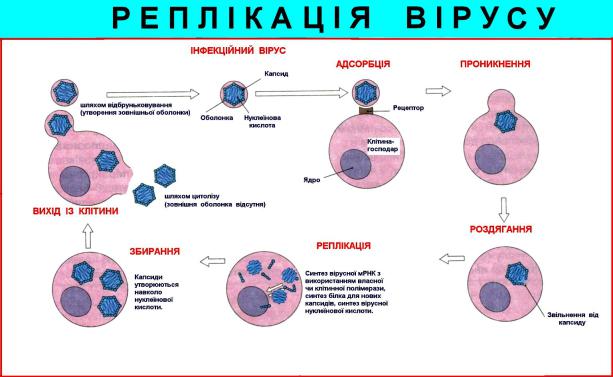
Стадії репродукції вірусів
Процес репродукції вірусів може бути умовно розділений на дві фази. Перша фаза охоплює події, що ведуть до адсорбції і проникнення вірусу в клітину, звільненню його внутрішнього компонента і модифікації його таким чином, що він здатний викликати інфекцію. Відповідно перша фаза містить у собі три стадії: 1) адсорбцію вірусу на клітинах; 2) проникнення в клітини; 3) роздягання вірусу в клітині. Ці стадії спрямовані на те, щоб вірус був доставлений у відповідні клітинні структури і його внутрішній компонент був звільнений від захисних оболонок. Як тільки ця мета досягнута, починається друга фаза репродукції, протягом якої відбувається експресія вірусного генома. Ця фаза складається з п'яти стадій:
1) транскрипції; 2) трансляції іРНК; 3) реплікації генома; 4) збірки вірусних компонентів 5) вихід вірусу з клітини.
Схема реплікації вірусу в клітині. За А.Ройтом, 2000
27
ЛЕКЦІЯ 6. ПРОНИКНЕННЯ ВІРУСІВ В КЛІТИНУ
Адсорбція віріонів на поверхні клітини.
Проникнення вірусів у клітину. Рецепторний ендоцитоз. Злиття вірусної і клітинної мембран.
Роздягання вірусу в клітині.
Адсорбція віріонів на поверхні клітини.
Прикріплення вірусних часток до поверхні клітини-хазяїна — перша стадія інфекційного процесу. Початковий контакт вірусу з клітиною відбувається в результаті випадкового зіткнення по типу броунівського руху.
В основі адсорбції лежать два механізми. Перший з них (неспецифічний) визначається силами електростатичної взаємодії, що виникають між різнойменно зарядженими групами, розташованими на поверхні клітини і вірусу. У цьому процесі беруть участь заряджені позитивно амінні групи вірусного білка і кислі фосфатні, сульфатні і карбоксильні групи клітинної поверхні, що мають негативний заряд. Другий - специфічний. Специфічність зв'язку між вірусом і клітиною обумовлена комплементарними клітинними і вірусними рецепторами.
Процес адсорбції можливий при наявності відповідних рецепторів на поверхні клітки і «впізнаючих” їх субстанцій на поверхні вірусу. Впізнавання клітинних рецепторів вірусними білками (рецепторами), що веде до прикріплення вірусної частки до клітини, є високоспецифічним процесом. Білки на поверхні вірусу, що впізнають специфічні угруповання на плазматичній мембрані клітини і зумовлюють прикріплення до них вірусної частки, називаються прикріпними білками (рецепторами).
Віруси використовують клітинні рецептори, призначені для проходження в клітину необхідних для її життєдіяльності речовин: живильних речовин, гормонів, факторів росту і т.д. Рецептори можуть мати різну хімічну природу і являти собою білки, вуглеводний компонент білків і ліпідів, ліпіди. Рецепторами для вірусів грипу і параміксовірусів є сіалова кислота в складі глікопротеїдів і гліколіпідів, для рабдо- і реовірусів - також вуглеводний компонент у складі білків і ліпідів, для пікорна- і аденовірусів - білки, для деяких вірусів - ліпіди. Специфічні клітинні рецептори відіграють роль не тільки в прикріпленні вірусної частки до клітинної поверхні. Вони визначають подальшу долю вірусної частки, її внутрішньоклітинний транспорт і доставку у визначені ділянки цитоплазми і ядра, де вірус здатний ініціювати інфекційний процес. Вірус може прикріпитися і до неспецифічних рецепторів і навіть проникнути в клітку, однак тільки прикріплення до специфічного рецептора приведе до виникнення інфекції.
Прикріплення вірусної частки з клітинної поверхні спочатку відбувається шляхом утворення одиничного зв'язку вірусної частки з рецептором. Однак таке прикріплення неміцне, і вірусна частка може легко відірватися від клітинної поверхні (оборотна адсорбція). Для того щоб наступила необоротна адсорбція, повинні з'явитися множинні зв'язки між
28
вірусною часткою і багатьма молекулами рецепторів, тобто повинне відбутися стабільне мультивалентне прикріплення. Кількість молекул клітинних рецепторів у ділянках адсорбції може доходити до 3 тис. Стабільне зв'язування вірусної частки з клітинною поверхнею в результаті мультивалентного прикріплення відбувається завдяки можливості вільного переміщення молекул рецепторів у ліпідному шарі плазматичної мембрани, що визначається рухливістю білково-ліпідного шару. Збільшення плинності ліпідів є одним з найбільш ранніх подій при взаємодії вірусу з клітиною, наслідком якого є формування рецепторних полів у місці контакту вірусу з клітинною поверхнею і стабільне прикріплення вірусної частки до виниклого угрупованням — необоротна адсорбція.
Вірусні прикріпні білки (вірусні рецептори). Прикріпні білки можуть знаходитися в складі унікальних органел, таких, як структури відростка в Т- бактеріофагів чи фібри в аденовірусів, що добре видні в електронному мікроскопі; можуть формувати морфологічно менш виражені, але не менш унікальні структури білкових субодиниць на поверхні вірусних мембран, як, наприклад, шипи в оболонкових вірусів, «корону» у коронавірусів.
Простіі віруси тварин містять прикріпні білки в складі капсида. У складних вірусів ці білки входять до складу суперкапсида і представлені множинними молекулами.
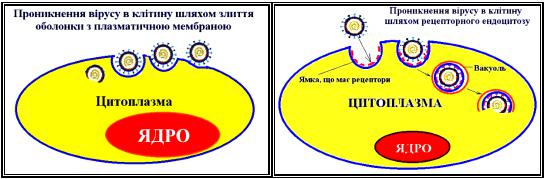
Проникнення вірусів у клітину. Рецепторний ендоцитоз.
Історично склалося представлення про два альтернативні механізми проникнення в клітину вірусів тварин — шляхом віропексису (ендоцитозу) і шляхом злиття вірусної і клітинної мембран. Однак обоє ці механізми не виключають, а доповнюють один одного. Термін «віропексис», запропонований у 1948 р. Фазекасом де Сен Гро, означає, що вірусна частка попадає в цитоплазму в результаті інвагінації ділянки плазматичної мембрани й утворення вакуолі, що містить вірусну частку.
Рецепторний ендоцитоз. Віропексис являє собою окремий випадок рецепторного чи адсорбційного ендоцитозу. Цей процес є звичайним механізмом, завдяки якому в клітину надходять живильні і регуляторні білки, гормони, ліпопротеїни й інші речовини з позаклітинної рідини. Рецепторний ендоцитоз відбувається в спеціалізованих ділянках плазматичної мембрани, де маються спеціальні ямки, покриті з боку цитоплазми особливим білком з великою мол. масою — клатрином.
29
На дні ямки розташовуються специфічні рецептори. Ямки забезпечують швидку інвагінацію й утворення покритих клатрином внутрішньоклітинних вакуолей. Напівперіод проникнення речовини усередину клітини по цьому механізмі не перевищує 10 хвилин із моменту адсорбції. Кількість утворених вакуолей в одну хвилину досягає більш 2 тис. Таким чином, рецепторний ендоцитоз являє собою добре злагоджений механізм, що забезпечує швидке проникнення в клітину чужорідних речовин.
Покриті вакуолі зливаються з іншими більш великими цитоплазматичними вакуолями, утворюючи рецептосоми, які містять рецептори, але без клатрину, а ті, у свою чергу, зливаються з лізосомами. Таким шляхом, прониклі в клітину білки звичайно транспортуються в лізосоми, де відбувається їх розпад на амінокислоти. Протилежністю рецепторного ендоцитозу є рідинний ендоцитоз, коли інвагінація відбувається не в спеціалізованих ділянках мембрани.
Більшість оболонкових і вірусів без оболонки проникає в клітину по механізму рецепторного ендоцитозу. Ендоцитоз забезпечує внутрішньоклітинний транспорт вірусної частки в складі ендоцитарної вакуолі, оскільки вакуоль може рухатися в будь-якому напрямку і зливатися з клітинними мембранами (включаючи ядерну мембрану), звільняючи вірусну частку у відповідних внутрішньоклітинних ділянках. Таким шляхом, наприклад, ядерні віруси попадають у ядро, а реовіруси - в лізосоми. Однак вірусні частки, що проникнули в клітину, знаходяться в складі вакуолі і відділені від цитоплазми її стінками. Їм ще необхідно пройти ряд етапів, перш ніж вони зможуть викликати інфекційний процес в клітині.
Злиття вірусної і клітинної мембран.
Для того щоб внутрішній компонент вірусу міг пройти через клітинну мембрану, ряд оболонкових вірусів еволюційно придбав механізм індукції злиття мембран. В оболонкових вірусів злиття обумовлене взаємодією вірусного білка злиття з ліпідами клітинної мембрани, у результаті якого вірусна ліпопротеїдна оболонка інтегрує з клітинною мембраною, а внутрішній компонент вірусу виявляється по іншу її сторону. У вірусів позбавлених оболонки один з поверхневих білків також взаємодіє з ліпідами клітинних мембран, у результаті чого внутрішній компонент проходить через мембрану.
Якщо при ендоцитозі вірусна частка є пасивним компонентом, то при злитті вона стає активним учасником процесу. Білком злиття є один з її поверхневих білків. Цей білок ідентифікований лише у вірусів, що мають суперкапсидну оболонку. В ортоміксо-, параміксо-, рабдо-, бунья-, корона-, тогавірусів (альфа-віруси) відомий конкретний білок, відповідальний за цю функцію. У параміксовірусів цей білок (F-білок) являє собою один із двох глікопротеїдів, що знаходяться на поверхні вірусної частки. Функцію білка злиття у вірусу грипу виконує мала субодиниця гемаглютиніну (НА2).
Параміксовіруси викликають злиття мембран при нейтральному рН, і внутрішній компонент цих вірусів може проникати в клітину безпосередньо
30