

Конспект_лекцій_Заг.вірусологія_0
.pdfЗі смертю клітини припиняється і репродукція вірусу, однак віруси, що не володіють цитопатогенною дією, можуть репродукуватися без загибелі клітини, тобто ЦПД - не обов'язковий результат продуктивної взаємодії вірусу і клітини.
Таким чином, варіанти відповідей клітини-хазяїна на вірусну інфекцію різноманітні. Для деяких систем вірус— клітина характерні ЦПД і загибель заражених клітин. В інших системах клітини ніяк зовні не реагують на інфекцію: вони зберігають морфологію, біохімічну активність і здатність ділитися. Між тим і такий стан супроводжується продукцією інфекційного потомства чи його окремих компонентів. У деяких випадках зараження параміксовірусами призводить до посилення проліферації клітин.
Хронічна (латентна) інфекція на рівні клітини. Це своєрідний стан рівноваги між вірусом і клітиною, коли не вдається знайти будь яких зовнішніх проявів інфекції. Хронічна інфекція в культурах клітин успішно відтворюється вірусами Сендай, ньюкасльської хвороби, краснухи, кору, грипу і деяких інших.
ЛЕКЦІЯ 12. ВІРУСИ БАКТЕРІЙ
Загальна характеристика бактеріофагів Бактеріофаги з ДНК-геномом Бактеріофаги з РНК геномом
Загальна характеристика бактеріофагів
Бактеріофаги (фаги) - різноманітно побудовані віруси - внутрішньоклітинні паразити прокаріотів.
Бактеріофаги - одна з найкраще вивчених груп вірусів. Прогрес у бактеріофагії зумовлений насамперед тим, що фаги виявилися дуже зручними моделями для генетичних досліджень, а також тим, що зусилля багатьох дослідників були сконцентровані на вивченні відносно невеликої кількості фагів (фаг лямбда, Т-парні фаги, дрібні РНК-фаги).
Саме вони послужили моделями для розроблення принципів молекулярної генетики. Дослідження багатьох модельних фагів досягли дуже високого рівня. Для деяких з них повністю розшифрована нуклеотидна послідовність генома, побудовані генетичні карти, виявлені промотори, оператори, термінатори тощо.
Бактеріофаги широко застосовують при розробленні методів генетичного аналізу промислових бактеріальних продуцентів, їх використовують як вектори для клонування фрагментів ДНК, введення певних генів у промислові штами бактерій.
Бактеріофаги можуть становити промисловий інтерес. Це стосується передусім штамів, у яких утворення токсинів пов’язане з наявністю відповідних профагів. Деякі бактеріофаги можуть бути джерелами виділення чистої і гомогенної за складом ДНК (фаг φ Kz Pseudomonas aeruginosa). ДНК низки бактеріофагів, що випускається на комерційній основі,
51
використовують у молекулярній біології як стандарт молекулярної маси. Бактеріофаги використовують у терапії раневих, опікових і деяких кишечних інфекцій. Препарати з цих фагів також одержують у промислових масштабах. У практику біотехнології ввійшло виробництво низки специфічних фагових ферментів (полінуклеотидлігаза фага Т4, фаговий лізоцим ДНК-полімераза), що мають цінніші властивості, ніж бактерійні. Продовжується виділення та вивчення нових фагів, які можуть стати цінними моделями для розв’язання різноманітних проблем молекулярної біології, біотехнології та медицини.
Дослідження бактеріофагів сприяло розумінню багатьох генетичних явищ. Швидке розмноження бактеріофагів дає можливість за одну добу здійснювати схрещування у потомстві двох послідовних поколінь. Для аналогічних схрещувань на дрозофілі потрібно 3,5 тижнів, а на кукурудзі - щонайменше рік. Крім того, велика кількість фагових популяцій, які містяться в кількох мілілітрах культуральної рідини, дає змогу спостерігати дуже рідкісні генетичні явища. Малий розмір геномів багатьох фагів, порівняно з геномом бактерій, дає змогу ідентифікувати всі або більшість фагових генів і досить детально уявити собі генетичну організацію і регулювання генома. Геном фага φ Х 174 мітить лише 11 генів, фага λ - менше 60, тоді як геном E. coli нараховує декілька тисяч генів. Ці переваги зробили бактеріофаги незамінними генетичними об’єктами. Вони можуть служити моделями під час аналізу будови і роботи більш складних геномів.
Ф. Туорт і Ф. Д’Ерель першими, і незалежно один від одного, виявили трансмісивні репродукуючі агенти, здатні проходити через бактеріальні фільтри і спричиняти лізис бактерій. З моменту відкриття бактеріофагів були зроблені спроби використати їх для боротьби з патогенними бактеріями. Бактеріофаги є ідеальним об’єктом для біохіміків, генетиків і мікробіологів, котрі вивчають фундаментальні механізми життєвих процесів.
Головною ознакою дії фага на чутливі бактерії є лізис бактеріальної клітини, що супроводжується виходом в оточуюче середовище нових порцій вірусу. Культура бактерій зазвичай лізується повністю, хоча бувають випадки, коли процес не має завершення. Причиною цього може бути:
1.Припинення росту культури бактерій.
2.Наявність у культурі клітин, стійких до дії фага.
3.Лізогенія, тобто перехід культури в такий стан, за якого вона є носієм генома фага і є не чутливою до повторного зараження.
Для проведення кількісних робіт з бактеріофагами використовують три основних методи досліджень:
1.Підрахунок стерильних плям (зон) на чашках з поживним агаром, який засіяний чутливими до фага бактеріями.
2.Метод кінцевих розведень, при якому лізис рідких бактеріальних культур служить індикатором наявності фага.
3.Визначення тривалості часу, необхідного для лізису стандартної бактеріальної культури в рідкому поживному середовищі.
Слід зазначити, що лише перший із них, запропонований в 1917 р. Д’Ерелем, є загальноприйнятим.
52
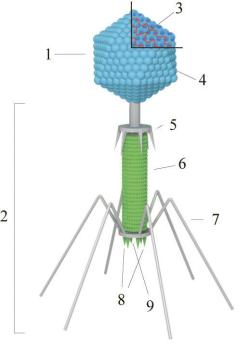
За формою, розміром і типом генома бактеріофаги побудовані порізному. Будову бактеріофагів в основному вивчали на прикладі фагів серії Т E. coli. Коліфаг Т4 характеризується змішаним типом симетрії і складається із поліедричної головки довжиною 100 нм і відростка, або "хвоста", приблизно такої ж довжини. Головка складається з капсомерів. У ній знаходиться ДНК.
Будова бактеріофага Т4: 1 – Головка, 2 – Хвіст, 3- Нуклеїнова кислота, 4 – Капсид, 5 – Комірець, 6 – Білковий чохол, 7 - Хвостові відростки, 8 –
шипи, 9 – базальна пластина.
Кількість білка і ДНК у структурі фага приблизно однакова. Хвіст фага Т4 має складну будову. В ньому можна розрізнити не менше трьох частин: пустотілий стрижень, скорочувальний чохол, який оточує стрижень і базальну пластинку з шипами і відростками на кінці стрижня. Головка з’єднана з хвостом за допомогою комірця.
Фагові частини можуть бути в двох станах: в одних частках головка різко виділяється і чохол відростка розтягнутий, у інших - чохол знаходиться в скороченому стані. Перший стан характерний для фага, в головці якого міститься ДНК, другий для фага, що здійснює ін’єкцію своєї ДНК у бактеріальну клітину.
Більшість бактеріофагів - це ДНК-вмісні віруси. Геномна ДНК різних фагів може бути однонитковою або двонитковою, лінійною або кільцевою. Прикладом вірусів з різними геномами можуть бути:
•двониткова лінійна ДНК - фаги E. coli Т4, λ, Т7;
•двониткова кільцева ДНК - фаги Alteromonas РМ2, Sulfolobus SSV 1;
•однониткова кільцева ДНК - фаги E. coli φХ174, M13.
53
Особливістю ДНК-геномів є те, що лінійні молекули ніколи не мають беззмістовних кінців. Кінці молекул можуть містити прямі або інвертовані повтори, виступаючі комплементарні (липкі) кінці.
Ще однією особливістю ДНК низки фагів є наявність у них метильованих основ (5-метилцитозину, 6-метиламінопурину), які можуть входити до складу ДНК як мінорні або мажорні основи. Так, ДНК фагів fd та φХ174 містять одну-дві метильовані основи, а у ДНК фага ХІ2, який лізує бактерії Xanthomonas oryzae, взагалі немає звичайного цитозину: він повністю заміщений 5-метилцитозином. Джерелом походження таких основ є ензиматичне метилювання вже синтезованого ланцюга ДНК. Цей процес здійснюють специфічні метилази, що використовують у ролі донора метальних груп S-аденозилметіонін.
Бактеріофаги з ДНК-геномом
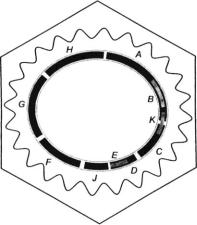
Бактеріофаг φХ174 є представником фагів з одноланцюговою ДНК. У цю групу також входять фаги М13 та G4. Вірусний геном є кільцевим і складається з 5386 нуклеотидів. Його позначають (+), тому що цей ланцюг є матрицею для синтезу комплементарного (-)-ланцюга. ДНК містить гени для синтезу 11 генних продуктів. Для генома фага φХ174 характерне явище перекривання генів, коли одна генна послідовність кодує більше, ніж один білок.
Генетична карта фага φХ174 Протягом інфікування фаг φХ174 адсорбується на ліпополісахариді
зовнішньої мембрани клітинної стінки бактерій і проникає у клітину. "Роздягання" вірусу супроводжується реплікацією вірусного генома.
Одноланцюговий ДНК-геном φХ174 після проникнення в клітину перетворюється у дволанцюгову ре-плікативну форму ДНК.
Реплікативна форма є замкненою, суперспіралізованою, дволанцюговою кільцевою ДНК. В її утворенні беруть участь РНК-праймаза, ДНК-полімераза, лігаза і гіраза - клітинно-кодуючі білки, задіяні в перетворенні ДНК фага в реплікативну форму. У фага φХ 174 реплікація починається з одного або багатьох специфічних ініціюючих сайтів в одноланцюговій кільцевій ДНК. РНК-праймаза синтезує короткий РНКпраймер для ініціації реплікації цієї ДНК. ДНК-реплікація відбувається в
54
замкненій, кільцевій ДНК, формуючи повну дволанцюгову реплікативну форму. ДНК-реплікація відбувається напівконсервативним шляхом.
Реплікативну форму ДНК використовють для синтезу фагової мРНК. Фагова мРНК починає синтезуватися на декількох промоторах на реплікативній формі і термінується декількома сайтами. Полігенна мРНК транслює різні фагові білки, включаючи А-білок, який бере участь у формуванні одноланцюгових ДНК-геномів для фагового потомства. Формування одноланцюгового фагового потомства починається з розрізання одноланцюгового фагового плюс-ланцюга реплікативної форми біля точки огі, що каталізується А-білком. А-білок ріже і лігує два кінці щойно синтезованого ланцюга для утворення кільцевої одноланцюгової ДНК. Після утворення одноланцюгової ДНК і продукування капсидного білка, новий фаг збирається. Лізис клітини-господаря відбувається тоді, коли виходить фагове покоління.
У фага φХ 174 описаний цікавий механізм лізису інфікованих клітин. Продукт гена Е цього фага (всього у нього дев’ять генів, деякі з них перекриваються) контролює включення активності бактеріальних автокаталітичних ферментів. Стимулюючий цим фагом лізис пригнічується за певних концентрацій MgS04 середовища, або якщо клітини перед інфікуванням були вирощені при pH більше 7,4. Така специфічність у реакції клітини може бути використана як чинник запобігання фаголізису на виробництві.
Бактеріофаг Mu є ДНК-матричним бактеріофагом, що може діяти як транспозон у клітині господаря. Коли геном фага Mu потрапляє у клітинугосподар (Е. coli), то нуклеази його не ріжуть, тому що ця ДНК має високе співвідношення аденіну, тобто є ацетоамідована. Інтегрування Mu-генома в бактерійну хромосому відбувається через дію транспозази, що кодується фагом. ДНК переміщується до численних сайтів хромосоми. Коли Mu-ДНК інтегрується, вона може вставлятися в структуру гена бактерійної хромосоми, що зумовлює мутацію.
Біля кожної інсерційної ділянки є п’ять пар основ дуплікації ДНК клітини-господаря. В результаті одноланцюгові області перетворюються у дволанцюгову ДНК. Продукт с-гена - репресор - попереджує про закінчення експресії Mu-геномів, що веде до утворення фагового покоління і лізису клітини-господаря. Як тільки продукується довгий С-білок, відбувається репресія,
Mu-фагова ДНК зберігається як профаг і реплікується у клітині протягом реплікації бактерійної клітини.
Ниткоподібний фаг М13 має кільцеву ДНК, що нараховує вісім генів і капсид зі спіральною симетрією. Капсид складається з 3000 молекул білків, укладених по спіралі. Вони прикріплюються до специфічного рецептора на пілях донорної клітини-господаря. Суміжні половинки генома фага підходять і зупиняються, формуючи петлі поблизу кінців. Реплікація ниткоподібного фага М13, відбувається подібно до φХ174. Вихід фага з клітини відбувається шляхом брунькування, спочатку фаг вставляється в цитоплазматичну
55
мембрану клітини-господаря. Зібраний фаг виходить з цитоплазматичної мембрани. Фаг М13 інфікує клітину, ріст якої сповільнений. Інфіковані клітини можуть продовжувати рости, поки утворються нові фагові частки.
Фаг Р1 є матричним бактеріофагом. Його ДНК не інтегрується в бактерійну хромосому під час інфікування клітини-господаря (Е. соlі). Геном Р1 фага трапляється в стані профага, подібно до плазміди в цитоплазмі. Тільки одна копія профага підтримується в клітині-господарі, тому репресор фагових генів перекриває реплікацію фагового генома з реплікацією ДНК клітини-господаря.
Віруси, що інфікують клітини нитчастих ціанобактерій Суапорhyta, мають геном, який складається з дволанцюгової ДНК; за структурою і складом часток цей вірус нагадує великі ДНК-вмісні фаги, які мають відростки. Інтерес до вивчення цих вірусів був викликаний можливістю використання їх у боротьбі з "цвітінням" водойм, яке спричиняють ціанобактерії і забруднюють прісноводні озера. Подібність між вірусами бактерій і ціанобактерій засвідчує тісний взаємозв’язок між цими двома групами мікроорганізмів.
Бактеріофаги з РНК геномом
У багатьох бактеріофагів геном представлений РНК. Добре вивченою є група фагів з одноланцюговою інфекційною РНК, до якої відносять РНКвмісні бактеріофаги R17, МS2, Qβ. РНК цих фагів на початку інфекції служить мРНК, тобто може безпосередньо транслюватися, утворюючи білки, частина яких необхідна для реплікації вірусної РНК.
Ці віруси мають вигляд ізометричних часток діаметром близько 25 нм. Їхній геном складається з РНК, що має молекулярну масу 106 Да. У клітинах бактерій геном фага транслюється з утворенням трьох білків: субодиниці РНК-реплікази, білка оболонки й А-білка (білок дозрівання). З двох останніх білків і РНК будуються інфекційні вірусні частки. Синтезована субодиниця РНК-реплікази об’єднується з трьома білками господаря і утворює вірусспецифічну РНК-репліказу, після чого починається транскрипція вірусної РНК та її реплікація.
Репліказа прикріплюється до З'-кінця вірусної РНК і рухається вздовж неї до 5'-кінця, утворюючи антипаралельний комплементарний ланцюг РНК (мінус РНК). Після закінчення синтезу комплементарної молекули репліказа прикріплюється до її З'-кінця і потім синтезує нову копію, в результаті чого виникає полінуклеотид, ідентичний материнській вірусній РНК ((+)- ланцюг). Завершивши синтез першого комплементарного (-)-ланцюга, РНК репліказа починає транскрибувати утворений комплементарний ланцюг РНК. Зазвичай синтезується геномна РНК.
Матрицею для синтезу нової геномної РНК, служить вільний мінусланцюг РНК, який не утворює з синтезованою РНК великої кількості водневих зв’язків.
Геном бактеріофага МS2 - це одноланцюгова РНК. Після проникнення і роздягання, РНК МS2-фага взаємодіє з рибосомами клітини господаря, де
56
транслюються 4 білки: білок дозрівання (А-білок), білок оболонки, білок лізису і РНК-репліказа. РНК-репліказа бактеріофага МS2 складається частково з фаг-кодуючого поліпептиду і частково з клітино-кодуючих поліпептидів. Поліпептиди клітини-господаря, що становлять частину вірусної реплікази, включають рибосомальний білок 81, який є частиною 30S рибосоми клітини-господаря і фактори елонгації.
Фагова РНК, яка є (+)-ланцюгом, може діяти як мРНК. Вона транскрибується фаговою РНК-полімеразою, що синтезує мінус-ланцюг РНК, використовуючи фагову РНК як матрицю. Молекули (+)РНК утворюються на (-)РНК. Нові (+)РНК ланцюги служать як мРНК для продовження синтезу фагового білка. Трансляція гена білка дозрівання відбувається тільки з утвореної форми (+)-ланцюга тоді, коли відбувається реплікативний процес. Це лімітує дозрівання потрібного білка, оскільки утворена фагова РНК складається в просторовий комплекс вторинної, третинної структури.
Найбільш доступним для трансляції білка оболонки є АУГ-стартовий сайт. Ці молекули білка оболонки збільшуються кількісно і накопичуються навколо АУГ стартового сайта РНК, зупиняючи синтез РНК-реплікази. Для повного збирання нового фага потрібно 180 молекул білка оболонки. Літичний білок потрібний для виходу фага МS2, кодується геном, який частково перекривається з геном оболонки і геном реплікази. Змінюється рамка зчитування, коли рибосома проходить через гени білка оболонки: тоді зчитується ген лізису. Трансляція заважає передчасному лізису клітини. Лізис починається, коли білок оболонки придатний для збирання фагової частки.
Бактеріофаг Qβ містить малу одноланцюгову РНК. Геном бактеріофага Qβ ((+)РНК-ланцюг) використовується як матриця для формування реплікативної двоииткової форми, що має плюс-ланцюг і комплементарний (-)РНК-ланцюг. Коли РНК-залежна РНК-полімераза використовує реплікативну форму як матрицю, продуктом є геномна (+)РНК, що йде в геном фагового покоління. Після збирання фага відбувається лізис клітини господаря, каталізований фагокодуючим літичним ферментом.
Із РНК-залежних-РНК-репліказ найкраще вивчена репліказа фага Qβ. Неочищена репліказа транскрибує РНК фага Qβ, а також полінуклеотиди. Після очищення репліказа синтезує РНК тільки за наявності специфічних факторів господаря.
Уфага Qβ білок оболонки пригнічує трансляцію гена реплікази так, що
знадходженням під час інфікування білка оболонки реплікази утворюється все менше. Таким же чином репліказа не тільки транскрибує мРНК вірусу, але і конкурує за неї з рибосомами. При додаванні реплікази до суміші вірусної мРНК і рибосом, репліказа зв’язується як з 3'-кінцем мРНК, так і з ділянкою, де рибосоми прикріплюються до гена білка оболонки. Завдяки цьому рибосоми не можуть синтезувати ні білок оболонки, ні репліказу (ген, який недосяжний для трансляції), якщо рибосоми не транслюють ген білка оболонки. Рибосоми, що раніше приєдналися до мРНК, завершують
57
трансляцію, і тільки після цього починається процес транскрипції. Бактеріофаг φ6 - це оболонковий, ікосаедричний, двонитковий РНК-
фаг, що регілікується в клітинах господаря Pseudomonas phaseolicola. Цей фаг містить три сегменти дволанцюгової РНК, кожен з яких синтезує мРНК. Фаг містить власну РНК-полімеразу, що може бути задіяна у реплікуванні генома фага, але механізм реплікації дволанцюгового РНК-генома цього фага ще не вивчений.
ЛЕКЦІЯ 13. ВІРУСИ РОСЛИН
Загальна характеристика фітовірусів Симптоматика та фізіологічні заражених рослин фітовірусами Поширення фітовірусів Віруси нижчих рослин і грибів
Загальна характеристика фітовірусів
Віруси рослин відрізняються в основному від всіх відомих рослинних патогенів не тільки в розмірах та морфології, а й в їхній простоті хімічної будови, фізичної структури, шляхах інфікування, репродукції, транслокації в господарі та симптоматиці, що вони викликають при зараженні господаря. Так як вони настільки малих розмірів, то вони не можуть бути так добре ідентифікованими та спостерігаємі методично, як інші патогени. Але все таки, клітинні включення, що утворюються при певних вірусних інфікуваннях і які являють собою вірусні частки, можна спостерігати в світловий мікроскоп.
Фітовіруси бувають різної форми та розмірів. Близько половини їх видовжені – нитковидні або паличковидні, інші – сферичні (ізометричні або поліедричні), або утворюють собою бациловидні часточки.
Деякі видовжені віруси являють собою структури в середньому 15 х 300 нм, але такі, які являють собою гнучкі, тонкі і більш довгі палички – на 10-13 нм ширші, а в довжині складають 480-2000 нм.
Наприклад, Рабдовіруси невеличкі, бациловидної форми, циліндричні, близько в 3 –5 разів довші відносно ширини (52-75 на 300-380 нм).
Більшість сферичних вірусів мають в поперечнику від 17 нм (сателітний вірус некрозу тютюну), до 60 нм (вірус раневих пухлин ).
Вірус бронзовості томатів вкритий зовнішньою мембраною і має гнучку, сферичну форму біля 100 в діаметрі.
Багато рослинних вірусів мають фрагментований геном, що складається із двух або більше нукленових фрагментів, що упаковані в один і той же білок, але по різному. Віруси складаються із двох складових – одна 195 на 25 нм, і другої – 43 на 25 нм, а вірус мозаїки люцерни складається із 4- х компонентів, що відрізняються по розміру.
Цікавий також факт про те, що деякі ізометричні віруси мають два або три різних компонентів однієї форми, але вміщуючи при цьому різні по довжині нуклеїновокислотні фрагменти.
58
Для таких мультикомпонентних вірусів всі ці ділянки нуклеїнової кислоти повинні бути в рослині, для того, щоб вірус був спроможен до свого нормального відтворення - репродукції.
Вірусна поверхня або капсид складається із різної кількості білкових складових, які розміщуються спірально на видовжених вірусах і упаковані по сторонах сферичних вірусів.
На поперечному зрізі видовжених вірусних частинок вони виглядають як порожня трубка із білковими субстанціями, формуючи при цьому оболонку для нуклеїнової кислоти, яка також спіралізована і упакована серед білкових закінчень капсомерів, що формують капсид.
Що стосується сферичних вірусів, то на зрізі також спостерігається утворення із білкових субодиниць навколо нуклеїнової кислоти, яка в свою чергу упакована невідомим манером.
У випадку Рабдовірусів і деяких сферичних відмічається про структурну належність ліпопротеїдного капсиду – або суперкапсиду. Всередені суперкапсиду вміщується нуклеокапсид, що складається із нуклеїнової кислоти і білкових субодиниць.
Кожен фітовірус складається як мінімум із нуклеїнової кислоти і білку. Деякі віруси складаються більш ніж із одної форми нуклеїнової кислоти та білків, деякі вміщують ліпіди та мембранні ліпіди.
Нуклеїнова кислота займає від 5 до 40% вірусу, білок – 60-95%. Найменьшу кількість нуклеїнової кислоти знайдено у паличковидних, видовжених вірусах, а найбільшу кількість нуклеїнової кислоти мають сферичні віруси.
Загальна маса нуклеопротеїну вірусних частинок варіюєж від 4,6 до 73 млн.Да, а маса очищенної НК складає в середньому 1-3 млн.Да. на вірусну частку для більшості вірусів, але деякі мають і 6 млн.Да, а 12-ти компонентна нуклеїнова кислота одного вірусу, що вражує тютюн (wound tumor virus) складає 16 млн.Да.
Цікаво те, що всі нуклеїнові кислоти вірусів в нативному вигляді, досить невеликих розмірів, тому, що вони специфічно упаковані.
Вірусні білки, як і всі білки, складаються із амінокислот, яких також відомо 20, як і для інших істот. Сіквенс амінокислот вірусного білку залежить від послідовності нуклеотидів в генетичному матеріалі і установлює природу даного білку.
Так, вірусний капсид фітовірусів складається із капсомерів, що повторюються. Вміст амінокислот та сіквенс капсомерів одного і того ж вірусу постійний, але відмінний для інших вірусів, навіть до іншого штаму одного і того ж вірусу.
Також слід відмітити, що амінокислотний склад і сіквенс різних вірусних білків однієї вірусної частинки буде різним, не говорячи про другі вірусні частинки.
Склад та сиквенс амінокислот вже відомий для багатьох вірусних білків. Наприклад, білкова субодиниця ВТМ складається із 158 амінокислот із молекулярною масою – 17,6 кДа.
59
В білку ВТМ білкові субодиниці розміщуються у вигляді спіралі, вміщуючи 16 та 1/3 субодиниць на один оберт (або 49 субодиниць на 3 оберти). Внутрішній канал ВТМ складає 4 нм, а загальний діаметр вірусної частинки складє 18 нм. Кожна вірусна частинка вміщує біля 130 обертів спіралі із білковими субодиницями. Нуклеїнова кислота в свій час упакована ретельно між біками.
Урабдовірусів спіральний нуклеопротеїн вкритий мембраною.
Уполіедричних фітовірусів білкові субодиниці упаковані ретельно в угрупування, що складаються із 20 чи іноді більше фасеток, формуючи капсид, що вкриває нуклеїнову кислоту.
Якщо розглянути в порівнянні кількість білків, що притаманні для різних організмів та форм існування, то виявляються дуже цікаві факти.
Майже всі віруси кодують білок чи білки необхідні для реплікації вірусного геному. В деяких вірусах присутні також і ферменти. Один чи декілька вірус-кодуємих білків складають капсид, що вкриває геномну нуклеїнову кислоту. Але все ж таки віруси спроможні кодувати білки із іншими функціями, наприклад:
-Рух чи транспорт – спроможність вірусного транспорту від клітини до клітини.
-Передача – можливість передачі безхребетними чи грибними векторами.
-Білковий процессінг – активність протеази, що розділяє поліпротеїн на функціональні продукти.
Майже всі віруси рослин включають до свого геному рибонуклеїнову кислоту, але близько 80 вірусів включають ДНК.
Обидві нуклеїнові кислоти довгі, спіралізовані і вміщують сотні, а частіше тисячі своїх субодиниць – нуклеотидів. Кожен нуклеотид складається із кільцевої структури, що називається основою, азотиста основа в свій час зв’язана із п’ятивуглецевим цукром – рибозою в РНК, та дезоксирибозою в ДНК, який в свою чергу зв’язаний із фосфорною кислотою.
Цукровий залишок одного нуклеотиду реагує із фосфатом другого нуклеотиду, і так повторюється багато разів, формуючи ланцюг РНК або ДНК.
Так, в молекулі РНК вірусів тільки одна із чотирьох основ – аденін, гуанін, цитозин та урацил можуть контактувати із кожною молекулою рибози. Перші два – А та Г – це пуріни, а інші – У та Ц – піримідини. Хімічні формули цих основ, що представлені в молекулі РНК представляють собою слідучі структури.
Структура ДНК подібна до РНК, але є дві маленькі відмінності, які дуже важливі – одна молекула кисню відсутня в гідроксильному радикалі цукру, та друге – урацил переструкторований в метилурацил, краще відомий як тимін – Т.
60