

total
.pdfФедеральное агентство по образованию Государственное образовательное учреждение высшего профессионального образования ПЕТРОЗАВОДСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ
Л. Е. Елаева, Г. И. Кузьмина
ФИЗИОЛОГИЯ ВОЗБУДИМЫХ СИСТЕМ И ВНУТРЕННИХ ОРГАНОВ
Учебное пособие для студентов медицинского, эколого-биологического и агротехнического факультетов
Петрозаводск Издательство ПетрГУ
2007
0 |
1 |
УДК 612.1/.8 ББК 28.9
Е 474
Рецензенты:
А. И. Шевченко, профессор, доктор медицинских наук; А. И. Горанский, профессор, доктор медицинских наук
ISBN 978-5-8021-0719-5 |
© Елаева Л. Е., Кузьмина Г. И., 2007 |
2
Часть I. ОБЩАЯ ФИЗИОЛОГИЯ ВОЗБУДИМЫХ СИСТЕМ
Все живые клетки и ткани способны реагировать на различного рода воздействия, т. е. изменять под их влиянием свою текущую функциональную активность.
Любые изменения внешней или внутренней среды достаточные, чтобы вызвать ответ со стороны клетки, получили название раздражители. Раздражители различаются по своей природе, это могут быть:
•физические факторы (свет, звук, температура и др.);
•химические (органические и неорганические вещества);
•физико-химические (осмотическое давление, рН);
•биологические (потенциал действия).
Способность клеток или тканей отвечать на действие раздражителей называется реактивностью. По характеру ответа на действие раздражителей все ткани делятся на специфические и неспецифические. Специфические ткани на действие раздражителя реагируют процессом возбуждения. Процесс возбуждения — это быстрое изменение заряда мембраны клетки в ответ на действие раздражителя. К возбудимым тканям относятся нервная, мышечная, железистая.
Глава 1 КЛЕТОЧНАЯ МЕМБРАНА
И МЕМБРАННЫЕ ПОТЕНЦИАЛЫ
Клеточная мембрана (плазмолемма) представлена двойным слоем фосфолипидов, в котором расположены молекулы белков. Фосфолипиды и белки могут быть связаны с углеводами, образуя гликопротеины и гликолипиды. Белки мембраны выполняют функцию каналов, переносчиков, ферментов, регуляторов, рецепторов; углеводы — рецепторов.
Одной из функций мембраны является транспорт веществ. Жирорастворимые вещества транспортируются, растворяясь в фосфолипидах мембраны. Водорастворимые вещества перемещаются с помощью белков. Если транспорт веществ осуществляется с затратой энергии АТФ, то он называется активным, а если без затраты АТФ — то пассивным. Транспорт одного вещества в одном направлении через каналы или не-
3
посредственно через фосфолипиды мембраны называется простой диффузией (унипорт). Транспорт веществ с участием белков-переносчиков называется облегченной диффузией. Транспорт двух веществ в противоположном направлении с помощью белков-переносчиков называется антипорт, в одном направлении — симпорт. Таким образом, транспорт водорастворимых веществ осуществляется либо с помощью белковпереносчиков, либо через каналы.
Каналы мембраны обладают способностью пропускать исключительно ионы и (или) воду. Ионные каналы представлены интегральными белками (гликопротеинами), пронизывающими липидный бислой мембраны. Систематизация и классификация каналов достаточно трудна. В настоящее время существует несколько классификаций каналов, которые в разных соотношениях учитывают свойства каналов, а именно селективность, способ открытия, скорость активации и инактивации, наличие и расположение рецепторов и регуляторных участков, чувствительность к химическим веществам (блокаторам, т. е. антагонистам, и активаторам, т. е. агонистам). Условно каналы делят на неспецифические (неселективные) и специфические (селективные). Существуют неспецифические каналы для ионов калия, натрия и кальция. Специфическиеканалыпропускаютодинвидионов.
Функционально целесообразной является классификация каналов на неуправляемые (нерегулируемые) и управляемые (регулируемые). Представителями первого вида являются калиевые каналы утечки. В покое, при исходном значении мембранного потенциала покоя, они открыты. Токи утечки участвуют в формировании мембранного потенциала покоя. Важнымсвойством регулируемых каналовявляетсяихспособностьнаходиться в открытом и закрытом состоянии. В первом состоянии они способны пропускать определенные ионы по их электрохимическому градиенту, во втором — нет. Выделяют два вида каналов в зависимости от природы управляющегофактора.
1. К первому виду относятся каналы, имеющие собственный сенсор внешнего сигнала. Он входит в состав макромолекулы канала. В зависимости от способа управления эти каналы делятся на:
а) потенциал-управляемые (электровозбудимые) — их проводимость зависит от электрического заряда на мембране (Nа-, К-, Са-, Сl-ные). Для открытия этих каналов необходимы значительные сдвиги потенциала мембраны (быстрые электровозбудимые Nа- и К-каналы, медленные электровозбудимые Са-Nа-каналы). Данный вид каналов участвует в формировании потенциала действия;
4
б) лиганд-управляемые (хемовозбудимые) — открываются и закрываются при воздействии специфических химических веществ (медиаторов). Обеспечивают быструю передачу сигналов в химических синапсах. Эти каналы открываются при связывании с рецептором специфических агонистов — ацетилхолина, глутамата, глицина, гамма-амино- масляной кислоты и называются ионотропными. Примером являются каналы постсинаптической мембраны мионеврального синапса, вегетативных ганглиев;
в) механосенситивные или стреч-каналы — чувствительны к растяжению мембраны. Эти каналы обнаружены как в возбудимых клетках (слуховых клетках, механорецепторах, нейросекреторных клетках, гладкомышечных, кардиомиоцитах), так и невозбудимых (глиальных, сосудистом эндотелии, эритроцитах). Механизм влияния растяжения на вероятность открытия стреч-каналов еще неясен. Во многих клетках были обнаружены как неселективные, так и селективные (для ионов К
иСl) стреч-каналы.
2.В каналах второго вида рецептор внешнего сигнала пространственно разобщен с каналом. Взаимодействие рецептора и канала осуществляется с помощью растворимых внутриклеточных вторичных посредников. Эти каналы называются метаботропные. При взаимодействии медиатора с рецептором на мембране активируется G-белок, который, в свою очередь, активирует фермент (аденилатциклазу, фосфолипазу). В результате в цитоплазме образуются сигнальные молекулы вторичных посредников, которые влияют на проводимость канала.
Мембранные потенциалы
Все клеточные мембраны поляризованы, т. е. регистрируется разность потенциалов между наружной и внутренней стороной мембраны. Эта разность потенциалов получила название мембранный потенциал
(МП), а в возбудимых тканях — мембранный потенциал покоя (МПП).
Величина МПП возбудимых клеток составляет от –50 мВ до –90 мВ. Механизм формирования МПП объясняет мембранно-ионная тео-
рия. Мембрана клеток возбудимых систем характеризуется наличием градиента катионов и анионов. Внутреннее содержимое клетки представлено преимущественно катионами калия и анионами органических кислот. Снаружи клетки находятся катионы натрия и анионы хлора. В состоянии покоя клеточная мембрана хорошо проницаема для ионов калия (в скелетных мышечных волокнах и для ионов хлора), менее про-
5
ницаема для ионов натрия и практически непроницаема для анионов органических кислот. Ионы калия по градиенту концентрации путем простой диффузии диффундируют из клетки через калиевые каналы утечки, заряжая наружную поверхность мембраны электроположительно. Анионы органических кислот остаются в клетке, тем самым обеспечивая появление разности потенциалов. Таким образом, внутренняя поверхность мембраны становится электроотрицательной по отношению к наружной ее поверхности. Эта разность потенциалов называется мембранным потенциалом покоя. Возникшая разность потенциалов препятствует дальнейшему выходу ионов калия из клетки, наступает равновесие между диффузией калия из клетки по концентрационному градиенту и входом этих катионов по электрическому градиенту. Мембранный потенциал, при котором достигается это равновесие, называется равновесным калиевым потенциалом. При изменении градиента ионов калия на мембране происходит изменение МПП. Функциональное значение МПП заключается в готовности клетки принимать информацию, т. е. реагировать на воздействия.
В состоянии покоя мембрана нервных клеток незначительно проницаема для ионов натрия и хлора, и вклад этих ионов в формирование МПП в нейронах невелик. Величина МПП нейронов в среднем составляет –70 мВ. Мембрана скелетных мышечных волокон в состоянии покоя относительно хорошо проницаема для ионов хлора, что приводит к увеличению трансмембранной разности потенциалов до –90 мВ. Снижение МПП носит название деполяризация, а его увеличение —
гиперполяризация.
Постоянство концентрации ионов калия и натрия на мембране обеспечивается работой натрий-калиевого насоса (натрий-калиевой АТФазы). Это интегральный белок, выполняющий функцию транспорта ионов калия в клетку и натрия из клетки. Используя одну молекулу АТФ насос выносит из клетки 3 иона натрия и вносит 2 иона калия. Таким образом, при работе насоса формируется дополнительный заряд мембраны — насосный потенциал. Следовательно, МПП нейрона имеет две составляющих: концентрационный потенциал и насосный потенциал, а натрий-калиевый насос обладает электрогенными свойствами, т. е. при работе формирует дополнительный заряд на мембране. Работа насоса блокируется сердечным гликозидом оуабаином. При блокаде насоса со временем происходит выравнивание концентраций натрия и калия на мембране и, как следствие, снижение МПП. Активация работы насоса происходит при увеличении концентрации натрия внутри клетки
6
и (или) увеличении концентрации калия на наружной поверхности мембраны.
При действии на клетку раздражителя наблюдается быстрое изменение заряда мембраны, снижение ММП и даже изменение заряда на противоположный. Этот процесс получил название процесс возбуждения. Возбуждение может протекать в двух формах: местное возбуждение
(локальный ответ, ЛО) и распространяющееся возбуждение (потенциал действия, ПД).
В зависимости от силы раздражители делятся на пороговые, подпороговые и сверхпороговые. Порог раздражения — это минимальная сила раздражителя, которая вызывает ответ клетки в форме ПД. Подпороговые раздражители вызывают местное возбуждение. Раздражители пороговой силы снижают МПП до некоторой величины, которая назы-
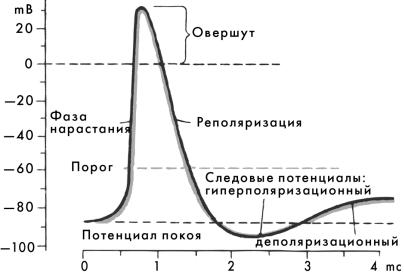
вается критическим уровнем деполяризации (КУД). При достижении КУД на мембране открываются потенциал-управляемые натриевые и калиевые каналы. Проницаемость мембраны для ионов натрия резко повышается, натрий лавинообразно устремляется внутрь клетки, что вызывает перезарядку мембраны, внутренняя сторона которой становится электроположительной по отношению к наружной. Эта фаза ПД называется быстрая деполяризация. При достижении пика ПД проницаемость мембраны для ионов натрия резко падает, а проницаемость для ионов калия становится максимальной. Ионы калия выходят из клетки через потенциал-управляемые каналы, в результате чего происходит восстановление МПП. Эта фаза называется быстрая реполяризация. Амплитуда ПД составляет 100—120 мВ, длительность 2—5 мс. Электровозбудимые натриевые каналы имеют два типа ворот — активационные (m) и инактивационные (h) — и могут находиться в трех состояниях:
—закрыты, готовы к работе (при МПП);
—открыты, т. е. активированы (фаза быстрой деполяризации ПД);
—инактивированы (пик ПД и начало фазы реполяризации). Электровозбудимые калиевые каналы имеют только один тип ворот
(активационные) и поэтому могут находиться в двух состояниях:
—закрыты, готовы к работе (при МПП);
—открыты, т. е. активированы (фаза де- и реполяризации). ПД может сопровождаться следовыми процессами:
—отрицательный следовой потенциал (следовая деполяризация);
—положительныйследовойпотенциал(следоваягиперполяризация).
7
Величина порога раздражения определяется соотношением МПП и КУД мембраны: порог = МПП – КУД.
Порог является мерой возбудимости мембраны. Увеличение порога раздражения свидетельствует о понижении возбудимости, понижение порога — о повышении возбудимости мембраны.
При действии на мембрану подпороговых раздражителей деполяризация не достигает критического уровня и ПД не возникает, формируется местное возбуждение. Местное возбуждение обладает следующими свойствами:
1)амплитуда зависит от силы раздражителя;
2)распространяется по мембране с затуханием, т. е. с уменьшением амплитуды;
3)способно к суммации и может перерасти в ПД, если деполяризация достигнет КУД;
4)возбудимость мембраны повышается, т. к. МПП приближается
кКУД.
Свойства ПД:
1)возникает при действии пороговых и сверхпороговых раздражителей. Амплитуда не зависит от силы раздражителя. Всегда максимален по величине, т. е. подчиняется закону «Все или ничего»;
2)распространяется по мембране без изменения амплитуды.
Во время развития ПД наблюдается изменение возбудимости мембраны:
а) фаза быстрой деполяризации — возбудимость резко падает. Развивается состояние полной невозбудимости (абсолютный рефрактерный период);
б) фаза быстрой реполяризации — возбудимость постепенно восстанавливается (относительный рефрактерный период);
в) следовая деполяризация — возбудимость повышена; г) следовая гиперполяризация — возбудимость понижена.
Состояние рефрактерности мембраны обусловлено полной или частичной инактивацией электровозбудимых натриевых каналов.
Значение рефрактерного периода заключается в обеспечении дискретности ПД. Рефрактерность в свою очередь определяет лабильность возбудимой ткани. Лабильность — это способность возбудимых систем воспроизводить ритм раздражения в свойственных им реакциях. Длительность рефрактерного периода определяет лабильность ткани: чем длиннее рефрактерный период, тем меньше лабильность. Мерой лабильности является максимальная частота элементарных актов воз-
8
буждения, которые может воспроизводить возбудимая система в строгом соответствии с ритмом стимуляции. Высокой лабильностью обладают нервные волокна от 500 до 1000 имп/с, меньшей — мышечные — до 250 имп/с, самой низкой лабильностью характеризуется синапс —
100 имп/с.
Функционирование Nа- и К-электровозбудимых каналов обеспечивает процесс возбуждения в клетках, поэтому большой интерес представляют вещества, изменяющие активность каналов. Часть этих веществ имеют естественное происхождение (природные нейротоксины), часть — синтетические препараты. К природным блокаторам потенци- ал-управляемых натриевых каналов относится тетродотоксин — алкалоид животного происхождения, выделяемый из рыбы фуку. Этот яд действует снаружи, формируя пробку в канале. По-иному действует батрахотоксин, секрет кожных желез колумбийской лягушки. Он блокирует быструю инактивацию канала, увеличивая период его открытия, что приводит к стойкой деполяризации мембраны. Подобным образом действует растительный алкалоид аконитин. Местные анестетики (новокаин, лидокаин, бензокаин, тетрокаин), которые по химической природе являются третичными аминами, препятствуют генерации ПД в нервных волокнах, встраиваясь между m- и h-воротами. Электровозбудимые калиевые каналы блокирует тетраэтиламмоний.
Законы раздражения
По биологическому значению все раздражители делятся на адекватные и неадекватные. Адекватными называются раздражители, к восприятию которых данная возбудимая структура приспособилась в процессе эволюции. К неадекватным относятся раздражители, не являющиеся в естественных условиях средством возбуждения, однако способные при большой силе вызвать процесс возбуждения. В качестве адекватного раздражителя возбудимых структур в физиологии и медицине используют электрический ток. Для раздражения (стимуляции) нервов или мышц используют приложенный извне электрический ток, во время протекания которого возникает процесс возбуждения.
Полярный закон раздражения
Деполяризующее действие на мембрану оказывает ток выходящего направления. Ток входящего направления гиперполяризует мембрану.
9
Закон порога
ПД возникает при действии на мембрану пороговых и сверхпороговых раздражителей. При действии подпороговых раздражителей возникает местное возбуждение (см. выше). На основании закона порога возбудимые структуры делятся на высоковозбудимые (низкопороговые) и низковозбудимые (высокопороговые).
Закон аккомодации
Величина порогового стимула зависит от скорости его нарастания. При медленном нарастании деполяризующего тока наблюдается повышение порога, т. е. аккомодация. При некоторой минимальной крутизне нарастания тока распространяющееся возбуждение вообще не возникает, а возникает лишь местное возбуждение. Причиной аккомодации является инактивация электровозбудимых натриевых каналов под влиянием длительной деполяризации мембраны и сдвиг КУД в позитивную сторону.
Фазы потенциала действия; временной ход потенциала действия в нерве
Закон гипербол (кривая силы — длительности)
Пороговая сила деполяризующего тока находится в обратной зависимости от длительности его действия, т. е. чем больше длительность стимула, тем меньше его пороговая величина.
Графическим выражением закона является гипербола. Минимальная сила прямоугольного стимула, способного вызвать процесс возбуждения, называется реобазой. Время действия стимула, равного одной реобазе, достаточное, чтобы вызвать процесс возбуждения, называется полезным временем. Время действия стимула, равного удвоенной реобазе, называется хронаксией. Данная кривая показывает, что при действии на мембрану коротких раздражителей, как бы велики они ни были, процесс возбуждения не возникает. С другой стороны, слабые раздражители, как бы долго они ни действовали, не способны вызвать ПД.
Таким образом, кривая сила — длительность, построенная для ка- кой-либо ткани, характеризует ее возбудимость. Сдвиг кривой вверх и вправо указывает на понижение возбудимости, вниз и влево — на повышение.
Механизм распространения возбуждения
Распространение возбуждения по мембранам нервных и мышечных волокон осуществляется с помощью местных токов. Между возбужденным и невозбужденным участком возникает разность потенциалов, что и создает условия для протекания локального тока. Локальный ток течет от возбужденного участка к невозбужденному. В невозбужденном участке он имеет выходящее направление и электротонически деполяризует мембрану, что вызывает в ней процесс возбуждения. Таким образом, раздражителем для невозбужденного участка мембраны волокна является ПД, возникающий на возбужденном участке, а для развития ПД имеет значение величина локального тока, достаточная, чтобы вызвать деполяризацию мембраны до КУД.
Отношение амплитуды ПД к порогу деполяризации мембраны называется гарантийным фактором (или фактором надежности). Как правило, фактор надежности равняется 5—7, что обеспечивает эффективное распространение возбуждения даже при некоторых изменениях возбудимости мембраны. При снижении гарантийного фактора до единицы ПД не распространяется.
10 |
11 |
Глава 2 НЕРВНО-МЫШЕЧНЫЙ СИНАПС
ÈСКЕЛЕТНОЕ МЫШЕЧНОЕ ВОЛОКНО
Вусловиях целого организма возбуждение переходит с одной структуры на другую, с нервной клетки на нервную, с нервной на мышечную или на железистую. Структурное образование, с помощью которого происходит передача возбуждения с одной возбудимой структуры на другую, называется синапсом. В зависимости от локализации синапсы принято делить на центральные, образованные нейронами ЦНС, и периферические, образованные нейронами и эффекторными клетками.
Аксоны мотонейронов образуют синапсы с волокнами скелетных мышц. Благодаря своей форме они называются нервно-мышечными концевыми пластинками. Они имеют все типичные морфологические характеристики химических синапсов и состоят из трех частей:
1) пресинаптической мембраны нервного окончания;
2) синаптической щели (10—50 нм);
3) постсинаптической мембраны (субсинаптической), которая образует складки, увеличивающие площадь ее поверхности.
Пресинаптическое окончание содержит митохондрии и синаптические пузырьки диаметром около 50 нм. В пузырьках находится медиатор — ацетилхолин (Ах), который выделяется в синаптическую щель при возбуждении. Количество медиатора в одном пузырьке называется квантом и составляет примерно 10 000 молекул Ах. Количество пузырьков, выделяющихся в процессе возбуждения, называется квантовое число и достигает в разных синапсах от 200 до 2000. На постсинаптической мембране находятся специфические макромолекулы, которые называются холинорецепторами. По химической природе они являются липопротеинами. Холинорецепторы связаны с хемовозбудимыми на- трий-калиевыми каналами мембраны. В области синаптической щели находится фермент ацетилхолинэстераза, который расщепляет Ах на холин и уксусную кислоту.
Механизм проведения возбуждения через синапс
ПД, распространяющийся по нервному волокну, достигает пресинаптической мембраны и вызывает ее деполяризацию, что приводит к повышению проницаемости мембраны для ионов кальция. Вход кальция в пресинаптическое окончание вызывает движение везикул к пре-
12
синаптической мембране. Мембраны пузырьков сливаются с пресинаптической мембраной, что вызывает выделение медиатора в щель. Этот процесс называется секрецией медиатора (экзоцитоз). Медиатор диффундирует через синаптическую щель и взаимодействует с холинорецепторами. Проницаемость постсинаптической мембраны для ионов натрия и калия резко повышается, и начинается движение натрия внутрь клетки, а калия из клетки. Натриевый ток превышает калиевый, т. к. натрий двигается по электрическому и концентрационному градиентам. На постсинаптической мембране развивается местная деполяри-
зация, которая называется потенциалом концевой пластинки (ПКП)
и достигает –50 мВ.
Взаимодействие Ах с рецепторами постсинаптической мембраны продолжается очень короткое время (1—2 мс), т. к. он разрушается ферментом холинэстеразой. Значительная часть продуктов расщепления Ах поступает в пресинаптическое окончание, где вновь используется для синтеза Ах. Выход ионов калия через калиевые каналы утечки восстанавливает МПП.
Потенциал концевой пластинки обладает всеми характеристиками местного возбуждения. На постсинаптической мембране никогда не возникает ПД, т. к. отсутствуют потенциал-управляемые каналы.
Между постсинаптической и соседними участками электровозбудимой мембраны мышечного волокна возникают местные токи выходящего направления, которые деполяризуют мембрану до КУД, что приводит к генерации ПД.
Свойства синаптического проведения
1.Одностороннее проведение информации, только с пресинаптической мембраны на постсинаптическую.
2.Задержка проведения информации (синаптическая задержка).
3.Низкая лабильность.
4.Высокая чувствительность к химическим веществам, в том числе
кядам.
Блокада мионеврального синапса
1.Блокада высвобождения медиатора происходит при избытке ионов магния, под действием ботулинового токсина.
2.Блокада холинорецепторов курареподобными препаратами (тубокурарин), бунгаротоксином (яд змей). При некоторых видах миастений
13
в организме могут вырабатываться антитела к холинорецепторам постсинаптической мембраны.
3. Блокада холинэстеразы под действием прозерина, эзерина носит обратимый характер, а фосфоорганические соединения (хлорофос) и нервно-паралитические газы (зарин) необратимо блокируют холинэстеразу.
Скелетное мышечное волокно
Мышечное волокно представляет собой симпласт, образованный в результате слияния отдельных миоцитов. Это многоядерное образование. Обладает следующими функциональными характеристиками: возбудимость, рефрактерность, лабильность, проводимость, сократимость.
МПП мышечных волокон довольно высокий и составляет 80— 90 мВ, формируется двумя ионными потоками: выходом калия из клетки и входом хлора в клетку. ПД мышечного волокна имеет длительность от 3 до 5 мс. Лабильность мышечных волокон — 250 имп/с. Проведение ПД по мембране осуществляется с помощью местных токов со скоростью 5 м/c.
Для связи процессов возбуждения и сокращения в мышечном во-
локне имеется система электромеханического сопряжения. Она пред-
ставлена поперечными (Т) и продольными (L) трубочками. Т-трубочки являются впячиваниями мембраны и выполняют функцию проведения возбуждения вглубь волокна. L-трубочки — производные эндоплазматической сети, замкнутые образования, имеющие на концах расширения — цистерны, в которых депонируются в связанной с белками форме ионы кальция. Накопления кальция в цистернах ЭПС обусловлено работой энергозависимого кальциевого насоса. ПД проходит по мембране, заходит в Т-трубочки, вызывает деполяризацию L-трубочек, что приводит к повышению проницаемости для ионов кальция и выделению кальция в пространство между миофибриллами.
Механизм мышечного сокращения
Механизм мышечного сокращения описывает теория скользящих нитей. Миофибриллы мышечного волокна состоят из белковых нитей — тонких нитей актина и толстых миозина. Нити актина и миозина образуют упорядоченную структуру — саркомер. Нити актина прикрепляются к Z-мембранам, а нити миозина располагаются между ними. Участок
14
миофибриллы между двумя соседними Z-мембранами — это один саркомер. В миофибрилле находятся тысячи саркомеров. При сокращении нити актина двигаются вдоль толстых нитей миозина, а Z-мембраны навстречу друг другу, длина нитей при этом не меняется. На нитях актина через определенные промежутки находятся места прикрепления головок миозина, которые в состоянии расслабления блокируются белком тро-
помиозином.
Тропомиозин связан с комплексом белковых молекул тропонина. При взаимодействии с ионами кальция происходит конформационная перестройка субъединиц тропонина, что меняет положение тропомиозина и открывает места прикрепления для головок миозина. Головка миозина прикрепляется к нити актина под углом 90°, после чего совершает поворот на 45° — гребок, что продвигает нить актина относительно миозина. Совместные гребущие усилия тысячи головок миозина создают мышечное сокращение.
Отделение головки миозина от нити актина возможно только с затратой энергии молекулы АТФ. В присутствии ионов кальция и магния АТФ-ную активность приобретает молекула миозина, которая расщепляет АТФ и обеспечивает фосфорилирование миозина и отделение головок. Сокращение продолжается до тех пор, пока в пространстве между нитями сохраняется высокая концентрация ионов кальция. Снижение концентрации кальция и, следовательно, расслабление происходит за счет активации кальциевого насоса мембран саркоплазматического ретикулума.
Режимы мышечного сокращения
1.Одиночное сокращение. В этом режиме мышца сокращается, если интервал между импульсами больше времени сокращения и расслабления мышцы.
2.Тетаническое сокращение развивается в результате суммации одиночных сокращений. Суммация одиночных сокращений происходит вследствие сохранения высокой концентрации кальция в пространстве между нитями.
Возможны два вида суммации:
— зубчатый тетанус развивается, если следующий импульс возникает на мембране мышечного волокна во время расслабления;
— гладкий тетанус развивается, если интервал между импульсами меньше времени одиночного сокращения.
15
Виды мышечного сокращения
1.Изотоническое сокращение. При сокращении происходит уменьшение длины мышечного волокна без изменения напряжения.
2.Изометрическое сокращение. При сокращении изменяется напряжение без изменения длины.
3.Смешанное (ауксотоническое) сокращение. Происходит изменение и длины и напряжения мышцы. Чаще всего встречается в естественных условиях работы мышцы.
Двигательные единицы и их типы
Морфофункциональной основой организации мышцы является двигательная единица (ДЕ). Двигательная единица — это альфа-мотонейрон и группа мышечных волокон, которые он иннервирует. ДЕ бывают трех типов(см. Моторныесистемы).
По составу ДЕ все мышцы человека являются смешанными, т. е. включают ДЕ разных типов.
Энергетика мышцы
Единственным прямым источником энергии для мышечного сокращения является молекула АТФ. Для образования АТФ в сокращающейся мышце существуют три энергетические системы:
1.Фосфагенная или АТФ-креатинфосфатная система. Ресинтез АТФ происходит за счет высокоэнергетического соединения креатинфосфата. Обеспечивает работу мышцы в течение 5—6 секунд.
2.Гликолитическая система. АТФ образуется в результате анаэробного распада углеводов. Обеспечивает работу мышцы в течение от 20 секунд до 1—2 минут.
3.Окислительная система. Энергия образуется в результате аэробного распада углеводов и жиров. При достаточном обеспечении мышцы кислородом может обеспечивать работу умеренной мощности в течение нескольких часов.
Глава 3 ФУНКЦИОНАЛЬНЫЕ ОСОБЕННОСТИ
ГЛАДКИХ МЫЩЦ
Гладкие мышцы находятся в стенках внутренних органов, кровеносных и лимфатических сосудов, радужной оболочки глаза, ресничном теле. Они
16
образованы веретенообразными клетками, между которыми имеется особый вид плотных контактов — нексусы. Нексусы делают возможным электротоническоераспространениевозбужденияпомышце.
МПП гладких мышц — 60—70 мВ. Некоторые клетки имеют неустойчивый МПП, который спонтанно понижается, и при достижении КУД происходит генерация ПД. Такие клетки называются пейсмекерами. Они формируют очаги автоматии в гладких мышцах. Возбуждение электротонически распространяется от клетки к клетке со скоростью
2—10 см/с.
Развитие фазы деполяризации ПД связано с входом в клетку ионов кальция через медленные кальций-натриевые каналы. ПД имеет большую длительность — от 50 до –250 мс.
Гладкая мышца не имеет поперечной исчерченности, т. к. миофибриллы расположены неупорядоченно. Миофибриллы состоят из сократительных белков актина и миозина. Сокращение миофибрилл происходит в результате скольжения нитей актина вдоль нитей миозина. Из регуляторных белков присутствует только тропомиозин. Плохо развит саркоплазматический ретикулум, поэтому кальций, необходимый для электромеханического сопряжения, поступает из внеклеточной жидкости во время ПД. Скорость сокращениянизкая, ипри частоте1 имп/свозникает гладкий тетанус. Энерготраты крайне невелики, поэтому гладкие мышцы приспособлены к длительному сокращению без развития утомления. Характерным свойством гладких мышц является пластичность, т. е. они могут быть полностью расслабленыкаквукороченном, такиврастянутомсостоянии.
Регуляция тонуса и сократительной активности гладких мышц осуществляется вегетативной нервной системой (ВНС), а также гуморальными и механическими факторами. Гладкие мышцы обладают высокой чувствительностью к биологически активным веществам крови и тканевой жидкости, т. к. имеют рецепторы на всей поверхности мембраны, а не только в области синаптических контактов.
Некоторые особенности нервных влияний на гладкие мышцы связаны со строением периферических вегетативных синапсов. Аксоны постганглионарных нейронов, направляясь к гладкой мышце, образуют сеть варикозных расширений. Расширения заполнены синаптическими пузырьками и могут формировать контакты с большим количеством гладкомышечных клеток. Гладкомышечные клетки, расположенные далеко от хода нервных волокон, возбуждаются потенциалами действия соседних миоцитов, которые электротонически распространяются по мембране и передаются через нексусы.
17
Глава 4 ФИЗИОЛОГИЯ НЕЙРОНА
И ЦЕНТРАЛЬНЫХ СИНАПСОВ
Нервная клетка (нейрон, нейроцит) является структурной и функциональной единицей центральной нервной системы (ЦНС).
Части нейрона и их функции
1.Дендриты — это отростки нейрона, на которых располагаются синапсы других нейронов, выполняют функции восприятия информации и проведения к телу нейрона.
2.Сома (тело нейрона) обеспечивает суммацию потенциалов, поступающих от дендритов и возникающих в собственных синапсах (интегративная функция).
3.Аксонный холмик — это начальный сегмент аксона, наиболее возбудимая часть нейрона, имеет самый низкий порог раздражения, место генерации ПД.
4.Аксон — отросток нейрона, который проводит возбуждение от тела нейрона к другим клеткам.
5.Пресинаптическое окончание (терминаль) выполняет функцию накопления медиатора и секреции в синаптическую щель.
Классификация нейронов
По выполняемой функции различают три основных типа нейронов: а) афферентные (чувствительные). По строению являются биполярными (или псевдоуниполярными), их тела вынесены за пределы ЦНС и располагаются в чувствительных ганглиях. Воспринимают сигналы, возникающие в рецепторных образованиях органов чувств и проводят
их в ЦНС; б) эфферентные (двигательные). По строению являются мультипо-
лярными, их аксоны выходят за пределы ЦНС и обеспечивают контроль за эффекторными органами;
в) вставочные (интернейроны, ассоциативные). По строению являются мультиполярными. Выполняют интегративную функцию. Обрабатывают информацию, поступающую от первичных афферентных нейронов и от других интернейронов и передают двигательным нейронам.
По выделяемому медиатору нейроны бывают холинергические, адренергические, пуринергические, серотонинергические и т. д.
18
По расположению в ЦНС выделяют спинальные, бульбарные, корковые и т. д.
Центральные синапсы — это синапсы, образуемые нейронами в пределах ЦНС. По строению они бывают: аксосоматические, аксодендритные, аксоаксональные. По механизму проведения возбуждения: химические, электрические, смешанные. В ЦНС человека и млекопитающих преобладающими являются химические синапсы.
Возбуждающие синапсы
В данных синапсах на постсинаптическое мембране при взаимодействии медиатора с мембранными рецепторами происходит повышение проницаемости мембраны для ионов калия и натрия. В результате ионных токов развивается местная деполяризация, которая называется воз-
буждающим постсинаптическим потенциалом (ВПСП). ВПСП обла-
дает всеми свойствами местного возбуждения. Имеет небольшую амплитуду, т. е. является подпороговым, и для сдвига МПП нейрона до КУД необходима суммация ВПСП. Суммация ВПСП может происходить в двух формах: временной и пространственной. При временной суммации увеличение амплитуды ВПСП происходит за счет возрастания частоты ПД в афферентном волокне. Происходит суммация ВПСП
водном синапсе. При пространственной суммации рост амплитуды ВПСП происходит при одновременном возникновении возбуждения
вблизко расположенных синапсах.
Тормозные синапсы
Располагаются, как правило, на соме нейронов. При действии медиатора на мембранные рецепторы постсинаптической мембраны в ней открываются калиевые и (или) хлорные каналы, что вызывает гиперпо-
ляризацию, которая называется тормозным постсинаптическим по-
тенциалом (ТПСП). Возбудимость нейрона при этом понижается, т. к. МПП удаляется от КУД.
Таким образом, интегративная деятельность нейрона — это суммация ТПСП и ВПСП во времени и пространстве. При повышении активности возбуждающих синапсов нейрон увеличивает частоту ПД, а при активации тормозных синапсов понижает ее.
Принцип Дейла
1. Нейрон во всех своих синаптических контактах выделяет один вид медиатора.
19