

ліпіди
.docЛІПІДИ.
БІОМЕМБРАНИ
Ліпіди
- це органічні речовини біологічного
походження, які нерозчинні у воді і
добре розчинні в неполярних розчинниках.
Більшість з них за хімічною будовою
являються естерами багатоатомних,
високомолекулярних або специфічно
побудованих спиртів з вищими жирними
кислотами, або належать до їх похідних
чи складників.
За хімічною будовою
ліпіди поділяють на прості і складні.
Прості
ліпіди не містять інших компонентів
крім залишків спирту та вищих жирних
кислот. До них належать жири, воски та
стериди.
Складні ліпіди крім залишків
спирту та жирних кислот містять ще інші
компоненти, за хімічним складом яких
поділяються на фосфоліпіди, гліколіпіди,
сульфоліпіди.
2.1.
Жирні кислоти
Жирні
кислоти, які входять до складу ліпідів,
містять, переважно, парну кількість
атомів карбону, найчастіше - 16 або 18.
Серед них є насичені і ненасичені (Табл.
2.1.).
В більшості ненасичених жирних
кислот подвійний зв'язок розташований
між 9-м і 10-м атомами карбону. Додаткові
подвійні зв'язки розташовуються між С9
подвійним зв'язком і СН3-кінцем молекули,
причому між ними міститься метиленова
група:. -СН=СН-СН2-СН=СН. Подвійні зв'язки
знаходяться переважно в цис-конформації,
що приводить до сильного згину вуглецевого
ланцюга:
 а
– насичений ланцюг б – цис-ненасичений
ланцюг
а
– насичений ланцюг б – цис-ненасичений
ланцюг
Радикали жирних кислот з кількома подвійними зв’язками мають декілька згинів ланцюга, а тому є компактними, тоді як насичені, завдяки вільному обертанню навколо sp3-гібридизованих атомів С, - витягнуті. Ця відмінність будови проявляється у особливостях агрегатного стану. При температурі тіла насичені жирні кислоти від С12 до С24 знаходяться у твердому віскоподібному стані, а ненасичені – у рідкому. Радикали ненасичених жирних кислот хімічно активні. За місцем кратних зв’язків вони вступають в реакції приєднання, окиснення, полімеризації. Тому їх можна якісно виявити за допомогою бромної води (реакція приєднання), знебарвлення розчину перманганату калію (реакція окиснення). Для кількісного визначення ступеня ненасиченості ліпідів використовують йодометричне титрування. Ненасичені кислоти слабкими окисниками окиснюються в диоксикислоти, а при енергійному окисненні відбувається розрив -С=С- зв'язку з утворенням двох молекул карбонових кислот: В поліненасичених жирних кислотах -СН2- групи, розташовані між двома подвійними зв'язками, також проявляють високу реакційну здатність, оскільки в результаті відщеплення від них атома гідрогену утворюється стабільний аніон з делокалізованим негативним зарядом: -СН=СН-СН2-СН=СН- > -СН?СН?СН?СН?СН- Взаємодія цих СН2 груп з киснем приводить до утворення гідропероксидів: >НС-О-О-Н. Пероксиди розкладаються, при цьому відбувається полімеризація, утворюються поперечні зшивки між ланцюгами. Перекисне окиснення ліпідів (ПОЛ) протікає в біологічних системах з незначною інтенсивністю, але посилюється в присутності деяких токсичних (чужорідних) речовин, наприклад йонів важких металів, або під дією проникаючої радіації. Одним з кінцевих продуктів перекисного окиснення є малоновий диальдегід: НОС-СН2-СОН, за концентрацією якого судять про інтенсивність ПОЛ. Він та інші продукти ПОЛ мають руйнівну дію на різноманітні клітинні структури і молекули. В природних сумішах ліпідів містяться антиоксиданти, які певною мірою захищають ненасичені радикали від руйнування. До них, зокрема, належить вітамін Е, каротиноїди. Самі ненасичені жирні кислоти також проявляють антиоксидантні властивості. Вода, вуглекислота, гемоглобін також «гасять» збуджені електронні стани. Таблиця 2.1. ВИЩІ ЖИРНІ КИСЛОТИ – СКЛАДНИКИ ЛІПІДІВ
|
Число атомів карбону |
Систематична назва |
Раціональна назва |
Скорочене позна-ченняа |
Раціональна назва ацильної групи |
|
Насичені жирні кислоти |
||||
|
4 |
Бутанова |
Масляна |
4:0 |
Бутирил |
|
12 |
Додеканова |
Лауринова |
12:0 |
Лауроїл |
|
14 |
Тетрадеканова |
Міристинова |
14:0 |
Міристоїл |
|
16 |
Гексадеканова |
Пальмітинова |
16:0 |
Пальмітоїл |
|
18 |
Октадеканова |
Стеаринова |
18:0 |
Стеароїл |
|
20 |
Ейкозанова |
Арахінова |
20:0 |
|
|
22 |
Докозанова |
Бегенова |
22:0 |
|
|
24 |
Тетракозанова |
Лігноцеринова |
|
|
|
Ненасичені жирні кислотиб |
||||
|
4 |
|
Кротонова |
4:12 (2t) |
Кротоноїл |
|
16 |
|
Пальмітоолеїнова |
16:1 (9с) |
|
|
18 |
|
Олеїнова |
18:1 (9с) |
Олеоїл |
|
18 |
|
Вакценова |
18:1 (11с) |
|
|
18 |
|
Лінолева |
18:2 (9с, 12с) |
|
|
18 |
|
Ліноленова |
18:3 (9с, 12с, 15с) |
|
|
20 |
|
Арахідонова |
20:4 (5с, 8с, 11с, 14с) |
|
Примітка: а – першим йде число атомів карбону, потім число подвійних зв’язків. В скобках подається номер першого з атомів карбону при подвійному зв’язку і тип зв’язку – цис-(с) або транс-(t). б – систематичні назви в цьому випадку дуже складні і майже не використовуються; наприклад назва ліноленової кислоти – цис, цис, цис-9,12,15-октадекатрієнова кислота
На
підставі вищесказаного можна було би
вважати, що ненасичені жирні кислоти є
небажаними компонентами природних
ліпідів. Проте вони абсолютно необхідні
для забезпечення агрегатного стану
біологічних мембран. Крім того, вони,
на відміну від насичених жирних кислот,
необмежено розчиняються у неполярних
розчинниках, і тому протидіють відкладенню
холестерину на стінках кровоносних
судин та інших утворень ліпідної природи.
Поліненасичені жирні кислоти в організмі
людини не синтезуються і повинні
поступати з їжею. Їх основним джерелом
є рослинні олії. Рекомендована доза
складає для соняшникової олії біля
однієї столової ложки на добу.
Окрему
групу біологічно активних речовин
утворюють циклічні похідні ненасичених
С20 кислот – простагландини:
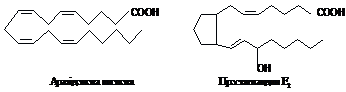 Їх
вперше вдалося одержати в кристалічному
стані в 1960 р. Зараз відома вже ціла родина
простагландинів. Представники її
відрізняються наявністю окси- або
кетогруп. Вони викликають скорочення
гладкого м'язу при концентрації всього
10-9 М, тоді як в сім'яній рідині їх вміст
становить біля 10-3 М. Надзвичайний інтерес
і перспективу представляє дослідження
їх участі в протіканні запального
процесу, алергічних реакцій.
2.2.
Прості ліпіди
Жири(тригліцериди)
- це естери вищих жирних кислот та
трьохатомного спирту гліцерину. Структура
гліцериду:
Їх
вперше вдалося одержати в кристалічному
стані в 1960 р. Зараз відома вже ціла родина
простагландинів. Представники її
відрізняються наявністю окси- або
кетогруп. Вони викликають скорочення
гладкого м'язу при концентрації всього
10-9 М, тоді як в сім'яній рідині їх вміст
становить біля 10-3 М. Надзвичайний інтерес
і перспективу представляє дослідження
їх участі в протіканні запального
процесу, алергічних реакцій.
2.2.
Прості ліпіди
Жири(тригліцериди)
- це естери вищих жирних кислот та
трьохатомного спирту гліцерину. Структура
гліцериду:
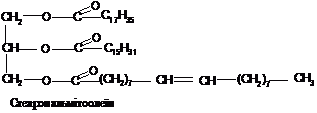 При
їх повному гідролізі в слаболужному
середовищі в організмі утворюються
гліцерин та мила (солі жирних кислот),
а при частковому -ди- та моногліцериди,
в яких естерифіковані відповідно дві
або одна гідроксильні групи в молекулі
гліцерину. Природні жири при зберіганні
піддаються гідролізу за участю
мікроорганізмів (згіркають). Неприємний
запах, який при цьому з'являється,
зумовлений утворенням низькомолекулярних
жирних кислот.
Якість жиру визначають
за його хімічними параметрами. Кількість
вільних жирних кислот в жирі
характеризується кислотним
числом жиру,
яке показує скільки міліграмів лугу
(КОН) потрібно для нейтралізації вільних
жирних кислот, що містяться в 1 г жиру.
Загальне число жирних кислот (вільних
і зв’язаних) в складі жиру
характеризують числом
омилення жиру, яке
показує, скільки міліграмів гідроксиду
калію необхідно для нейтралізації всіх
вищих жирних кислот, що містяться в 1 г
жиру. Ступінь ненасиченості жиру
визначають як масу йоду, г, яка може
прореагувати з 100 г жиру (йодне
число). Йодне
число, наприклад, волового жиру становить
- 38-46, а конопляної олії - 150.
Жири
виконують важливі функції в тваринних
та рослинних організмах. Вони відкладаються
в жировій тканині тварин - підшкірній
жировій клітковині та навколо окремих
органів і використовуються по мірі
потреби як джерело енергії. 90 % видів
рослин мають великі запаси жиру в
насінні. У соняшника вони досягають
60 % ваги насіння. Жири е найбільш
ефективним енергетичним ресурсом. При
повному окисненні 1 г жиру вивільняється
біля 39 кДж енергії, тоді як при окисненні
1 г вуглеводів або білків - 17 кДж. Вони
енергетично забезпечують довготривалу
м'язову роботу, наприклад, сезонну
міграцію птахів, забезпечують
життєдіяльність тварин під час зимової
сплячки, голодування. В нормі резервні
ліпіди становлять 10-15 % від маси тіла.
При ожирінні їх вміст збільшується до
25-35 %.
Жири захищають організм від
переохолодження. Так, у "моржів"
збільшений шар підшкірної жирової
тканини, а в насінні рослин помірного
поясу накопичуються жири у 80 % видів,
тоді як в тропіках лише у 5 % рослин.
При
повному окисненні жирів утворюється
досить багато води (107 г на 100 г
жиру). Тому, наприклад, відкладення жиру
в горбі у верблюда служить джерелом
ендогенної води.
Воски -
естери вищих спиртів і вищих жирних
кислот. В складі восків знайдено декілька
десятків кислот і спиртів, переважно
насичених. Найчастіше зустрічаються
спирти цетиловий (С16Н33ОН) і мірициловий
С30Н61ОН. У бджолиному воску, наприклад,
переважає пальмітиновомірициловйй
ефір: С15Н31СООС30Н61. Воски зустрічаються
як у тваринному, так і в рослинному
царстві, де виконують, в основному,
захисні функції. Так, в рослинах вони
покривають тонким шаром листки, стебла,
плоди, охороняючи їх від води і проникнення
мікроорганізмів. Бджолиний віск оберігає
мед, личинок бджіл. Ланолін - тваринний
віск - оберігає волосся і шкіру від дії
води. У фітопланктону воски виконують
функцію клітинного палива.
Цікавим
прикладом ролі жирів і восків у біологічній
адаптації є їх використання кашалотом.
В його голові, маса якої становить
третину маси тіла, над верхньою щелепою
знаходиться спеціальний утвір –
спермацетовий мішок, на який припадає
90 % маси голови. В ньому міститься до
4 т спермацету – суміші, багатої на
ненасичені вищих жирних кислот За
допомогою цього мішку кашалот регулює
плавучість. При температурі 370 спермацет
рідкий, а при 310 – твердий. Тому кашалот
може до 50 хв. знаходитись на великій
глибині, полюючи на кальмарів, і
підніматись на поверхню на короткий
час для поповнення запасів кисню,
підтримуючи, завдяки різному стану
спермацету, плавучість, відповідну
густині води.
При
їх повному гідролізі в слаболужному
середовищі в організмі утворюються
гліцерин та мила (солі жирних кислот),
а при частковому -ди- та моногліцериди,
в яких естерифіковані відповідно дві
або одна гідроксильні групи в молекулі
гліцерину. Природні жири при зберіганні
піддаються гідролізу за участю
мікроорганізмів (згіркають). Неприємний
запах, який при цьому з'являється,
зумовлений утворенням низькомолекулярних
жирних кислот.
Якість жиру визначають
за його хімічними параметрами. Кількість
вільних жирних кислот в жирі
характеризується кислотним
числом жиру,
яке показує скільки міліграмів лугу
(КОН) потрібно для нейтралізації вільних
жирних кислот, що містяться в 1 г жиру.
Загальне число жирних кислот (вільних
і зв’язаних) в складі жиру
характеризують числом
омилення жиру, яке
показує, скільки міліграмів гідроксиду
калію необхідно для нейтралізації всіх
вищих жирних кислот, що містяться в 1 г
жиру. Ступінь ненасиченості жиру
визначають як масу йоду, г, яка може
прореагувати з 100 г жиру (йодне
число). Йодне
число, наприклад, волового жиру становить
- 38-46, а конопляної олії - 150.
Жири
виконують важливі функції в тваринних
та рослинних організмах. Вони відкладаються
в жировій тканині тварин - підшкірній
жировій клітковині та навколо окремих
органів і використовуються по мірі
потреби як джерело енергії. 90 % видів
рослин мають великі запаси жиру в
насінні. У соняшника вони досягають
60 % ваги насіння. Жири е найбільш
ефективним енергетичним ресурсом. При
повному окисненні 1 г жиру вивільняється
біля 39 кДж енергії, тоді як при окисненні
1 г вуглеводів або білків - 17 кДж. Вони
енергетично забезпечують довготривалу
м'язову роботу, наприклад, сезонну
міграцію птахів, забезпечують
життєдіяльність тварин під час зимової
сплячки, голодування. В нормі резервні
ліпіди становлять 10-15 % від маси тіла.
При ожирінні їх вміст збільшується до
25-35 %.
Жири захищають організм від
переохолодження. Так, у "моржів"
збільшений шар підшкірної жирової
тканини, а в насінні рослин помірного
поясу накопичуються жири у 80 % видів,
тоді як в тропіках лише у 5 % рослин.
При
повному окисненні жирів утворюється
досить багато води (107 г на 100 г
жиру). Тому, наприклад, відкладення жиру
в горбі у верблюда служить джерелом
ендогенної води.
Воски -
естери вищих спиртів і вищих жирних
кислот. В складі восків знайдено декілька
десятків кислот і спиртів, переважно
насичених. Найчастіше зустрічаються
спирти цетиловий (С16Н33ОН) і мірициловий
С30Н61ОН. У бджолиному воску, наприклад,
переважає пальмітиновомірициловйй
ефір: С15Н31СООС30Н61. Воски зустрічаються
як у тваринному, так і в рослинному
царстві, де виконують, в основному,
захисні функції. Так, в рослинах вони
покривають тонким шаром листки, стебла,
плоди, охороняючи їх від води і проникнення
мікроорганізмів. Бджолиний віск оберігає
мед, личинок бджіл. Ланолін - тваринний
віск - оберігає волосся і шкіру від дії
води. У фітопланктону воски виконують
функцію клітинного палива.
Цікавим
прикладом ролі жирів і восків у біологічній
адаптації є їх використання кашалотом.
В його голові, маса якої становить
третину маси тіла, над верхньою щелепою
знаходиться спеціальний утвір –
спермацетовий мішок, на який припадає
90 % маси голови. В ньому міститься до
4 т спермацету – суміші, багатої на
ненасичені вищих жирних кислот За
допомогою цього мішку кашалот регулює
плавучість. При температурі 370 спермацет
рідкий, а при 310 – твердий. Тому кашалот
може до 50 хв. знаходитись на великій
глибині, полюючи на кальмарів, і
підніматись на поверхню на короткий
час для поповнення запасів кисню,
підтримуючи, завдяки різному стану
спермацету, плавучість, відповідну
густині води.
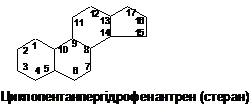 Стериди-
це естери специфічно побудованих
циклічних спиртів стеролів (стеринів)
і вищих жирних кислот В організмі
переважно поширені вільні стероли
(близько 90 %), а не їх естери. Стероли
є продуктами окиснення поліциклічного
вуглегідрогену циклопентанопергідрофенантрену.
Вони містять чотири конденсованих
кільця (це гідрофобна частина молекули)
і полярну групу – ОН.
Головним стеролом
у людини і тварин є холестерол (холестерин),
в грибах міститься ергостерол, для
рослин типовий сітостерол:
Стериди-
це естери специфічно побудованих
циклічних спиртів стеролів (стеринів)
і вищих жирних кислот В організмі
переважно поширені вільні стероли
(близько 90 %), а не їх естери. Стероли
є продуктами окиснення поліциклічного
вуглегідрогену циклопентанопергідрофенантрену.
Вони містять чотири конденсованих
кільця (це гідрофобна частина молекули)
і полярну групу – ОН.
Головним стеролом
у людини і тварин є холестерол (холестерин),
в грибах міститься ергостерол, для
рослин типовий сітостерол:
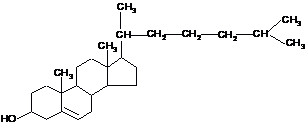
Холестерол (холестерин)
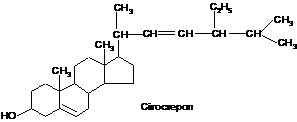
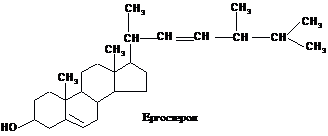
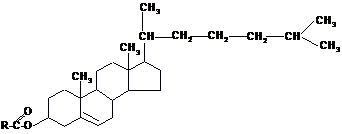 Холестерид
(естер холестеролу)
Холестерид
(естер холестеролу)
Стериди є обов’язковими структурними компонентами біомембран. Особливо багато їх у зовнішніх мембранах клітин (до 30 % всіх мембранних ліпідів). Важливу біологічну роль виконують похідні стеролів, у яких вкорочений (окиснений) бічний ланцюг. До яких належать стероїдні гормони, жовчні кислоти, вітамін Д. Серед наведених нижче прикладів кортикостерон є одним з найбільш поширених гормонів кори наднирників, естрадіол – жіночий статевий гормон, тестостерон – чоловічий статевий гормон, холева кислота – одна із жовчних кислот:
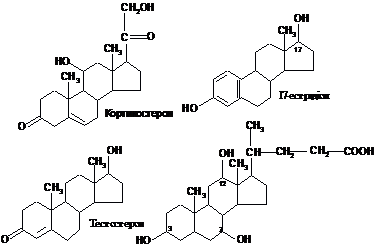 Холева
(3,7,12-тригідроксооксихоланова) кислота
Холева
(3,7,12-тригідроксооксихоланова) кислота
Всього
в організмі людини міститься біля 140 г
холестеролу. Його концентрація в крові
становить біля 2 г/л, причому третина
його знаходиться у вільному стані. За
нормальних умов він знаходиться у
твердому стані (t0пл.=1480). Одне з найбільш
поширених сучасних захворювань -
атеросклероз - пов'язане з відкладанням
стеролів та стеридів на внутрішній
поверхні кровоносних судин, що приводить
до обмеження кровотоку, а отже інсульту,
інфаркту або інших паталогій.
2.3.
Складні ліпіди
Складні
ліпіди можна розглядати як похідні
простих ліпідів, в яких залишок спирту
сполучений з полярною групою. Молекули
складних ліпідів мають загальний план
будови. Вони містять декілька (2, 4)
неполярних довгих вуглеводневих
радикалів (з яких хоча б один ненасичений)
і які, переважно, належать залишкам
вищих жирних кислот Це гідрофобні
“хвости” молекули, сполучені із залишком
спирту (який також може утворювати
неполярний радикал, як, наприклад,
сфінгозин). Полярна група (“голова”)
представлена фосфатом, азотистою
основою, вуглеводом, або його сульфопохідним.
Таким чином, складні ліпіди – це дифільні
молекули, які складаються з полярної
“головки” і декількох неполярних
хвостів:
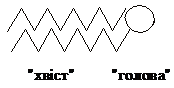 У
водному середовищі такі молекули
утворюють міцели, моношари або бішари,
в яких полярні “голови” орієнтовані
до води, а неполярні “хвости” обернені
до гідрофобної фази або повітря, уникаючи
контактів з водою:
У
водному середовищі такі молекули
утворюють міцели, моношари або бішари,
в яких полярні “голови” орієнтовані
до води, а неполярні “хвости” обернені
до гідрофобної фази або повітря, уникаючи
контактів з водою:
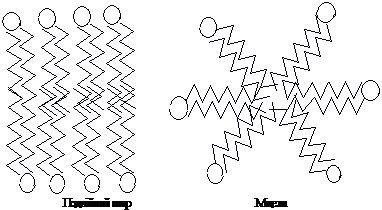 Фосфоліпіди –
найбільш поширені складні ліпіди. Їх
полярна група містить залишок ортофосфорної
кислоти і азотистої основи або
багатоатомного спирту.
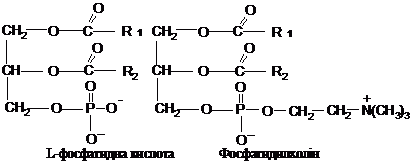
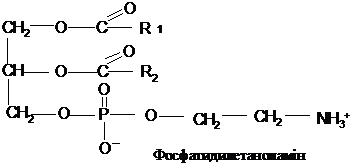
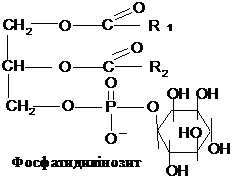
Фосфоліпіди,
залежно від природи спирту, сполученого
із залишками вищих жирних кислот,
поділяються на фосфогліцериди і
сфінгоміеліни. Фосфогліцериди складаються
із залишку гліцерину, з’єднаного по С1
і С2 із залишками вищих жирних кислот
(по С2, як правило, ненасиченої) і по С3 –
із залишком фосфорної кислоти. Утворена
сполука називається фосфатидною
кислотою. Через залишок фосфату вона
з’єднується з азотистою основою (
найчастіше холіном, етаноламіном або
серином) або циклічним спиртом
інозитолом.
Фосфоліпіди –
найбільш поширені складні ліпіди. Їх
полярна група містить залишок ортофосфорної
кислоти і азотистої основи або
багатоатомного спирту.
Фосфоліпіди,
залежно від природи спирту, сполученого
із залишками вищих жирних кислот,
поділяються на фосфогліцериди і
сфінгоміеліни. Фосфогліцериди складаються
із залишку гліцерину, з’єднаного по С1
і С2 із залишками вищих жирних кислот
(по С2, як правило, ненасиченої) і по С3 –
із залишком фосфорної кислоти. Утворена
сполука називається фосфатидною
кислотою. Через залишок фосфату вона
з’єднується з азотистою основою (
найчастіше холіном, етаноламіном або
серином) або циклічним спиртом
інозитолом.

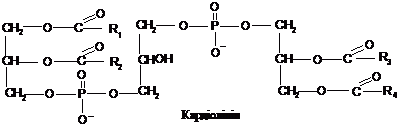
 Кардіоліпіни
є “подвійними” фосфогліцеридами. У
них два залишки фосфатидної кислоти
об’єднуються гліцероловим “містком”.
Кардіоліпіни
є “подвійними” фосфогліцеридами. У
них два залишки фосфатидної кислоти
об’єднуються гліцероловим “містком”.
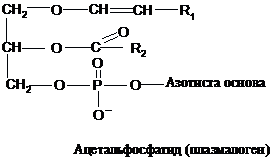
 До
фосфогліцеридів належать також
ацетальфосфатиди (плазмологени), які
відрізняються від розгянутих вище
представників тим, що замість одного
із залишків вищої жирної кислоти містять
?, ?-ненасичений спирт, який утворює етер
із залишком гліцерину. В розбавлених
кислотах вони гідролізують з утворенням
альдегіду відповідного ненасиченого
спирту.
До
фосфогліцеридів належать також
ацетальфосфатиди (плазмологени), які
відрізняються від розгянутих вище
представників тим, що замість одного
із залишків вищої жирної кислоти містять
?, ?-ненасичений спирт, який утворює етер
із залишком гліцерину. В розбавлених
кислотах вони гідролізують з утворенням
альдегіду відповідного ненасиченого
спирту.
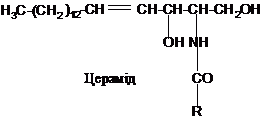
 Сфінгоміеліни
містять залишок ненасиченого аміноспирту
сфінгозину. Він з’єднується із залишком
вищої жирної кислоти амідним зв’язком.
Утворене похідне носить назву
цераміду:
Сфінгоміеліни
містять залишок ненасиченого аміноспирту
сфінгозину. Він з’єднується із залишком
вищої жирної кислоти амідним зв’язком.
Утворене похідне носить назву
цераміду:
 За
фізіологічних умов (рН біля 7,0) залишки
фосфату, аміно- і карбоксильні групи
полярних голів іонізовані.
Фосфоліпіди
є структурними компонентами біомембран.
На їх частку припадає до 50 % всіх мембранних
ліпідів. Кардіоліпіни у великих кількостях
містяться у внутрішніх мембранах
мітохондрій. Сфінгоміелінів особливо
багато в мієлінових оболонках нервових
клітин певного типу. Крім того вони
входять до складу ліпідів крові, багатьох
інших тканин.
Гліколіпіди
та сульфоліпіди
В
молекулах гліколіпідів полярним
компонентом служить не фосфат, а залишок
вуглеводу (галактози, глюкози, манози
або олігосахариду), з'єднаний з центральною
ланкою ефірним зв'язком. Центральною
ланкою може бути гліцерин (такі гліколіпіди
винайдені в бактеріях та зелених
рослинах), або сфінгозин у цереброзидах
та гангліозидах). Цереброзиди містять
залишок моно- або дисахариду:
За
фізіологічних умов (рН біля 7,0) залишки
фосфату, аміно- і карбоксильні групи
полярних голів іонізовані.
Фосфоліпіди
є структурними компонентами біомембран.
На їх частку припадає до 50 % всіх мембранних
ліпідів. Кардіоліпіни у великих кількостях
містяться у внутрішніх мембранах
мітохондрій. Сфінгоміелінів особливо
багато в мієлінових оболонках нервових
клітин певного типу. Крім того вони
входять до складу ліпідів крові, багатьох
інших тканин.
Гліколіпіди
та сульфоліпіди
В
молекулах гліколіпідів полярним
компонентом служить не фосфат, а залишок
вуглеводу (галактози, глюкози, манози
або олігосахариду), з'єднаний з центральною
ланкою ефірним зв'язком. Центральною
ланкою може бути гліцерин (такі гліколіпіди
винайдені в бактеріях та зелених
рослинах), або сфінгозин у цереброзидах
та гангліозидах). Цереброзиди містять
залишок моно- або дисахариду:
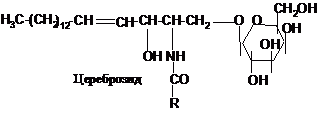 До
складу гангліозидів входять дуже складні
розгалужені олігосахариди, що містять
хоча б один залишок N-ацетилнейрамінової
(сіалової) кислоти:
До
складу гангліозидів входять дуже складні
розгалужені олігосахариди, що містять
хоча б один залишок N-ацетилнейрамінової
(сіалової) кислоти:
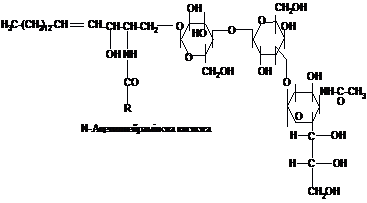 Вища
жирна кислота містить, як правило, 24
атоми С. В клітинних мембранах винайдено
більш ніж 15 класів гангліозидів.
Сульфоліпіди можна розглядати як похідні
гліколіпідів, в яких залишок сульфату
утворює естерний зв’язок з гідроксилом
вуглеводного компоненту.
Гліколіпіди,
як свідчать їх назви, знаходяться у
великих кількостях в нервовій тканині
- мієлінових оболонках нервів, сірій
речовині мозку, однак досить поширені
і в інших тканинах. Гангліозиди входять
до складу рецепторних ділянок мембран.
При деяких хворобах гліколіпіди
накопичуються в аномально великих
кількостях у зв'язку з порушенням
обміну.
Природні ліпіди утворюють
надзвичайно складні суміші, в яких
компоненти зустрічаються в різноманітних
поєднаннях. Ліпіди утворюють комплекси
з білками за рахунок гідрофобних
взаємодій з їх неполярними групами.
Такі утворення є формою транспорту
ліпідів в крові, характерні для клітинних
мембран.
2.4.
Біомембрани
Всі
живі клітини оточені і перетинаються
мембранами. Мембрани виконують такі
біологічні функції: бар’єрну (відокремлюють
клітину від зовнішнього середовища і
поділяють її на компартменти); транспортну
(забезпечують перенесення речовин між
клітиною і оточуючим середовищам і між
клітинними компартментами); структурну
(зумовлюють чітку організацію речовин
в клітині і збільшують ефективність
перетворень за рахунок імобілізації
(фіксації) ферментів і субстратів,
узгодженості процесів у просторі і
часі); сигнальну (забезпечують контакт
з іншими клітинами, передачу нервових
або гормональних сигналів).
На підставі
знань про властивості ліпідів, які
входять до складу мембран, їх взаємодію
з білками, вуглеводами та оточуючим
водним середовищем, Гортер та Грендель
ще в 1925 р. постулювали, що мембрани мають
двошарову структуру (модель "бутерброду"),
в якій дифільні молекули ліпідів
орієнтуються полярними головками до
оточуючого водного середовища, а
гідрофобними хвостами - всередину шару.
Уточнення структури та функції біомембран
за допомогою електронної мікроскопії
та хімічних методів привело до створення
сучасної рідинно-мозаїчної моделі
мембрани (С. Дж. Сінгер, Г. Ніколсон, 1972
р.), яка конкретизує уявлення про двошарову
структуру мембран (рис. 2.1).
На ліпіди
припадає біля 40 % сухої маси мембран.
В одній мембрані може поєднуватись
більше ста різних ліпідів, причому у
певному, визначеному для кожного типу
мембрани, порядку. Головними ліпідами
мембран є фосфоліпіди (біля 80 % всіх
ліпідів) і холестерин (3 – 15 %). Масове
співвідношення ліпід : білок становить
для типових мембран 1:1, а для міеліну –
4:1. Біля 5 % маси мембран припадає на
вуглеводи.
Дифільні молекули фосфоліпідів
утворюють подвійний шар товщиною біля
7 нм. За фізіологічних умов центральна
частина мембрани знаходиться в
рідкокристалічному стані. Ліпіди деяких
ракових клітин, що утворюють більш рідкі
плівки, ніж нормальні, визначають
аномальну поведінку цих клітин. Для
багатьох мембран, зокрема мітохондрій,
існування у вигляді рідких кристалів
забезпечується високим вмістом
ненасичених радикалів. Більш рідкий
стан плівки, що містить ненасичені
ланцюги, пояснюється тим, що в місцях
подвійних зв'язків порушується строга
паралельність ланцюгів, вони легко
згинаються і розміщуються хаотично. В
мембранах мієлінової оболонки рідкий
стан зумовлений надзвичайно високим
вмістом холестерину (біля 25 % всіх
ліпідів). В мембранах, багатих на
ненасичені радикали, холестерин, навпаки,
здійснює ущільнюючу дію, зменшуючи
нахил між ланцюгами і збільшуючи їх
впорядкованість. Отже холестерин у
мембрані має функцію регулятора
щільності, тобто агрегатного стану,
необхідного для нормального
функціонування.
Хімічний склад
гідрофобного шару мембрани визначає
важливі її властивості, наприклад,
проникність. При заміні ненасичених
радикалів в мембранах бактерій на
насичені, дифузія речовин сповільнюється
в 20 разів. Через мембрани легко проникає
вода, але вони практично непроникні для
йонів типу Na+, Cl- або Н+ і для полярних
молекул, наприклад глюкози. Таким
речовинам, на відміну від гідрофобних,
для проникнення через мембрану необхідні
спеціальні транспортні системи, або
переносники. Мембрани мають високий
електричний опір, тобто є гарними
ізоляторами. Полярні голови ліпідів,
особливо йонізовані, на поверхнях
мембрани утворюють кристалічну
впорядковану структуру завдяки міцним
взаємодіям протилежно заряджених та
полярних груп. Поверхня мембрани впливає
і на властивості близьких шарів води,
збільшуючи кислотність, понижуючи
температуру замерзання.
Ліпідний
склад біомембран може дещо змінюватись
залежно від харчування, від температурного
режиму існування. Наприклад, при пониженні
температури середовища мембрани бактерій
та рослин збагачуються ненасиченими
жирними кислотами, чим забезпечують
підтримання рідкого стану. Організми
теплокровних тварин протидіють
"затвердінню" мембран, підтримуючи
постійну температуру тіла за рахунок
обміну речовин. Цікаво адаптується до
низьких температур пінгвін. У нього
температура в ногах зменшується від
тіла до кінцівок і відповідно збільшується
вміст ненасичених жирних кислот в
мембранах клітин.
Білки біомембран
поділяють на два типи. Білки першого
типу - інтегровані, міцно зв'язані своїми
гідрофобними радикалами з гідрофобними
групами ліпідів. Деякі з них пронизують
мембрану наскрізь. До них належать
ферменти та транспортні білки. Білки
другого типу - периферійні, слабо
взаємодіють з мембраною, розташовуються
на її поверхні і притягуються до її
поверхні за рахунок електростатичної
взаємодії.
Деякі молекули досить
вільно переміщаються в мембрані - це
частина білків, ліпіди в площині мембрани.
Тому досить популярним є уявлення про
мембрану як про ліпідне море, в якому
плавають білкові айсберги. Ліпіди в
мембрані міняють своїх сусідів в
середньому 10 раз в секунду. Інколи,
приблизно 1 раз на 14 діб, молекула ліпіду
«перестрибує» з одного шару в інший. Це
переміщення називають «фліп-флоп».
Рідкість переміщень молекул ліпідів
між шарами зумовлена тим, що треба
затратити багато енергії, щоб перенести
полярну групу фосфоліпіду через
центральну, вуглеводневу частину
мембрани. Білкові молекули мембрани
можуть бути досить міцно закріплені за
внутрішні структури клітини (мікрофіламенти
та мікротрубочки).
Зовнішня та внутрішня
поверхні мембрани неоднакові. Білки
внутрішньої поверхні, утримуючись
внутрішньоклітинними структурами,
формують "скелет" (каркас) мембрани.
На зовнішній поверхні клітинних мембран
білки утворюють комплекси з вуглеводами
- глікопротеїди, які, можливо, забезпечують
злипання однотипних клітин. На ній також
містяться рецептори, які впізнають
гормони, ділянки, що визначають тканинну
сумісність, тобто є носіями антигенів
та інших імунологічних детермінант. До
складу цих специфічних структур входять
гангліозиди та глікопротеїди. Розгалужені
олігосахаридні ланцюги розташовуються
як “антени” на зовнішньому боці
клітинних мембран, утворюючи “пухнасту”
оболонку, товщина якої становить біля
100 нм, тобто приблизно в 10 р. більша
товщини подвійного ліпідного шару. У
внутрішніх мембранах клітини також
міститься цілий ряд молекул та комплексів,
які виконують свою функцію тільки при
умові її узгодженості і певній орієнтації
мембрани.
Вища
жирна кислота містить, як правило, 24
атоми С. В клітинних мембранах винайдено
більш ніж 15 класів гангліозидів.
Сульфоліпіди можна розглядати як похідні
гліколіпідів, в яких залишок сульфату
утворює естерний зв’язок з гідроксилом
вуглеводного компоненту.
Гліколіпіди,
як свідчать їх назви, знаходяться у
великих кількостях в нервовій тканині
- мієлінових оболонках нервів, сірій
речовині мозку, однак досить поширені
і в інших тканинах. Гангліозиди входять
до складу рецепторних ділянок мембран.
При деяких хворобах гліколіпіди
накопичуються в аномально великих
кількостях у зв'язку з порушенням
обміну.
Природні ліпіди утворюють
надзвичайно складні суміші, в яких
компоненти зустрічаються в різноманітних
поєднаннях. Ліпіди утворюють комплекси
з білками за рахунок гідрофобних
взаємодій з їх неполярними групами.
Такі утворення є формою транспорту
ліпідів в крові, характерні для клітинних
мембран.
2.4.
Біомембрани
Всі
живі клітини оточені і перетинаються
мембранами. Мембрани виконують такі
біологічні функції: бар’єрну (відокремлюють
клітину від зовнішнього середовища і
поділяють її на компартменти); транспортну
(забезпечують перенесення речовин між
клітиною і оточуючим середовищам і між
клітинними компартментами); структурну
(зумовлюють чітку організацію речовин
в клітині і збільшують ефективність
перетворень за рахунок імобілізації
(фіксації) ферментів і субстратів,
узгодженості процесів у просторі і
часі); сигнальну (забезпечують контакт
з іншими клітинами, передачу нервових
або гормональних сигналів).
На підставі
знань про властивості ліпідів, які
входять до складу мембран, їх взаємодію
з білками, вуглеводами та оточуючим
водним середовищем, Гортер та Грендель
ще в 1925 р. постулювали, що мембрани мають
двошарову структуру (модель "бутерброду"),
в якій дифільні молекули ліпідів
орієнтуються полярними головками до
оточуючого водного середовища, а
гідрофобними хвостами - всередину шару.
Уточнення структури та функції біомембран
за допомогою електронної мікроскопії
та хімічних методів привело до створення
сучасної рідинно-мозаїчної моделі
мембрани (С. Дж. Сінгер, Г. Ніколсон, 1972
р.), яка конкретизує уявлення про двошарову
структуру мембран (рис. 2.1).
На ліпіди
припадає біля 40 % сухої маси мембран.
В одній мембрані може поєднуватись
більше ста різних ліпідів, причому у
певному, визначеному для кожного типу
мембрани, порядку. Головними ліпідами
мембран є фосфоліпіди (біля 80 % всіх
ліпідів) і холестерин (3 – 15 %). Масове
співвідношення ліпід : білок становить
для типових мембран 1:1, а для міеліну –
4:1. Біля 5 % маси мембран припадає на
вуглеводи.
Дифільні молекули фосфоліпідів
утворюють подвійний шар товщиною біля
7 нм. За фізіологічних умов центральна
частина мембрани знаходиться в
рідкокристалічному стані. Ліпіди деяких
ракових клітин, що утворюють більш рідкі
плівки, ніж нормальні, визначають
аномальну поведінку цих клітин. Для
багатьох мембран, зокрема мітохондрій,
існування у вигляді рідких кристалів
забезпечується високим вмістом
ненасичених радикалів. Більш рідкий
стан плівки, що містить ненасичені
ланцюги, пояснюється тим, що в місцях
подвійних зв'язків порушується строга
паралельність ланцюгів, вони легко
згинаються і розміщуються хаотично. В
мембранах мієлінової оболонки рідкий
стан зумовлений надзвичайно високим
вмістом холестерину (біля 25 % всіх
ліпідів). В мембранах, багатих на
ненасичені радикали, холестерин, навпаки,
здійснює ущільнюючу дію, зменшуючи
нахил між ланцюгами і збільшуючи їх
впорядкованість. Отже холестерин у
мембрані має функцію регулятора
щільності, тобто агрегатного стану,
необхідного для нормального
функціонування.
Хімічний склад
гідрофобного шару мембрани визначає
важливі її властивості, наприклад,
проникність. При заміні ненасичених
радикалів в мембранах бактерій на
насичені, дифузія речовин сповільнюється
в 20 разів. Через мембрани легко проникає
вода, але вони практично непроникні для
йонів типу Na+, Cl- або Н+ і для полярних
молекул, наприклад глюкози. Таким
речовинам, на відміну від гідрофобних,
для проникнення через мембрану необхідні
спеціальні транспортні системи, або
переносники. Мембрани мають високий
електричний опір, тобто є гарними
ізоляторами. Полярні голови ліпідів,
особливо йонізовані, на поверхнях
мембрани утворюють кристалічну
впорядковану структуру завдяки міцним
взаємодіям протилежно заряджених та
полярних груп. Поверхня мембрани впливає
і на властивості близьких шарів води,
збільшуючи кислотність, понижуючи
температуру замерзання.
Ліпідний
склад біомембран може дещо змінюватись
залежно від харчування, від температурного
режиму існування. Наприклад, при пониженні
температури середовища мембрани бактерій
та рослин збагачуються ненасиченими
жирними кислотами, чим забезпечують
підтримання рідкого стану. Організми
теплокровних тварин протидіють
"затвердінню" мембран, підтримуючи
постійну температуру тіла за рахунок
обміну речовин. Цікаво адаптується до
низьких температур пінгвін. У нього
температура в ногах зменшується від
тіла до кінцівок і відповідно збільшується
вміст ненасичених жирних кислот в
мембранах клітин.
Білки біомембран
поділяють на два типи. Білки першого
типу - інтегровані, міцно зв'язані своїми
гідрофобними радикалами з гідрофобними
групами ліпідів. Деякі з них пронизують
мембрану наскрізь. До них належать
ферменти та транспортні білки. Білки
другого типу - периферійні, слабо
взаємодіють з мембраною, розташовуються
на її поверхні і притягуються до її
поверхні за рахунок електростатичної
взаємодії.
Деякі молекули досить
вільно переміщаються в мембрані - це
частина білків, ліпіди в площині мембрани.
Тому досить популярним є уявлення про
мембрану як про ліпідне море, в якому
плавають білкові айсберги. Ліпіди в
мембрані міняють своїх сусідів в
середньому 10 раз в секунду. Інколи,
приблизно 1 раз на 14 діб, молекула ліпіду
«перестрибує» з одного шару в інший. Це
переміщення називають «фліп-флоп».
Рідкість переміщень молекул ліпідів
між шарами зумовлена тим, що треба
затратити багато енергії, щоб перенести
полярну групу фосфоліпіду через
центральну, вуглеводневу частину
мембрани. Білкові молекули мембрани
можуть бути досить міцно закріплені за
внутрішні структури клітини (мікрофіламенти
та мікротрубочки).
Зовнішня та внутрішня
поверхні мембрани неоднакові. Білки
внутрішньої поверхні, утримуючись
внутрішньоклітинними структурами,
формують "скелет" (каркас) мембрани.
На зовнішній поверхні клітинних мембран
білки утворюють комплекси з вуглеводами
- глікопротеїди, які, можливо, забезпечують
злипання однотипних клітин. На ній також
містяться рецептори, які впізнають
гормони, ділянки, що визначають тканинну
сумісність, тобто є носіями антигенів
та інших імунологічних детермінант. До
складу цих специфічних структур входять
гангліозиди та глікопротеїди. Розгалужені
олігосахаридні ланцюги розташовуються
як “антени” на зовнішньому боці
клітинних мембран, утворюючи “пухнасту”
оболонку, товщина якої становить біля
100 нм, тобто приблизно в 10 р. більша
товщини подвійного ліпідного шару. У
внутрішніх мембранах клітини також
міститься цілий ряд молекул та комплексів,
які виконують свою функцію тільки при
умові її узгодженості і певній орієнтації
мембрани.