

Лекция 4. Физиология спинного и заднего мозга. Физиология мозжечка. Физиология среднего мозга.
Слайд 1
К ЦНС принято относить спинной мозг (СМ) и головной мозг (ГМ). СМ расположен в позвоночном канале, имеет сегментарное строение. Сегмент – это участок СМ от которого отходит пара спинномозговых нервов. СМ человека состоит из 31 сегмента: 8 шейных (C I-VIII), 12 грудных (Th I-XII), 5 поясничных (L I-V), 5 крестцовых (S I-V) и 1 копчиковый (Co I).
СМ обеспечивает выполнения целого ряда спинномозговых рефлексов, участвует в регуляции висцеральных функций (регулирует работу внутренних органов и систем) и осуществляет проводниковую деятельность.
Принципы и закономерности функциональной организации СМ.
1. Рефлекторный принцип.
2. Метамерная организация соматических спинномозговых рефлексов. На раннем этапе эмбриогенеза формирование СМ сопровождается закладкой парных зачатков тканей сомитов, состоящих из дерматомов и миотомов (кожных и мышечных сегментов – метамеров). Афферентные нейроны каждого сегмента СМ иннервируют определенные области дерматома (зачаточного сегмента кожи). Количество СМ сегментов соответствует числу метамеров, однако каждый метамер получает иннервацию от двух-трех смежных сегментов СМ. Благодаря этой особенности каждый соматический рефлекс имеет определенное рецептивное поле (рис. 1).
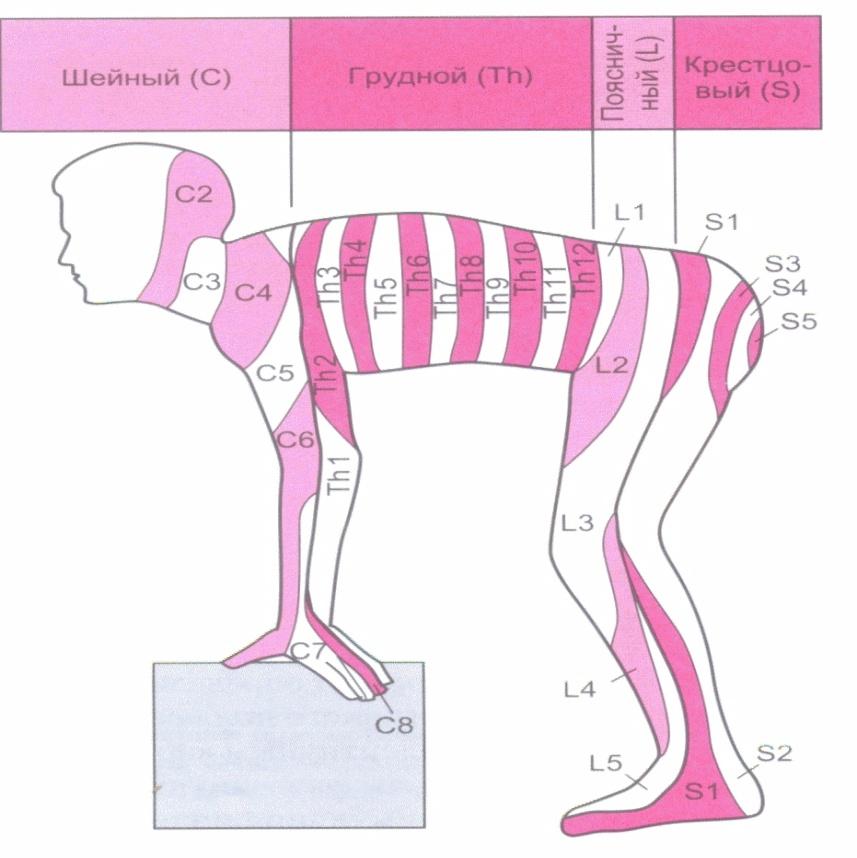
Рис. 1. Схема метамерной организации спинномозговых рефлексов
3. Эфферентные нейроны СМ представляют собой общий конечный путь для иррадиирущих в ЦНС возбуждений (закон общего конечного пути Шеррингтона).
4. Афферентные нервные волокна входят в состав задних корешков СМ, а передние корешки СМ образованы аксонами двигательных нейронов (закон о неоднородности передних и задних корешков СМ – закон Белла-Мажанди).
В сером веществе спинного мозга находятся тела двигательных (мотонейроны), вставочных и вегетативных нейронов.
Мотонейроны. Различают крупные, или a‑мотонейроны, и мелкие, или γ‑мотонейроны.
a–Мотонейроны входят в состав медиальных и латеральных ядер. Это наиболее крупные клетки спинного мозга. Их аксоны в составе двигательных нервных волокон типа Aa образуют нервно-мышечные синапсы с экстрафузальными (сократительными) поперечнополосатыми мышечными волокнами (МВ) скелетной мышцы и участвуют в формировании двигательных единиц. Аксоны a‑мотонейронов посылают коллатерали к вставочным нейронам (клетки Реншоу), образующим тормозные синапсы с a‑мотонейронами.
γ–Мотонейроны располагаются вместе с a‑мотонейронами в передних рогах спинного мозга. Они значительно меньше a‑мотонейронов и передают ПД по нервных волокнам, осуществляя двигательную иннервацию интрафузальных МВ в составе мышечных веретён.
Вставочные нейроны (интернейроны) получают информацию от одних нейронов и передают её другим. Аксоны вставочных нейронов участвуют также в образовании проводящих путей. Классические примеры вставочных нейронов — клетки Реншоу. Количество вставочных нейронов в 30 раз превышает количество мотонейронов. Вставочные нейроны небольшого размера, их возбудимость высокая, часто они спонтанно активны, генерируют импульсы частотой до 1000-1500 Гц (имп/сек), имеют много связей друг с другом, часть из них синаптически контактирует с мотонейронами. Особые свойства вставочных нейронов (дивергенция, конвергенция, повторные разряды, латеральное торможение, торможение через клетки Реншоу) имеют непосредственное отношение к моторным функциям спинного мозга. Только некоторые сигналы, поступающие из спинномозговых нервов, или сигналы из мозга сразу передаются к мотонейронам, практически все сигналы попадают к вставочным нейронам и только после того, как они интегрируются в с сигналами из других источников, они окончательно конвергируют на мотонейроны.
Вегетативные нейроны расположены в висцеральных ядрах грудного и поясничного отделов, а также в крестцовом отделе (ядро Онуфровича) промежуточной зоны серого вещества. Нейроны образуют синаптические контакты с афферентными волокнами общей висцеральной чувствительности (общие висцеральные афференты). Ядро Онуфровича содержит преганглионарные парасимпатические нейроны. Их аксоны проходят в передних корешках крестцового отдела.
Спинной мозг играет важную роль в обеспечении тонических и позных соматических рефлексов. Даже в расслабленном состоянии тела благодаря постоянным эфферентным возбуждениям все мышцы находятся в некотором тоническом напряжении. Спинной мозг может в определенной степени поддерживать естественную позу тела децеребрального животного.
Мышца как орган имеет свои рецепторы и афферентные пути. Рецепторами мышц являются интрафузальные мышечные волокна (мышечные веретена), от которых отходят афферентные нервные волокна (Ia), сигнализирующие о растяжении мышцы (рис. 2).
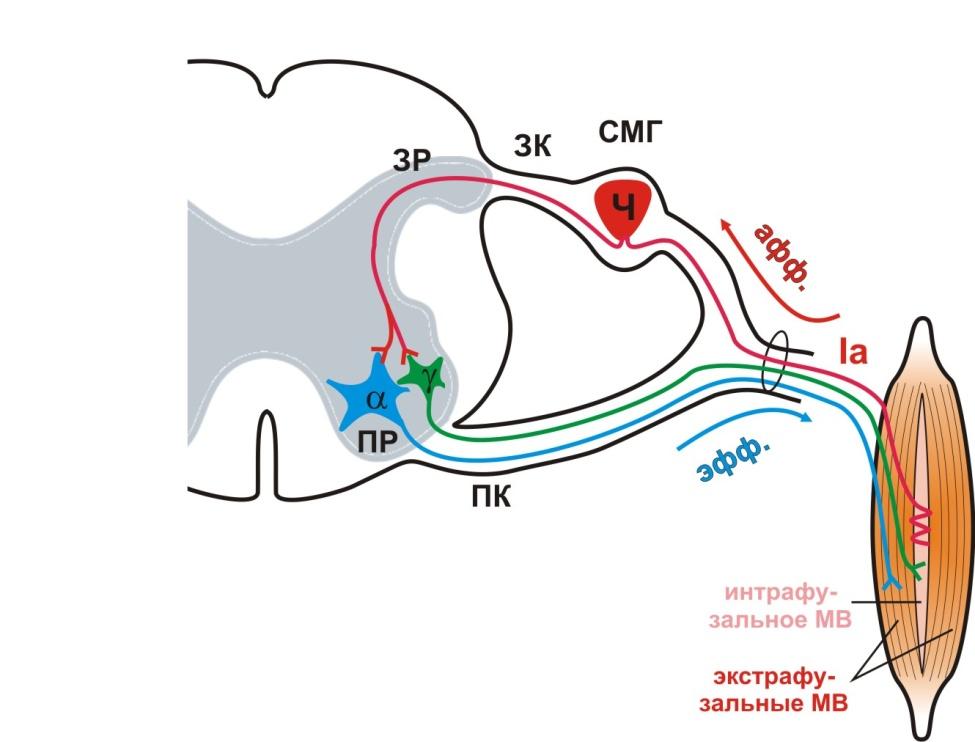
Рис. 2. Схема иннервации мышцы
При растяжении мышцы под нагрузкой происходит растяжение мышечных веретен и возбуждение рецепторных окончаний Ia афферентных волокон. Эти возбуждения передаются на a-мотонейроны сократительных (экстрафузальных) мышечных волокон, что обеспечивает повышение тонуса мышцы или вызывает (при резком ее растяжении) сокращение мышцы (рефлекс растяжения илимиотатический рефлекс). Параллельно афферентные Ia возбуждения конвергируют на g-мотонейроны, которые регулируют напряжение мышечных веретен (рис. 6).
Второй тип мышечных рецепторов - сухожильные рецепторы Гольджи – обеспечивают тоническое напряжение и сокращение мышц в зависимости от растяжения или раздражения сухожилий. Некоторые сухожильные рефлексы вам должны быть знакомы: коленный рефлекс, плечевой рефлекс (рис. 3).

Рис. 3. Схема коленного рефлекса
При легком ударе неврологическим молоточком по сухожилию 4-х главой мышцы бедра возбуждение рецепторов Гольджи (РГ) возбуждение распространяется по Ib афферентным волокнам на a-мотонейроны мышц разгибателей (рис. 3). Одновременно это возбуждение через тормозный синапс вставочного нейрона (В) вызывает торможение a-мотонейронов мышц сгибателей, вызывая из расслабление (реципрокное торможение). Реципрокное торможение обеспечивает согласованность процессов сокращения и расслабление мышц антагонистов при рефлексах сгибания и разгибания конечностей.
Спинной мозг способен самостоятельно обеспечивать более сложные циклические рефлексы: шагательный, прыгательный, плавательный, чесательный (циклические потому, что состоят из последовательно повторяющихся двигательных актов).
Однако, в норме, деятельность СМ находится под контролем ГМ, который постоянно осуществляет более тонкую подстройку спинномозговых соматических рефлексов. Выделяют несколько уровней такого контроля: со стороны вестибулярных ядер (ВЯ), нейронов ретикулярной формации (РФ) и нейронов моторной коры (МК) (рис. 4).
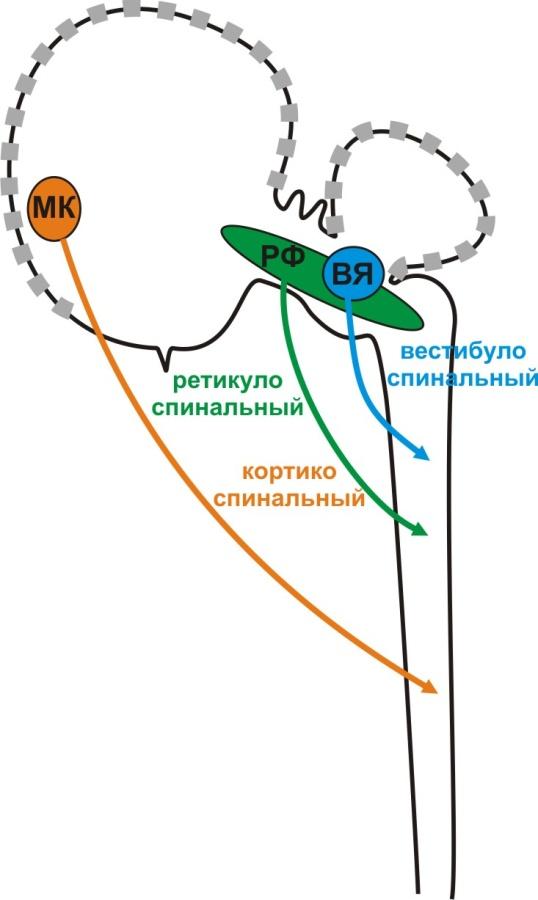
Рис. 4. Контроль головного мозга за деятельностью спинного мозга
Нарушение взаимодействия спинного и головного мозга в результате травмы или воспалительного процесса приводит к развитиюспинального шока. Тяжесть спинального шока (СШ) зависит не только от характера повреждения, но и от уровня организации ЦНС. У лягушки (земноводных) спинальный шок после отделения ГМ от СМ продолжается несколько минут, у млекопитающих СШ может продолжаться неделями и месяцами. При СШ наблюдается выпадение рефлексов, спинномозговые центры которых расположеныкаудальнее точки повреждения.
При половинной перерезке (пересечении) СМ возникает синдром Броун-Секара, на стороне повреждения (ниже места поражения) развивается паралич двигательной системы и нарушается проприоцептивная чувствительность, а на противоположной стороне туловища нарушается болевая чувствительность.
В спинном мозге расположены центры вегетативной нервной системы: симпатического отдела - в грудных и поясничных сегментах, парасимпатического отдела – крестцовых. Роль СМ в регуляции вегетативных функций мы будем рассматривать в разделе «физиология вегетативной нервной системы».
Животные, у которых произведено пересечение СМ на границе с продолговатым, получили название – спинальные животные. У таких животных отсутствует самостоятельное дыхание, низкое кровяное давление и тонус сосудов, они не могут поддерживать постоянную температуру тела, у них отсутствуют формы целенаправленной деятельности (пищевой рефлекс, оборонительный и т.п.)
Физиология продолговатого мозга и моста.
Задний мозг, куда относят функционально близкие отделы ГМ - продолговатый мозг и мост, играет важную роль в поддержании тонических и позных рефлексов, участвует в регуляции висцеральных функций и осуществляет проводниковую деятельность.
Принято выделять статические и статокинетические рефлексы заднего мозга. К статическим рефлексам относят тонические и позные рефлексы в отсутствие перемещения тела в пространстве. Значение заднего мозга в тонических и позных рефлексах определяется регулирующим влияние на эфферентные нейроны СМ со стороны нейронов вестибулярных ядер (ядра преддверно-улиткового нерва -VIII пары черепных нервов). Важную роль в статических позных рефлексах играют афферентные возбуждения от мышц шеи (отсюда специальное название этих рефлексов – тонические шейные лабиринтные рефлексы).
Шейные тонические лабиринтные рефлексы впервые описаны Р. Магнусом на кошках с перерезанным мозгом на уровне шатра мозжечка. Запрокидывание головы кошки назад усиливает тонус мышц-разгибателей в передних конечностях и ослабляет в задних. Наклон головы вниз вызывает противоположенный эффект (рис. 5).
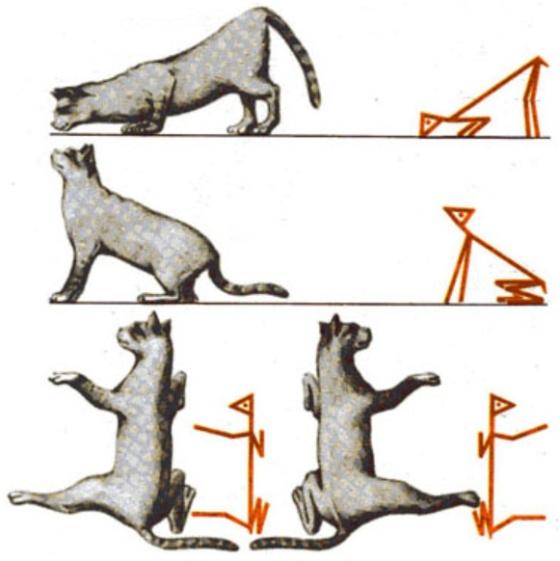
Рис. 5. Шейные тонические рефлексы кошки (по Р. Магнусу)
Нарушение равновесия за счет поворота головы вправо или влево относительно продольной оси вызывает компенсаторное усиление тонуса разгибателей тех конечностей, в сторону которых повернута голова (рефлекс вращения по Магнусу, рис. 5).
Статокинетические рефлексы обеспечивают поддержание позы тела в пространстве в момент его перемещения. Всем знакомы лифтные рефлексы. В начальный момент движения лифта вверх может произойти сгибание в коленных суставах, но в норме мы не падаем по причине повышения тонуса мышц-разгибателей. При движении лифта вниз, напротив, тонус разгибателей снижается. Почти каждому известны головной и зрительный нистагм. Они, например, проявляются в форме продолжающегося движения головы и глазных яблок после остановки вращения человека вокруг своей оси.
В заднем мозге расположены ядра 8-ми черепных нервов (ЧН): V пара - тройничный; VI пара – отводящий; VII – лицевой; VIII пара – преддверно-улитковый; IX пара – языкоглоточный; X пара – блуждающий; XI пара – добавочный; XII пара – подъязычный.
На нейронах ядер ЧН замыкаются рефлекторные дуги важнейших безусловных защитных рефлексов: мигательного (роговичного; V и VII), рвотного (V, IX, X), чихательного (V, VII, IX, X, XII), кашлевого (V, VII, IX, X).
В заднем мозге расположены центры рефлексов пищевого поведения: жевания, сосания, глотания. Глотательный цепной рефлекс состоит из трех последовательных фаз: ротовой, глоточной и пищеводной. При раздражении пищевым комком рецепторов корня языка и задней стенки зева возбуждение по чувствительным волокнам V, IX и X ЧН направляется к двигательным нейронам двойного ядра (общее двигательное ядро для IX и X ЧН) и тройничного нерва, затем по эфферентным волокнам указанных нервов – к мышцам языка и глотки.
В ретикулярной формации продолговатого мозга расположены центры регуляции вегетативных функций: дыхательный центр и сосудодвигательный центр.
В XIX веке М. Флуранс обнаружил, что повреждение каудальной части дна IV желудочка приводит к остановке дыхания (укол Флуранса). Позднее Н. А. Миславский (1885) выделил в дыхательном центре две функционально различные части: инспираторнуюи экспираторную, активность нейронов которых коррелирует с фазами дыхательного цикла (вдохом – инспирацией, и выдохом – экспирацией)
Нейроны ДЦ обладают автоматией, поэтому дыхательный центр является ритмоводителем дыхания (задает ритм дыхания частотой 15-16 дыхательных циклов в минуту). По этой причине травмы основания черепа опасны и могут привести к остановке дыхания в результате нарушения функций нейронов ДЦ. Эфферентные возбуждения нейронов ДЦ по ретикуло-спинномозговому пути переключаются на дыхательные мотонейроны СМ и в составе диафрагмального и межреберных нервов направляются к одноименным мышцам. Еще одна особенность ДЦ заключается в реципрокном характере активности инспираторных и экспираторных нейронов (рис. 6).
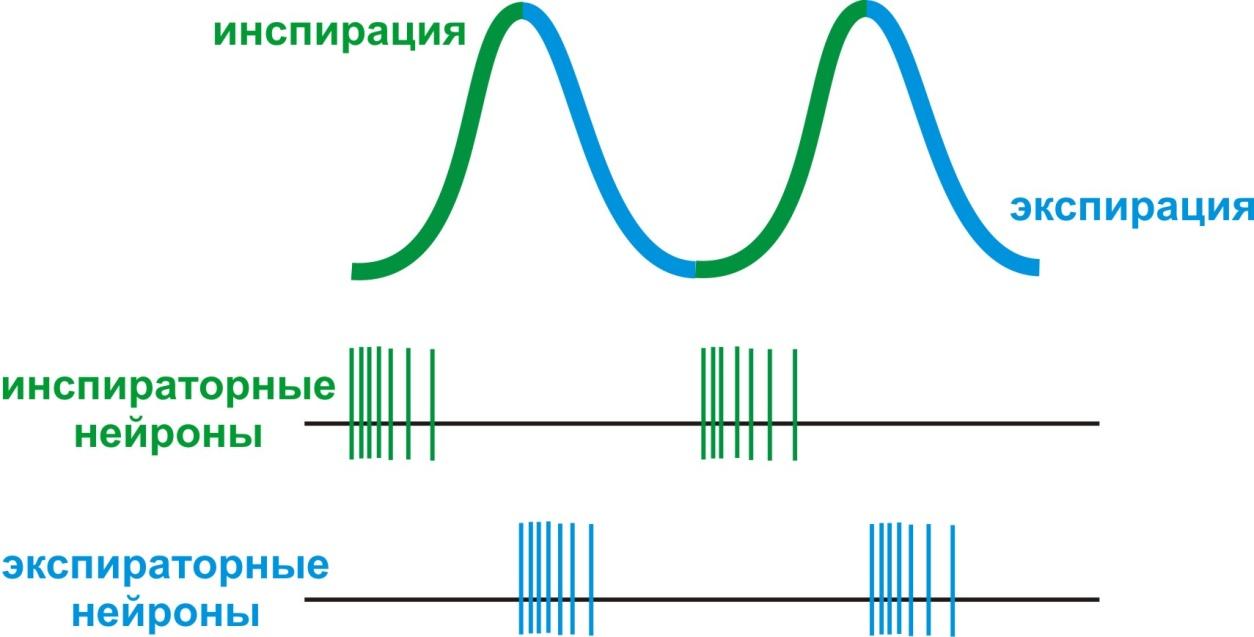
Рис. 6. Реципрокные взаимосвязи нейронов дыхательного центра
Вторым центром регуляции дыхания является пневмотаксический центр (ПТЦ), расположенный в мосту. ПТЦ не является ритмоводителем, но играет важную роль в регуляции глубины и частоты дыхания.
Другим жизненно важным центром ретикулярной формации продолговатого мозга является сосудодвигательный центр (СДЦ), локализация которого впервые была определена Ф. В. Овсянниковым (1871). Сосудодвигательный или вазомоторный центр расположен в дорсолатеральном направлении от дна IV мозгового желудочка до пирамид.
Установлено, что стимуляция ростральных частей СДЦ увеличивает тонус гладких мышц кровеносных сосудов – вазоконстрикцию (тем самым повышает артериальное давление - АД) и вызывает тахикардию (увеличение частоты сердечных сокращений). Стимуляция каудального отдела СДЦ, напротив, вызывает расширение сосудов (вазодилатацию), снижение АД и брадикардию (снижение частоты сердцебиения). Нейроны, активность которых приводит к вазоконстрикции, получили название прессорных нейронов, а нейроны, вызывающие дилатацию – депрессорных. Активность прессорных нейронов определяется тоническим влиянием симпатической нервной системы.
Задний мозг участвует в регуляции вегетативных функций. Это связано с расположением парасимпатических нейронов в составе некоторых черепных нервов: лицевой и языкоглоточный ЧН имеют слюноотделительные ядра (верхнее и нижнее, соответственно). Основным парасимпатическим нервом является блуждающий нерв (n. vagus), который регулирует работу органов грудной и верхней части брюшной полостей.
Подробнее о парасимпатических регуляциях мы поговорим в разделе «физиология вегетативной нервной системы».
Животные, у которых произведено пересечение ГМ по переднему краю варолиева моста, получили название – бульбарные животные. Такие животные самостоятельно дышат, поддерживают постоянный уровень кровяного давления, но не могут поддерживать постоянную температуру тела и передвигаться (но лягушка, кролик способны осуществлять некоторые виды движений).