

1 . История развития генетических исследований.
Истоки генетики, как и всякой науки, следует искать в практике. Генетика возникла в связи с разведением домашних животных и возделыванием растений, а также с развитием медицины. С тех пор как человек стал применять скрещивание животных и растений, он столкнулся с тем фактом, что свойства и признаки потомств зависят от свойств избранных для скрещивания родительских особей. Отбирая и скрещивая лучших потомков, человек из поколения в поколение создавал родственные группы - линии, а затем породы и сорта с характерными для них наследственными свойствами. Генетика – наука о наследственности и ее изменчивости – получила развитие в начале XX в., после того как исследователи обратили внимание на законы Г. Менделя, открытые в 1865 г., но остававшиеся без внимания в течение 35 лет. В короткий срок генетика выросла в разветвленную биологическую науку с широким кругом экспериментальных методов и направлений. Ее бурное развитие было обусловлено как запросами сельского хозяйства, нуждавшегося в детальной разработке проблем наследственности у растений и животных, так и успехами биологических дисциплин, таких, как морфология, эмбриология, цитология, физиология и биохимия, подготовивших почву для углубленного изучения законов наследственности и материальных носителей наследственных факторов. Название генетика было предложено для новой науки английским ученым У. Бэтсоном в 1906 г.
Развитию науки о наследственности и изменчивости особенно сильно способствовало учение Ч. Дарвина о происхождении видов, которое внесло в биологию исторический метод исследования эволюции организмов. Сам Дарвин приложил немало усилий для изучения наследственности и изменчивости. Он собрал огромное количество фактов, сделал на их основе целый ряд правильных выводов, однако ему не удалось установить закономерности наследственности.
Его современники, так называемые гибридизаторы, скрещивавшие различные формы и искавшие степень сходства и различия между родителями и потомками, также не смогли установить общие закономерности наследования.
В истории развития генетики можно выделить три важных или основных этапа:
1)первый этап (с 1900 г. ~ до 1912 г.) – период триумфального шествия менделизма, утверждения открытых Менделем законов наследственности гибридологическими опытами, проведенными в разных странах на высших растениях и животных (лабораторных грызунах, курах, бабочках и др.), в результате чего выяснилось, что законы эти имеют универсальный характер. В течение немногих лет генетика оформилась как самостоятельная биологическая дисциплина и получила широкое признание.
2)Второй этап характеризуется переходом к изучению явлений наследственности на клеточном уровне, а также на данном этапе рассматривается феномен «лысенковщины». Главной отличительной чертой второго этапа истории генетики (~ 1912 до 1925 г.) было создание и утверждение хромосомной теории наследственности. Ведущую роль в этом сыграли экспериментальные работы американского генетика Т.Моргана (1861-1945) и трёх его учеников – А.Стертеванта, К.Бриджеса, Г.Меллера, проведённые на плодовой мушке дрозофиле , которая благодаря ряду своих свойств (удобству содержания в лаборатории, быстроте размножения, высокой плодовитости, малому числу хромосом) стала с тех пор излюбленным объектом генетических исследований.
3)Третий этап истории генетики (~ 1925 – 1940 г.) ознаменован в первую очередь открытием возможности искусственно вызвать мутации. До тех пор существовала ошибочная концепция, что мутации возникают в организме самопроизвольно, под влиянием каких-то чисто внутренних причин.
4)Наиболее характерными чертами четвёртого этапа истории генетики (1940-1955) было развитие работ по генетике физиологических и биохимических признаков и вовлечение в круг генетического эксперимента микроорганизмов и вирусов, что повысило разрешающую способность генетического анализа. Изучение биохимических процессов, лежащих в основе формирования наследственных признаков разных организмов, пролило свет на то, как действуют гены и, в частности, привело к важному обобщению, сделанному американскими генетиками Дж. Бидлом и Э.Тэтумом, согласно которого всякий ген определяет синтез в организме одного фермента (эта формула: «один ген – один фермент» впоследствии: «один ген – один белок»).
5)современный этап развития генетики Для последнего современного этапа истории генетики, начавшегося приблизительно в середине 1950-х г., наиболее характерно исследование генетических
явлений на молекулярном уровне благодаря внедрению в генетику новых химических, физических, математических подходов и методов, совершенных приборов и сложных реактивов.
В результате беспрецедентно быстрого прогресса в области молекулярной биологии и молекулярной генетики, появления в последнее десятилетие принципиально новых методов манипулирования с генетическим материалом, положивших начало генетической инженерии, был полностью раскрыт генетический код (в этой расшифровке большую роль сыграли работы Крика и его сотрудников в Англии, С.Очоа и М.Ниренберга в Америке), удалось выделить отдельные гены и установить их нуклеиновую последовательность, понять тонкое строение генов различных про – эукариотов, изучить принципы регуляции генной активности. В 1969 г. в США Г.Корана с сотрудниками синтезировали химическим путём вне организма первый простой по своей структуре ген (один из генов дрожжей), а в начале 1970-х годов в ряде американских лабораторий, а затем в лабораториях других стран, в том числе в СССР, иным способом – с помощью особых ферментов – были синтезированы вне организма много гораздо более крупных и сложноустроенных генов про- и эукариотов.
достижения молекулярной биологии и связан с использованием методов и принципов точных наук - физики, химии, математики, биофизики и др.- в изучении явлений жизни на уровне молекул.
2 . Вклад отечественных ученых в развитие общей и медицинской генетики.
Начало развития генетики в нашей стране приходится на первые годы Советской власти. В 1919 г. в Петроградском университете была создана кафедра генетики, которую возглавил Юрий Александрович Филипченко (1882–1930). В 1930 г. открылась Лаборатория генетики Академии наук
СССР под руководством Николая Ивановича Вавилова (с 1933 г. – Институт генетики).В 1920–1930-е гг. наша страна лидировала по всем разделам генетики. в рамках евгеники в СССР оформилось и получило развитие действительно строго научное направление исследований роли наследственных факторов в становлении различных нормальных и патологических признаков у человека, получившее название «медицинская генетика». Евгенический кризис для советских исследователей был преодолен довольно легко.Медицинская генетика как наука прикладная пыталась реализовать новые научные знания о природе наследственных заболеваний таким образом, чтобы извлечь из них практическую пользу в плане диагностики или лечения наследственных заболеваниКоротко остановимся на истории медицинской генетики в России. Заслуживает упоминания капитальный труд В.М. Флоринского (1833—1899) «Усовершенствование и вырождение человеческого рода», впервые увидевший свет в 1866 г. и переизданный в 1926 г. В своей работе В.М.Флоринский, профессор Медико-хирургической академии в С.-Петербурге, рассмотрел широкий круг вопросов от строения яйца, сперматозоида и оплодотворения до необходимости социального совершенствования общества в целях гармонического развития народа. В.М. Флоринский четко выделял ряд заболеваний наследственной природы, которые чаще возникают у детей супругов, состоящих в родственных браках (глухота, пигментный ретинит, альбинизм, некоторые врожденные уродства), рассматривал положительную роль смешения народов. В отличие от Гальтона В.М.Флоринский кладет в основу своей гигиены бракосочетания «прививку» населению здорового выработанного вкуса.
Николай Константинович Кольцов Еще в 1927 г. Н. Кольцов в своих работах высказал предполо жение о существовании
«наследственных молекул». В настоящее время мы знаем, что этим он предвидел матричный принцип ре пликации ДНК.
Кроме гипотезы молекулярного строения и матричной репро дукции хромосом, Николай Владимирович Тимофеев-Ресовский родился в 1900 г. После окончания Московского
университета он работал в Институте экспериментальной биологии, которым руководил Н. Кольцов. Необыкновенная эрудиция, широта научных знаний позволи ли ученому работать в различных областях биологии. Но основ ными его исследованиями были работы в области радиационной генетики, молекулярной биологии, радиобиологии.
Он изучил влияние радиации на процесс передачи наслед ственных признаков, рассматривал гены как мишени, находя щиеся под действием ионизирующей радиации.
1)Кольцов Николай Константинович (1872–1940) – предсказал свойства носителей генетической информации; разрабатывал теорию гена; разрабатывал учение о социальной генетике (евгенике).
2)Вавилов Николай Иванович (1887–1943) – сформулировал закон гомологических рядов, разработал учение о виде как системе.
3)Мичурин Иван Владимирович (1855–1935) – открыл возможность управления доминированием.
4)Серебровский Александр Сергеевич (1892–1948) – создал учение о генофонде и геногеографии: «Совокупность всех генов данного вида я назвал генофондом, чтобы подчеркнуть мысль о том, что в лице генофонда мы имеем такие же национальные богатства, как и в лице наших запасов угля, скрытых в наших недрах».
5)Четвериков Сергей Сергеевич (1880–1959) – в работе «О некоторых моментах эволюционного процесса с точки зрения современной генетики» доказал генетическую неоднородность природных популяций.
6)Дубинин Николай Петрович (1907–1998) – доказал делимость гена; независимо от западных исследователей установил, что важную роль в эволюции играют вероятностные, генетикоавтоматические процессы.
7)Шмальгаузен Иван Иванович (1884–1963) – разработал теорию стабилизирующего отбора; открыл принцип интеграции биологических систем.
8)Николай Владимирович Тимофеев-Ресовский (1900–1981) – заложил основы современной генетики популяций.
3. Наследственность и изменчивость живого, их формы.
Наследственностью называется свойство организмов обеспечивать материальную и функциональную преемственность между поколениями ,а так же специфический характер индивидуального развития организмов в определенных условиях среды. Благодаря наследственности у родителей и детей обнаруживается сходство морфологических признаков, химического состава тканей, физиологических отравлений и других особенностей. Вследствие этого каждый вид организмов воспроизводит себя в ряду поколений.
Изменчивость - это явление противоположное наследственности. Заключается в изменении наследственных задатков, а так же их проявлении в процессе развития организмов.
Виды изменчивости
Различают наследственную и ненаследственную изменчивость.
Наследственная (генотипическая) изменчивость связана с изменением самого генетического материала. Ненаследственная (фенотипическая, модификационная) изменчивость — это способность организмов изменять свой фенотип под влиянием различных факторов. Причиной модификационной изменчивости являются изменения внешней среды обитания организма или его внутренней среды.
Формы изменчивости.
Биологическую изменчивость подразделяют на фенотипическую и генотипическую. Классификация по характеру изменения признаков и механизму:
Ненаследственная:
-фенотипическая.
Частный случай фенотипической изменчивости – фенокопии. Фенокопии – вызванные условиями внешней среды фенотипические модификации, имитирующие генетические признаки. Под влиянием внешних условий на генетически нормальный организм копируются признаки совсем другого генотипа. Проявление дальтонизма может произойти под влиянием питания, плохой психической конституции, повышенной раздражительности. У человека возникает заболевание витилиго (1% людей) – нарушение пигментации кожи. Фенокопии появляются в большинстве случаев при действии внешней среды на ранних стадиях эмбриогенеза, что приводит к врожденным заболеваниями порокам развития. Наличие фенокопий затрудняет диагностику заболеваний.
- модификационная - отражает изменение фенотипа под воздействием факторов внешней среды (усиление и развитие мышечной и костной массы у спортсменов, увеличение эритропоэза в условиях высокогорья и крайнего севера).
-Случайная
Наследственная:
генотипическая изменчивость распространяется на генетический аппарат. Подразделяется на мутационную и комбинативную.
1) мутационная.
Мутации - это редкие, случайно возникшие стойкие изменения генотипа, затрагивающие весь геном, целые хромосомы, части хромосом. Они возникают под действием мутагенных факторов физического, химического или биологического происхождения.
Мутации бывают:
∙спонтанные - это мутации, возникшие ненаправленно, под действием неизвестного мутагена
∙индуцированные - мутации, вызванные искусственно действием известного мутагена.
∙вредные, полезные и нейтральные;
Мутационный процесс повышает генетическое разнообразие популяций, что создает предпосылки для эволюционного процесса.
2) комбинативная
Результат независимого расхождения хромосом в процессе мейоза, оплодотворения, кроссинговера с рекомбинацией генов. Связана с новым сочетанием неизменных генов родителей в генотипах потомства.. При комбинативной изменчивости происходит перекомбинация генов, возникает новый индивидуальный набор хромосом, а значит, новый генотип и фенотип.
Факторы комбинативной изменчивости.
1.Независимое и случайное расхождение гомологичных хромосом в анафазе I мейоза.
2.Кроссинговер.
3.Случайное сочетание гамет при оплодотворении.
4.Случайный подбор родительских организмов.
Геномная мутация - это изменение числа хромосом. Виды геномных мутаций:
1.Полиплоидия — изменение числа гаплоидных наборов хромосом в кариотипе. Под кариотипом понимают число, форму и количество хромосом, характерные для данного вида. Различают нуллисомию (отсутствие двух гомологичных хромосом), моносомию (отсутствие одной из гомологичных хромосом) и полисомию (наличие двух и более лишних хромосом)
2.Гетероплоидия — изменение числа отдельных хромосом в кариотипе.
Генные мутации встречаются наиболее часто. Причины генных мутаций:
∙выпадение нуклеотида;
∙вставка лишнего нуклеотида (эта и предыдущая причины приводят к сдвигу рамки считывания);
∙замена одного нуклеотида на другой.
Только с генными мутациями связано появление новой, ранее не существующей в природе биоинформации.
Хромосомные мутации — это изменения структуры хромосом и процессе клеточного деления. Хромосомные и геномные мутации в функционально генетическом отношении сводятся либо к изменению количества биоинформации (делеция - потеря участка хромосомы, дупликации участков хромосом-удвоение участка хромосомы за счет неравного кроссинговера , гаплойдные, диплойдные клетки), либо к перекомбинфции блоков биоинформации разного объема (транслокацииперемещение участка хромосомы на другую хромосому, транспозиции, инверсии - поворот участка хромосомы на 180°, инсерции).
Классификация по эволюционному значению:
Групповая, реализованная группой организмов, и индивидуальная изменчивость – у одного организма или группы его клеток.
4 . Формы взаимодействия аллельных генов. Плейотропное действие гена. Множественный аллелизм.
Гены, от которых зависит развитие альтернативных признаков, принято называть алельными. Алельные гены расположены в гомологичных хромосомах в одних и тех же локусах. Доминирование — это такое взаимодействие аллельных генов, при котором проявление одного из аллелей (А) не зависит от присутствия в генотипе другого аллеля (А') и гетерозиготы АА' фенотипически не отличаются от гомозигот по этому аллелю (АА). Такая ситуация наблюдается, например, когда один из аллелей гена А («дикий») способен обеспечить формирование определенного варианта признака (синтез пептида с определенными свойствами), а другой А'—не обладает такой способностью. Наличие в генотипе АА' единственного нормального аллеля А приводит к формированию нормального признака. Этот аллель выступает как доминантный в данном гетерозиготном генотипе. Присутствие другого аллеля (А') фенотипически не проявляется, поэтому его называют рецессивным.
Полное доминирование — взаимодействие двух аллелей одного гена, когда доминантный аллель полностью исключает проявление действия второго аллеля. В фенотипе присутствует только признак, задаваемый доминантной аллелью.
Неполное доминирование — доминантный аллель в гетерозиготном состоянии не полностью подавляет действие рецессивного аллеля. Гетерозиготы имеют промежуточный характер признака. Неполное доминирование наблюдается, когда фенотип гетерозигот BB' отличается от фенотипа гомозигот по обоим аллелям (BB или B'B') промежуточным проявлением признака. Это объясняется тем, что аллель, способный сформировать нормальный признак, находясь в двойной дозе у гомозиготы BB, проявляется сильнее, чем в единственной дозе у гетерозиготы BB'. Указанные генотипы отличаются экспрессивностью, т.е. степенью выраженности признака. Демонстрацией такого типа взаимодействия генов могут быть многочисленные наследственные заболевания у человека, проявляющиеся клинически у гетерозигот по мутантным аллелям, а у гомозигот заканчивающиеся смертью. Иногда гетерозиготы имеют почти нормальный фенотип, а гомозиготы характеризуются пониженной жизнеспособностью.Так, гомозиготы по аллелю серповидноклеточности эритроцитов в связи с развитием у них тяжелой формы анемии и других фенотипических проявлений обычно не переживают детский возраст. Напротив, гетерозиготы — это как правило нормальные люди. Вместе с тем, они все-таки испытывают кислородную недостаточность в большей степени в сравнении с гомозиготами по аллелю дикого типа, в частности при подъеме на высоту Сверхдоминирование — более сильное проявление признака у гетерозиготной особи, чем у любой гомозиготной.
Кодоминирование представляет собой такой тип взаимодействия аллельных генов, при котором каждый из аллелей проявляет свое действие. В результате этого формируется некий промежуточный вариант признака, новый по сравнению с вариантами, определяемыми каждым аллелем самостоятельно. Примером может служить формирование IV, или АВ-группы, крови у человека, гетерозиготного по аллелям IA и IB, которые по отдельности детерминируют образование II и III групп крови.Кодоминирование — проявление у гибридов нового признака, обусловленного взаимодействием двух разных аллелей одного гена. Фенотип гетерозигот не является чем-то промежуточным между фенотипами разных гомозигот.
Межаллельная комплементация относится к достаточно редко встречаемым способам взаимодействия аллельных генов. В этом случае возможно формирование нормального признака D у организма, гетерозиготного по двум мутантным аллелям гена D(D'D"). Допустим, что ген D отвечает за синтез какого-то белка, который имеет четвертичную структуру, состоящую из нескольких одинаковых пептидных цепей. Мутантный аллель D' определяет синтез измененного пептида D', a мутантный аллель D" приводит к синтезу другой, но тоже измененной структуры пептида D". Можно представить ситуацию, когда взаимодействие таких измененных пептидов (D' и D") при формировании четвертичной структуры, как бы взаимно компенсируя эти изменения, обеспечивает образование белка с нормальными свойствами. В то же время отдельно взаимодействующие пептиды D' или D" формируют аномальные белки. Таким образом, с определенной вероятностью у гетерозигот D'D" в результате межаллельной комплементации может образовываться нормальный признак в виде белка с нормальными свойствами.
Аллельное исключение—такой вид взаимодействия аллельных генов в генотипе организма, который можно понять на примере рассмотренного выше механизма инактивации одной из Х-хромосом у особей гомогаметного пола, приводящего в соответствие дозы Х-генов у всех представителей вида. Инактива-ция одного из аллелей в составе Х-хромосомы способствует тому, что в разных клетках организма, мозаичных по функционирующей хромосоме, фенотипически проявляются разные аллели (рис. 3.79). Аллельное исключение наблюдается также в В-лимфоцитах, синтезирующих специфичные антитела к определенным антигенам. Моноспецифичность таких иммуноглобулинов требует выбора, который должна осуществить каждая клетка между экспрессией отцовского или материнского аллеля.
Множественный аллелизм — различные состояния (три и более) одного и того же локуса хромосом, возникшие в результате мутаций.
Присутствие в генофонде вида одновременно различных аллелей гена называют множественным аллелизмом. Примером этому служат разные варианты окраски глаз у плодовой мухи: белая, вишневая, красная, абрикосовая, эозиновая,— обусловленные различными аллелями соответствующего гена. У человека, как и у других представителей органического мира, множественный аллелизм свойствен многим генам. Так, три аллеля гена I определяют групповую принадлежность крови по системе АВ0 (IA, IB, I0). Два аллеля имеет ген, обусловливающий резуспринадлежность. Более ста аллелей насчитывают гены α- и β-полипептидов гемоглобина.Причиной множественного аллелизма являются случайные изменения структуры гена (мутации), сохраняемые в процессе естественного отбора в генофонде популяции. Многообразие аллелей, рекомбинирующихся при половом размножении, определяет степень генотипического разнообразия среди представителей данного вида, что имеет большое эволюционное значение, повышая жизнеспособность популяций в меняющихся условиях их существования. Кроме эволюционного и экологического значения аллельное состояние генов оказывает большое влияние на функционирование генетического материала. В диплоидных соматических клетках эукариотических организмов большинство генов представлено двумя аллелями, которые совместно влияют на формирование признаков. .
У человека серией множественных аллелей представлен ген, определяющий группу крови. При этом гены, обусловливающие группы крови А и В, не являются доминантными по отношению друг к другу и оба доминантны по отношению к гену, определяющему группу крови О. Следует помнить, что в генотипе диплоидных организмов могут находиться только два гена из серии аллелей. Остальные аллели данного гена в разных сочетаниях входят в генотип других особей данного вида. Таким образом, множественный аллелизм характеризует разнообразие генофонда целого вида, т. е. является видовым, а не индивидуальным признаком.
5 . Взаимодействие неаллельных генов, их виды.
Неаллельные гены, т.е. располагающиеся на разных хромосомах или на одной хромосоме, но в разных локусах.
Комплементарность.
Если формирование сложного признака требует совокупного действия(взаимодействия) аллелей конкретных неаллельных генов, то очевидно, он будет проявляться в фенотипе лишь тех особей, которые в генотипах имеют именно требуемую комбинацию аллелей. По существу речь идет о том что в результате совокупного действия возникает новое качество , которое не было при отсутствии соответствующего взаимодействия. Комплементарность – явление, при котором 2 неаллельных гена. Находясь в генотипе, одновременно приводят к формированию нового признака. Если присутствует один из пары – проявляется он.
Примером служат группы крови у человека, процесс формирования половой принадлежности у людей(например синдром Морриса). Комплементарность может быть доминантная и рецессивная. Для того чтобы человек имел нормальный слух, необходимо чтобы работали, согласовано многие гены, и доминантные и рецессивные. Если, хотя бы по одному гену он будет гомозиготен по рецессиву – слух будет ослаблен. Комплементарность — взаимодействие неаллельных генов, которые обусловливают развитие нового признака, отсутствующего у родителей.
Эпистаз – такое взаимодействие генов, когда ген одной аллельной пары маскируется действием другой аллельной пары. Это обусловлено тем, что ферменты катализируют разные процессы клетки, Когда на одном метаболическом пути действуют несколько генов. Действие их должно быть согласовано во времени.
Подавляющий ген называется геном-супрессором, а подавляемый - гипостатическим геном. Повидимому, действие гена-супрессора на подавляемый ген сходно с принципом доминантность - рецессивность. Но существенное различие заключается в том, что эти гены не являются аллельными, т. е. расположены в негомологичных хромосомах или занимают различные локусы в гомологичных. Различают доминантный рецессивный эпистаз. При доминантном эпистазе доминантный аллель гена-супрессора подавляет проявление доминантного аллеля другого гипостатического гена. При рецессивном эпистазе, или криптомерии, рецессивный аллель. Пример рецессивного эпистаза: «бомбейский феномен», случаи альбнизма у людей .Изменяющейся эпистаз. У кроликов и некоторых других животных известен рецессивный ген гималайской («горностаевой») окраски, обусловливающей своеобразную пятнистость меха (на белом или светлом фоне кончики лап, ушей, морды и хвоста имеют чёрную окраску). Однако такая окраска развивается только при выращивании молодняка гималайской породы при умеренных температурах. При повышенной температуре весь мех у особей того же гималайского генотипа оказывается белым, а при пониженной — чёрным. Этот пример указывает на то, что на эпистаз влияют факторы внешней среды, в данном случае температуры.
Полимерия. Если гены располагаются, каждый в своем отдельном локусе, но их взаимодействие проявляется в одном и том же направлении – это полигены. Один ген проявляет признак незначительно. Полигены дополняют друг друга и оказывают мощное действие – возникает полигенная система – т. е. система является результатом действия одинаково направленных генов. Гены подвергаются значительному влиянию главных генов, которых более 50. Полигенных систем известно множество.
Развитие колличественных признаков нередко обусловлено полимерными генами или полигенамисистемой неаллельных генов,в равной мере влияющих на формирование соответствующего признака и характеризующих суммирующим действием. Соответственно степень выраженности признака зависит от совокупной дозы доминантных аллелей во всех полимерных генов системы. В потомстве гетерозигот наблюдается разная степень выраженности признака, зависящая от суммарной дозы соответствующих аллелей. Например, степень пигментации кожи у человека, определяемая четырьмя парами генов, колеблется от максимально выраженной у гомозигот по доминантным аллелям во всех четырех парах (Р1Р1Р2Р2Р3Р3Р4Р4) до минимальной у гомозигот по рецессивным аллелям (р1р1р2р2р3р3р4р4)В разобранном примере доминантные аллели полигенов определяют синтез пигмента, а рецессивные — практически не обеспечивают этого признака. В клетках кожи организмов, гомозиготных по рецессивным аллелям всех генов, содержится минимальное количество гранул пигмента.По этому типу у людей наследуется интенсивность окраски кожных покровов, рост, уровень интеллекта и др.
В некоторых случаях доминантные и рецессивные аллели полигенов могут обеспечивать развитие разных вариантов признаков. Например, у растения пастушьей сумки два гена одинаково влияют на определение формы стручочка. Их доминантные аллели образуют одну, а рецессивные — другую форму стручочков Модифицирующее действие. Гены модификаторы сами по себе не определяют какой - то признак, но
могут усиливать или ослаблять действие основных генов, вызывая таким образом изменение фенотипа. В качестве примера обычно приводится наследование пегости у собак и лошадей. Числового расщепления никогда не даётся, так как характер наследования больше напоминает полигенное наследование количественных признаков. 1919 год Бриджес ввел термин генмодификатор. Теоретически любой ген может взаимодействовать с другими генами, а значит, и проявлять модифицирующее действие, но некоторые гены – модификаторы в большей степени. Они часто не имеют собственного признака, но способны усиливать или ослаблять проявление признака, контролируемого другим геном. В формировании признака кроме основных генов проявляют свое действие и модифицирующие гены.
6 . Закономерности наследования признаков по Г.Менделю. Менделирующие признаки у человека. ОСНОВНЫЕ ЗАКОНОМЕРНОСТИ НАСЛЕДОВАНИЯ Основные закономерности наследования были открыты Г.Менделем. По уровню развития науки
своего времени Мендель не мог еще связать наследственные факторы с определенными структурами клетки. В настоящее же время мы знаем, что гены находятся в хромосомах, поэтому при объяснении закономерностей Менделя мы будем исходить из современных представлений на клеточном уровне. Он достиг успеха в своих экспериментах благодаря использованию гибридологического метода
— скрещивания организмов, различающихся по каким-либо признакам, и анализа всех последующих поколений с целью установления закономерностей наследования этих признаков. Гибридологический метод и до настоящего времени остается одним из основных в генетических исследованиях.
Объектом для исследования Мендель избрал горох, имеющий много рас, отличающихся альтернативными признаками.
Скрещивание, в котором родительские особи различаются по одному признаку, называется моногибридным, по двум признакам - дигибридным, по многим признакам — полигибридным. МОНОГИБРИДНОЕ СКРЕЩИВАНИЕ 1 правило Менделяправило единообразия гибридов первого поколения: при скрещивании
гомозиготных особей, отличающихся друг от друга по одной паре альтернативных (взаимно исключающих) признаков, все потомство в первом поколении единообразно как по фенотипу, так и по генотипу.
ПРАВИЛО РАСЩЕПЛЕНИЯ 2 правило Менделя: при скрещивании 2 гетерозиготных особей, отличающихся одной парой
альтернативных признаков, в потомстве происходит расщепление в отношении 3:1 по фенотипу и 1:2:1 по генотипу. Согласно 2 правилу можно заключить:
∙аллельные гены, находясь в гетерозиготном состоянии, не изменяют друг друга;
∙при созревании гамет у гибридов образуется приблизительно равное число гамет с доминантным и рецессивным геном;
∙при оплодотворении мужские и женские гаметы, несущие доминантные и рецессивные гены, свободно комбинируются.
НЕПОЛНОЕ ДОМИНИРОВАНИЕ Свойством неполного доминирования обладает ряд генов, вызывающих наследственные аномалии и
болезни человека. Так, например, наследуется серповидноклеточная анемия и атаксия Фредриха, характеризуемая прогрессирующей потерей координации произвольных движений. ПОЛИГИБРИДНОЕ СКРЕЩИВАНИЕ
Дигибридное скрещивание как пример полигибридного скрещивания.
При полигибридном скрещивании родительские организмы отличаются по нескольким признакам. Примером такого скрещивания может служить дигибридное, при котором родительские организмы отличаются по двум признакам.
Правило независимого комбинирования
3 правило: скрещивание гомозиготных особей, отличающихся двумя (или более) парами альтернативных признаков - во 2 поколении отмечается независимое комбинирование признаков, в результате чего появляются гибридные формы, несущие признаки в сочетаниях, не свойственных родительским особям.
ВЗАИМОДЕЙСТВИЕ ГЕНОВ Развитие любых признаков у организмов является следствием сложных взаимодействий между генами.
Взаимодействие между генами:
∙ из одной аллели: неполное доминирование , доминирование, сверхдоминирование из различных аллелей: комплементарное действие, эпистаз, полимерия
7 . Типы наследования признаков, их характеристика. Экспрессивность и пенетрантность.
Степень выраженности признака в фенотипе получило название экспрессивности. Таким образом, под экспрессивностью понимают выраженность фенотипического проявления гена. Она связана с изменчивостью признака в пределах нормы реакции. Экспрессивность также является показателем, характеризующим фенотипическое проявление наследственной информации. Она характеризует степень выраженности признака и, с одной стороны, зависит от дозы соответствующего аллеля гена при моногенном наследовании или от суммарной дозы доминантных аллелей генов при полигенном наследовании, а с другой — от факторов среды. Примером служит интенсивность красной окраски цветков ночной красавицы, убывающая в ряду генотипов АА, Аа, аа, или интенсивность пигментации кожи у человека, увеличивающаяся при возрастании числа доминантных аллелей в системе полигенов от 0 до 8. Влияние средовых факторов на экспрессивность признака демонстрируется усилением степени пигментации кожи у человека при ультрафиолетовом облучении, когда появляется загар, или увеличением густоты шерсти у некоторых животных в зависимости от изменения температурного режима в разные сезоны года.
Один и тот же признак может проявляться у некоторых организмов и отсутствовать у других, имеющих тот же ген. Частота встречаемости данного признака в поколении называется пенетрантность.
Экспресивность и пенетрантность поддерживаются естественным отбором. Обе закономерности необходимо иметь в виду при изучении наследственности у человека. Следует помнить, что гены, контролирующие патологические признаки, могут иметь различную экспрессивность и пенетрантность. Изменяя условия среды, можно влиять на экспрессивность, пенетрантность и доминирование.
Явление экспрессивности указывает, что доминированием можно управлять.
Основные типы наследования:
Прямое наследование, при котором варианты признаков сохраняются в неизменном виде из поколения в поколение – это самый простой тип наследования признаков. Прямое наследование часто наблюдается у растений, которые размножаются вегетативным путем или образуют семена при самоопылении, реже – при размножении животных (в пределах одной породы) или перекрестном опылении у растений (в пределах одного сорта или линии).
Непрямое наследование – это более сложный тип наследования, который наблюдается при размножении животных и семенном размножении у растений (которое по сути также является половым). Для изучения непрямого наследования необходима гибридизация – скрещивание организмов, различающихся по генотипу. При непрямом наследовании некоторые варианты признаков проявляются в каждом поколении (такие признаки называются доминантными, «господствующими»), а другие варианты могут временно «исчезать», а затем проявляться в последующих поколениях (такие признаки называются рецессивными, «отступающими»).
Сложные типы наследования признаков потому и называются сложными, что заранее предсказать появление новых вариантов признаков очень трудно. В некоторых случаях «внезапно» возникают новые варианты признаков, которых не было ни у родителей, ни у бабушек–дедушек, ни у тетушек– дядюшек. Иногда такое «внезапное» появление признаков совершенно необоснованно называют мутацией. Пример. Аквариумные рыбки меченосцы (и близкая к меченосцам группа – пецилии) характеризуются разнообразной окраской: зеленовато-серой, темно-красной (кирпичной), яркокрасной (алой), лимонной (светло-желтой), пятнистой (тигровой и ситцевой)
8. Понятие "сцепление" генов. Х-сцепленное наследование признаков у человека.
Явление, в основе которого лежит локализация генов в одной хромосоме. Сцепление генов впервые обнаружено в 1906 У. Бэтсоном и Р. Пеннетом в опытах по скрещиванию душистого горошка. Позднее сцепление генов было детально исследовано Т. Морганом с сотрудниками в экспериментах с дрозофилой. Сцепление генов выражается в том, что аллели сцепленных генов, находящиеся в одной группе сцепления, имеют тенденцию наследоваться совместно. Это приводит к образованию у гибрида гамет преим. с «родительскими» сочетаниями аллелей. Для обозначения сцепления генов используют символы АВ/ав или АВ/Ab сцепление доминантных (или рецессивных) аллелей друг с другом АВ/ав наз. фазой сцепления, а сцепление доминантных аллелей с рецессивными Ав/аВ — фазой отталкивания. В обоих случаях сцепление генов приводит к более низкой частоте особей с «неродительскими», рекомбинантными сочетаниями признаков, чем ожидается при независимом наследовании признаков. При полном сцеплении генов образуются только два типа гамет (с исходными сочетаниями сцепленных генов), при неполном — и новые комбинации аллелей сцепленных генов. Неполное сцепление генов— результат кроссинговера между сцепленными генами, поэтому полное сцепление генов возможно у организмов, в клетках которых кроссинговер в норме не происходит (напр., половые клетки самцов дрозофилы). Т. о., полное сцепление генов является скорее исключением из правила неполного сцепления генов. Кроме того, полное сцепление генов может имитироваться явлением плейотропии. В некоторых случаях в мейозе регулярно происходит неслучайное расхождение негомологичных хромосом к одному полюсу, что приводит к образованию гамет преим. с определенными сочетаниями аллелей несцепленных между собой генов. Разные пары генов в пределах одной группы сцепления характеризуются различной степенью сцепления в зависимости от расстояния между ними. Чем больше расстояние между генами в хромосоме, тем меньше сила сцепления между ними и чаще образуются рекомбинантные типы гамет. Изучение сцепления генов и сцепленного наследования признаков послужило одним из подтверждений хромосомной теории наследственности и исходным толчком анализа и разработки теории кроссинговера.
Х-сцепленное наследование
Так как Х-хромосома присутствует в кариотипе каждого человека, то и признаки, наследуемые сцеплено с Х-хромосомой, проявляются у представителей обоих полов. Женщины получают эти гены от обоих родителей и через свои гаметы передают их потомкам. Мужчины получают Х-хромосому от матери и передают ее своему потомству женского пола.
Различают Х-сцепленное доминантное и Х-сцепленное рецессивное наследование. У человека Х- сцепленный доминантный признак передается матерью всему потомству. Мужчина передает свой Х- сцепленный доминантный признак лишь своим дочерям. Х-сцепленный рецессивный признак у женщин проявляется лишь при получении ими соответствующего аллеля от обоих родителей. У мужчин он развивается при получении рецессивного аллеля от матери. Женщины передают рецессивный аллель потомкам обоих полов, а мужчины — только дочерям.
При Х-сцепленном наследовании возможен промежуточный характер проявления признака у гетерозигот.
Y-сцепленные гены присутствуют в генотипе только мужчин и передаются из поколения в поколение от отца к сыну.
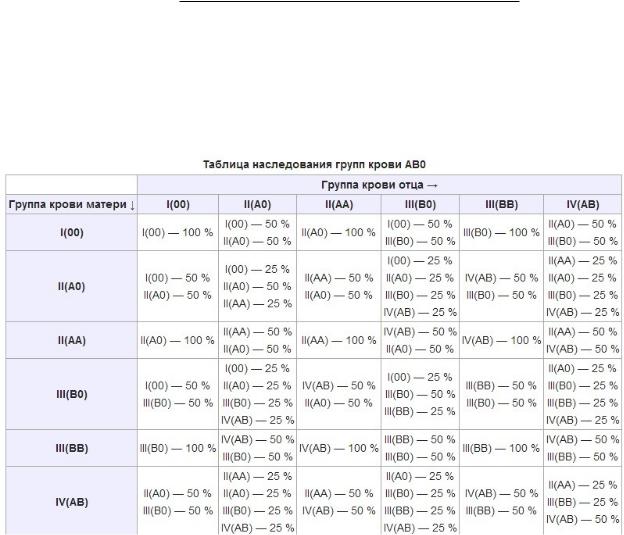
9. Наследование групп крови системы AB0 у человека
Основная система классификации крови -- система ABO (читается - а, б, ноль)
Гpуппы кpoви обозначают по наличию или отсутствию определенного типа «склеивающего» фактора (агглютиногена):
0 (I) — 1-я группа крови. А (II) — 2-я.
В (III) — 3-я
АВ (IV) — 4-я группа крови.