

5.5. Фауна мелких млекопитающих
Для изучения плейстоценового сообщества мелких млекопитающих использованы фаунистические материалы двух участков:
- раскоп 1994 г. - квадраты Е/6-8 (линия субквадратов а - в), слои 9-22 общей мощностью 4 м;
- раскоп 1995 г. - квадраты Е/5 (субквадраты а - г), Е/6-8 (линия субквадратов б - г), слои 9-22 общей мощностью 4,5 м.
Результаты определения собранных материалов и заключения по ним приведены для каждой коллекции отдельно, что обусловлено рядом причин. Отложения Денисовой пещеры отличаются сложным пространственным залеганием. Ряд слоев имеет линзовидный характер, а некоторые слои перемяты и смещены за счет конвективной неустойчивости грунтов. От квадрата к квадрату меняется мощность слоев, нарушается их последовательность за счет выклинивания некоторых горизонтов. Также неравномерна концентрация костного материала по площади. Эти обстоятельства придают высокую тафономическую индивидуальность отдельным участкам раскопа. Фактически каждый квадрат неповторим по условиям осадконакопления и набору костных остатков млекопитающих. Учитывая сложную тафономическую ситуацию, необходима дифференцированная оценка полученных результатов. Такой подход позволяет отделить случайные флуктуации таксономического разнообразия от истинных изменений состава фауны, отражающих динамику развития природной среды.
Общий список таксонов мелких позвоночных, установленный для плейстоценовых отложений Денисовой пещеры, включает более 40 названий (табл. 3).
В коллекции 1994 г. количество определимых костных остатков достигает 7 тыс. экз. При оценке относительного обилия различных таксонов было использовано 6 326 экз. В среднем на каждый слой приходится приблизительно 200 определимых остатков. В целом нижняя часть разреза охарактеризована более надежно, чем верхняя. Характер распределения костных остатков по стратиграфическим подразделениям разреза представлен на диаграммах общего (рис. 46) и выборочного (рис. 47) состава мелких животных. За время накопления толщи плейстоценовых отложений видовой состав млекопитающих менялся незначительно. Этот процесс отражает сходный механизм формирования танатоценоза пещеры и сходный состав населения мелких млекопитающих в долине Ануя в период накопления толщи слоев 22-9.
На фоне относительной стабильности состава фауны от нижних слоев к верхним обращает на себя внимание резкое уменьшение количества останков летучих мышей Chiroptera в отложениях слоя 21. Выше слоя 19 их количество сокращается приблизительно в десять раз и на этом уровне сохраняется до кровли плейстоценовых отложений. Объяснение этого феномена с палеоэкологических позиций найти трудно. Судя по составу мелких млекопитающих, от слоя 22 вверх по разрезу внешние природные факторы не претерпели существенных изменений. Видимо, единственной причиной, нарушившей условия обитания летучих мышей в пещере, был человек. Можно предполагать, что, начиная с эпохи накопления слоя 21, присутствие человека в пещере стало постоянным, а использование огня -регулярным. Это предположение подтверждается та-фономическими наблюдениями. Первые хорошо выраженные прослойки углей появляются в отложениях слоя 21. Начиная со слоя 20, резко возрастает количество артефактов палеолитического человека. Эти данные свидетельствуют о стабильном присутствии человека и задымлении полости пещеры, что привело к увеличению фактора беспокойства и к резкому сокращению численности летучих мышей в пещере.
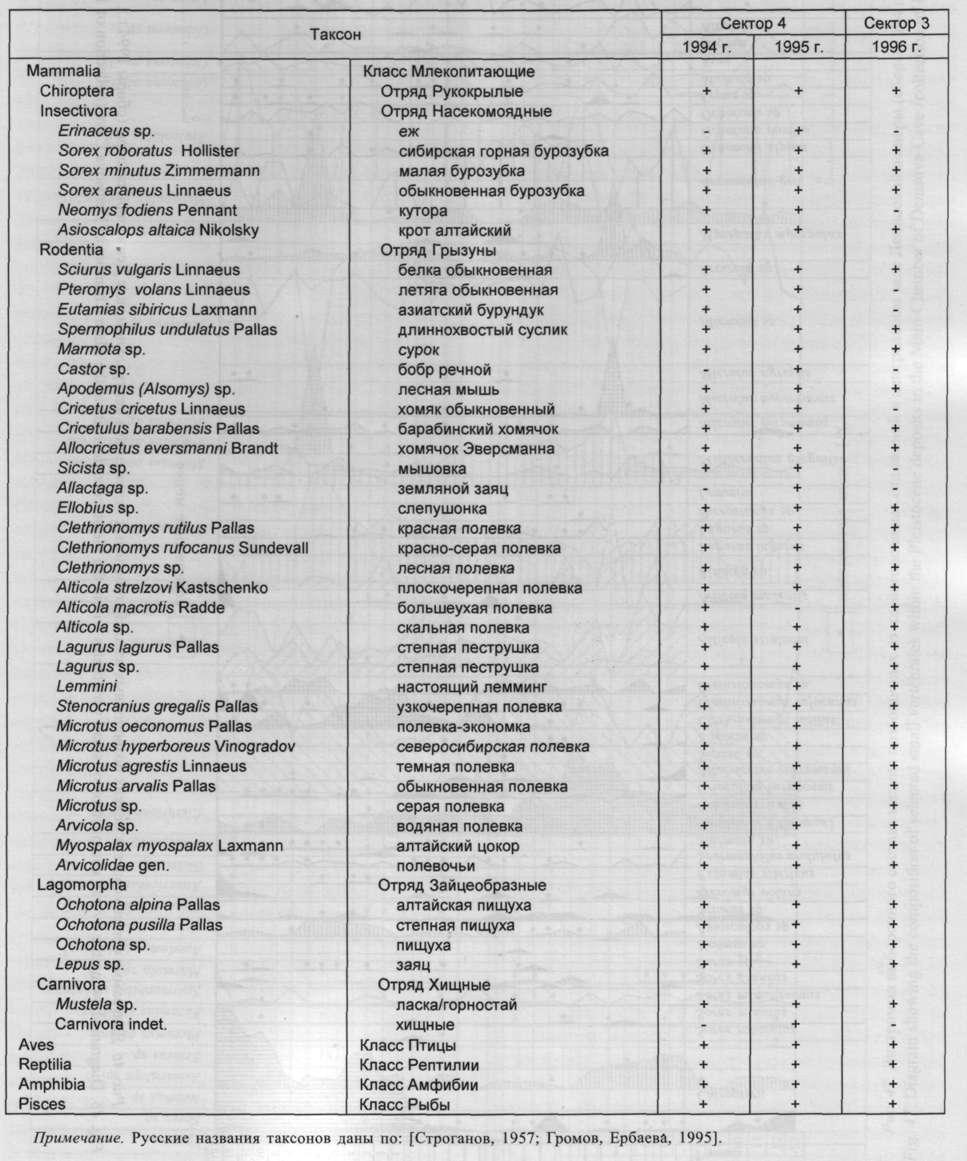
Таблица 3. Общий состав мелких позвоночных из плейстоценовых отложений в Денисовой пещере

Рис. 46. Диаграмма общего состава мелких позвоночных из плейстоценовых отложений в центральном зале Денисовой пещеры (сборы 1994 г.).
 Диаграмма
выборочного состава мелких позвоночных
из плейстоценовых отложений в центральном
зале Денисовой пещеры
Диаграмма
выборочного состава мелких позвоночных
из плейстоценовых отложений в центральном
зале Денисовой пещеры
Остальная часть микротериофауны дает ясную картину единого сообщества, однако его структура постепенно менялась во времени. Причем состав этого сообщества принципиально отличался от современного, что свидетельствует об ином облике окружающих ландшафтов в плейстоцене.
Ведущую группу мелких млекопитающих на протяжении палеолитического периода составляли полевки Microtinae. В питании хищных птиц наибольшую численность имели азиатские горные полевки родаAlticola, среди последних доминировала плоскочерепная полевкаAlticola strelzovi, обитатель каменистых сухих склонов, покрытых редкой растительностью. Количественное соотношение этой полевки с другими видами достаточно стабильно и варьирует по горизонтам от 10 до 20%, несколько увеличиваясь в слоях 11-9 до 29%. Содоминантами в составе фауны являлись узкочерепная полевкаStenocranius gregalis и рыжие полевкиClethrionomys. В целом численность узкочерепной полевки несколько выше численности рыжей полевки. По-разному выглядит тенденция их динамики во времени. Численность узкочерепной полевки вверх по разрезу постепенно нарастает, а рыжей полевки -снижается. Среди останков рыжих полевок преобладают зубыClethrionomys rutilus, изредка встречаются зубыС. rufocanus.
В плейстоценовых отложениях присутствуют степные пеструшки рода Lagurus. Их останки впервые появляются на уровне 10 горизонта 22.2. Однако до слоя 21 обилие пеструшек не превышает 1% от общего количества останков. Начиная со слоя 20, их численность постепенно нарастает, давая максимум в слое 9-11% от общего числа останков. По своей морфологии ископаемые пеструшки из неоплейстоценовых отложений пещеры близки современнойLagurus lagurus.
Останки полевки-экономки Microtus oeconomus составляют также незначительную долю в выборке -от 0,5 до 3%. Однако они встречаются стабильно, практически во всех горизонтах. Постоянно отмечаются землеройки родаSorex, хотя обилие их останков не превышает 2-3%. Сходную картину дают сусликиSpermophilus undulatus, даурский хомячокCricetulus barabensis и хомяк ЭверсманнаAllocricetus eversmanni, останки которых отмечены во всех горизонтах. Однако ни те ни другие не найдены на уровне 13 горизонта 22.3. Кроме того, встречаемость суслика нарастает вверх по разрезу, достигая максимума в горизонте 9.1. Численность даурского хомячка стабильна в средней части разреза. Хомяк Эверсманна дает пик в горизонте 9.1.
Другая группа мелких млекопитающих, останки которых постоянно встречаются в плейстоценовых отложениях Денисовой пещеры, - зайцеобразные. Среди них ведущее место занимает пищуха Ochotona. Ее останки найдены практически во всех горизонтах и составляют от 1 до 6%. Причем наибольшее количество приходится на нижнюю часть горизонта 22.1 (уровни 3 - 5). Всплеск численности фиксируется также в горизонтах 11.2 и 9.2. Полученный материал лишь в некоторых случаях позволяет диагностировать виды. Отмечено присутствие двух групп: мелкой степной пищухиOchotona pus ilia и более крупнойО. alpina. Однако количество определимых до вида останков недостаточно, чтобы анализировать различия в их послойной приуроченности.
Регулярно в отложениях пещеры встречаются останки зайцев. Их относительное количество колеблется в пределах 0,15 - 3%. В небольшом количестве, но по всей толще пещерных отложений, встречаются останки мелких хищников, преимущественно куньих. Их основная часть принадлежит представителям рода Mustela.
Третью группу в списке мелких млекопитающих составляют виды, количество костных остатков которых относительно невелико, а встречаемость по слоям спорадична. К ним относится сурок Marmota, число останков которого колеблется в интервале 0,2 - 1,2%. Останки сурка зарегистрированы на уровнях 10, 8 - 5, 1 слоя 22, в слое 20 и в горизонтах 19.1,12.1 и 9.3. Их небольшое количество и плохая сохранность не позволяют диагностировать вид. Вероятно, в танатоценоз пещеры сурок попадал не в погадках хищных птиц, а с пометом млекопитающих (волка, гиены) или как охотничий трофей человека. По нашим наблюдениям, сурок был объектом охоты палеолитического человека на стоянке Каменная Балка в Приазовье. Известно, что сурок добывался охотниками некоторых палеолитических стоянок Франции [Patou, 1987]. Еще реже и в меньшем количестве встречается бурундукEutamias sibiricus. Он отмечен на уровнях 12,6,4 и 3 слоя 22 и в горизонте 19.1. В двух случаях отмечены белкаSciurus (уровни 6 и 4 слоя 22) и летягаPteromys volans (уровень 12 горизонта 22.3 и горизонт 13.1).
В общей динамике численности мелких млекопитающих некоторое своеобразие имеет слепушонка Ellobius. Впервые ее останки появляются в горизонте 19.1. Далее по разрезу они отмечены в горизонтах 13.1,12.3,9.3 и 9.1. В горизонте 12.3 их количество достигает 2,4%.
Редкими представителями тафоценоза плейстоценовых слоев пещеры являются серый лемминг, темная полевка и водяная полевка. Представители родаЬеттиз найдены на уровнях 12, 10, 9 слоя 22 и в горизонтах 13.4,13.1,12.3. Число их останков колеблется от 0,2 до 0,95%. В четырех случаях найдена темная полевкаMicrotus agrestis - уровни 12,11 горизонта 22.3, горизонты 19.1 и 13.4. Водяная полевкаArvicola, близкая по своим морфологическим признакам современнойArvicola terrestris, отмечена в слоях 20,19,17, а также в горизонтах 13.3. и 9.2. К наиболее редким представителям ископаемой фауны принадлежат копытный лемминг и лесная мышь. По одному фрагменту зубов копытного леммингаDicrostonyx отмечено в горизонтах 19.1 и 12.1. Мыши рода Apodemus найдены в нижней части слоя 22 (уровни 11 и 10).
Помимо мелких млекопитающих в формировании состава тафоценоза палеолитических слоев принимали участие останки птиц, рыб, отчасти лягушек и мелких рептилий. Число останков птиц варьирует по разным слоям от 0,5 до 9,3%. Позвонки и отдельные кости рыб найдены во всех горизонтах разреза. Их относительное количество колеблется от 6,5 до 23%, составляя в среднем 7 - 9%. Останки лягушек и рептилий встречаются в небольшом количестве и спорадично по отдельным горизонтам разреза.
В коллекции 1995 г. общий объем определимых костных остатков составил 27 663 экз. Наиболее репрезентативный материал получен из отложений горизонта 22.1, а также в интервале горизонтов 12.1-11.1. Характер распределения костных остатков по стратиграфическим уровням разреза 1995 г. представлен на диаграммах общего (рис. 48) и выборочного (рис. 49) состава мелких позвоночных. Видовой состав мало меняется вверх по разрезу. Единственная группа мелких млекопитающих, количество останков которой резко меняется на границе слоев 22 и 21, - летучие мыши. Остальные виды позвоночных не претерпели резких колебаний численности в период накопления всей толщи. В целом ископаемая фауна Денисовой пещеры принципиально отличается от современной. Она отражает значительно большее разнообразие биотопических условий и пестроту ландшафтов по сравнению с современностью. Это разнообразие было обусловлено заметным участием в сообществах степных и высокогорных элементов.
 Рис.
48. Диаграмма общего состава мелких
позвоночных из плейстоценовых отложений
в центральном зале Денисовой пещеры
(сборы 1995 г.).
Рис.
48. Диаграмма общего состава мелких
позвоночных из плейстоценовых отложений
в центральном зале Денисовой пещеры
(сборы 1995 г.).
 Рис.
49. Диаграмма выборочного состава
мелких позвоночных из плейстоценовых
отложений в центральном зале Денисовой
пещеры (сборы 1995 г.).1-
деформация, 2 - содержание менее 1%.
Рис.
49. Диаграмма выборочного состава
мелких позвоночных из плейстоценовых
отложений в центральном зале Денисовой
пещеры (сборы 1995 г.).1-
деформация, 2 - содержание менее 1%.
Несмотря на общую однородность сообществ мелких млекопитающих, благодаря большому объему материала, удалось наметить определенную динамику количественного соотношения отдельных таксонов. Анализ этих флюктуации позволил проследить общие закономерности изменения биоты за время накопления плейстоценовых осадков Денисовой пещеры.
Летучие мыши Chiroptera являются единственным компонентом тафоценоза, который резко меняет свою численность. В слое 22 их относительное количество колеблется от 14,5% на уровнеЗ до 63,8% на уровне 13. К слою 21 обилие летучих мышей снижается до 9,9%. Далее по разрезу оно варьирует в интервале 3 - 6,6% до слоя 17, а в верхней части разреза их численность составляет менее 3%. Эти данные подтверждают отмеченную ранее закономерность и придают ей более достоверный характер. Скорее всего, на позднем этапе накопления осадков слоя 22 произошло принципиальное изменение в характере присутствия в пещере первобытного человека. Менее понятны два минимума численности летучих мышей на уровнях 8 и 3 слоя 22. При этом падение численности от уровня 13 к уровню 8 и от уровня 6 к уровню 3 имеет закономерный характер, что должно быть связано с природными явлениями, а не с чисто тафономическими причинами. Падение численности летучих мышей на уровне 8 слоя 22 отмечено также в материалах 1994 г., что существенно повышает уровень их достоверности.
Все летучие мыши из плейстоценовых отложений Денисовой пещеры являются видами, которые используют пещеры в качестве убежища. Видимо, накопление их костных остатков происходило в результате естественной гибели зверьков из состава зимующих колоний. Небольшая часть материала имеет следы обработки желудочным соком и, следовательно, попала в состав тафоценоза из погадок хищных птиц. В ископаемой фауне летучих мышей Денисовой пещеры установлено семь видов: остроухая ночница Myotis aff.blythi Tomes, прудовая ночницаМ. dasycneme Boie, ночница БрандтаМ. aff.brandti Eversmann, водяная ночница М cf.daubentoni Kuhli, сибирский (большой) трубконосMurina leusogaster Milne-Edwards, бурый ушанPlecotus auritus Linnaeus, северный кожанокEptesicus cf.nilssoni Keyserling, Blasius.
Максимальное число определимых останков Chiroptera приходится на слой 22. В этом же слое наблюдается максимальное видовое разнообразие летучих мышей. Здесь найдены останки всех рукокрылых, определенных для Денисовой пещеры. В отложениях горизонта 22.3 не найдена только Myotis cf.daubentoni, останки которой обнаружены в горизонте 22.1. Максимальное видовое разнообразие рукокрылых отмечено на уровне 6 горизонта 22.2. Здесь представлены пять видов летучих мышей; отсутствуют только остроухая ночницаМ. aff.blythi и водяная ночницаМ. cf.daubentoni. Наименьшее число видов отмечено на уровне 8 горизонта 22.2:Plecotus aff.auritus к Eptesicus cf.nilssoni. На этот же уровень приходится минимальная абсолютная численность останков Chiroptera. Наиболее многочисленны в слое 22 останки ушанаPlecotus aff.auritus - около 60% от всего определенного материала по этому слою.Eptesicus cf.Nilssoni иMyotis aff.dasycneme принадлежит по 12% останков. Костные остаткиMurina aff.leucogaster иMyotis aff.brandti составляют 3,5% . На долюMyotis aff.blythi ji M. cf.daubentoni приходится около 1% определенного по этому слою материала.
На рубеже слоев 22 и 21 резко падает абсолютное количество и видовое разнообразие останков рукокрылых. В слое 21 обнаружено только два вида летучих мышей - бурый ушан (Plecotus aff.auritus) и северный кожанок(Eptesicus cf.nilssoni). В отложениях слоев 19 и 17 отмечено небольшое количество останковPlecotus aff.auritus, Myotis blythi, Plecotus aff.auritus иМ. aff.dasycneme. Верхняя пачка плейстоценовой толщи (слои 13-9) также бедна ископаемыми рукокрылыми. Здесь найдены останки четырех видов:Myotis aff. blythi,M. aff.dasycneme, Eptesicus cf.nilssoni иPlecotus aff.auritus.
Следует отметить, что для значительной части ископаемых форм летучих мышей Денисовой пещеры использовалась открытая номенклатура. Это связано с тем, что по своей морфологии они не полностью идентичны аналогичным современным видам. Например, зубы плейстоценового северного кожанка Eptesicus cf.nilssoni и ночницы БрандтаMyotis aff.brandti значительно крупнее близких им современных форм.
Большинство ископаемых летучих мышей Денисовой пещеры в своей экологии связаны с широколиственными ил» хвойно-широколиственными лесами. Например, ночница Брандта населяет смешанные и широколиственные леса, по поймам проникает в тайгу и степь. Она распространена от Англии и востока Испании через Урал и Южную Сибирь до Кореи и Японии. Сибирский трубконос - обитатель смешанных хврйно-широколиственных и широколиственных (преимущественно горных) лесов от Алтая до Гималаев, Юго-Восточного Китая и Дальнего Востока. Северный кожанок рпсмроетраттеп п лесттой части Сеперпой Евразии: от Франции, Швейцарии и Кавказа до Гималаев, Тибета, Монголии и Дальнего Востока. На севере его ареал совпадает с границей лесной зоны. Водяная ночница является обитателем речных пойм лесной и лесостепной зон Европы, Южной Сибири; ее ареал простирается до Китая и Японии. Лишь остроухая ночница заселяет не только лесные, но и аридные ландшафты от Средиземноморья, через Кавказ, до Центральной Азии.
Еж Erinaceus sp. отмечен в плейстоценовой толще только в отложениях горизонта 14.2.
Землеройки Sorex sp. обычны в плейстоценовых отложениях пещеры, но ни в одном слое их количество не превышает 3%. Небольшое увеличение численности этих насекомоядных отмечено на уровнях 10, 8, 6 горизонта 22.2 и в слое 12. Подавляющая часть останков землероек принадлежитSorex araneus. Изредка встречается мелкаяS. minutus, еще реже - крупная 5.roboratus. Увеличение видового разнообразия землероек отмечено в отложениях слоя 12.
Кутора Neomys sp. очень редка в отложениях пещеры. Ее единичные фрагменты найдены в горизонтах 14.2 и 12.3. Скорее всего, это связано с тем, что кутора обитает в пойменных, хорошо защищенных биотопах.
Алтайский крот Asioscalops sp. найден практически во всех горизонтах плейстоценовой толщи. Максимальное число его останков приходится на слой 20-5,2%.
Белка Sciurus sp. очень редкий вид в отложениях пещеры. Ее единичные находки отмечены на уровне 5 горизонта 22.1 и в осадках горизонта 11.3.
Летяга Pteromys sp. также редкий вид. Ее останки обнаружены на уровнях 8,6 и 5 слоя 22, а также в отложениях горизонтов 19.1,14.2и 11.1.
Бурундук Eutamias sibiricus отмечен только на уровне 8 горизонта 22.2 и в отложениях слоя 21.
Длиннохвостый суслик Spermophilus undulatus обычен для всей толщи плейстоценовых отложений. Однако его относительная численность нигде не превышает 3%. Вероятно, это связано прежде всего с механизмом формирования тафоценоза: как объект охоты для птиц, гнездящихся в пещере, суслик был слишком велик.
Сурок Marmota sp. встречается с редкими перерывами почти по всему разрезу, за исключением нижней части слоя 22 (уровни 13-8).
Бобр Castor sp. известен только по сколам щечных зубов и эмали резцов, которые зарегистрированы в отложениях горизонтов 21.2 и 14.3.
Лесные мыши Apodemus (Alsomys) sp. определены по отдельным находкам в отложениях горизонтов 19.2. и 9.3
Хомяк обыкновенный Cricetus cricetus отмечен единичными останками на уровнях 5, 4, 1 горизонта 22.1 и в отложениях горизонтов 19.3,19.1. Эти материалы существенно дополняют сборы 1994 г., в которых хомяк был обнаружен только на уровне 3 горизонта 22.1.
Барабинский хомячок Cricetulus barabensis встречается по всей толще плейстоценовых отложений, хотя его относительная численность нигде не превышает 2 - 2,5%. В целом в нижних слоях он встречается чаще, а его количество несколько выше, чем в верхней части разреза.
Хомячок Эверсманна Allocricetus eversmanni также встречается почти во всех слоях разреза, но его количество не превышает 1,5- 2%. Этот вид, в противоположность барабинскому хомячку, проявляет более устойчивую численность п верхней части разреза.
Мышовка Sicista sp. является относительно редким видом. Ее останки найдены на уровне 7 горизонта 22.2, а также в отложениях горизонтов 21.1,12.3,12.1, 11.3 и 9.2. Численность мышовки колеблется в интервале 0,25 - 0,8%. В сборах 1994 г. этот грызун не зарегистрирован, что, возможно, объясняется чисто вероятностным фактором.
Тушканчик Allactaga sp. из группы земляных зайцев отмечен единичными зубами на уровнях 3 и 2 горизонта 22.1 и в отложениях горизонта 19.2.
Слепушонка Ellobius sp. спорадично встречается в средней и верхней частях разреза, начиная с уровня 1 горизонта 22.1.
Рыжие или лесные полевки Clethrionomys sp. составляют важнейший компонент тафоценозов Денисовой пещеры. Их костные остатки присутствуют во всех горизонтах разреза. Относительная численность рыжих полевок колеблется от 2 до 11%. Если учесть
результаты анализа современных погадок, то можно предполагать, что роль этих полевок в плейстоценовых сообществах долины Ануя была еще выше. В ископаемом состоянии, в отличие от современности, рыжие полевки представлены двумя видами - Clethrionomys rutilus иС. rufocanus.
Азиатские горные полевки AIticola sp. являются одной из самых массовых групп мелких млекопитающих плейстоцена. В среднем их численность составляет 10 -15% от общего количества мелких позвоночных, а в горизонтах 14.2 и 9.3 она превышает 20%. В ископаемой фауне азиатские горные полевки представлены двумя видами -Alticola strelzovi иA. macrotis. В современной фауне бассейна Ануя присутствует только первый вид -г плоскочерепная полевка.
Степная пеструшка Lagurus sp. - один из важных компонентов тафоценоза пещерных осадков. Этот вид встречается почти на всех горизонтах плейстоценовой толщи. Однако заметную роль пеструшка начинает играть в отложениях слоя 20. Здесь и выше по разрезу ее численность колеблется от 3 до 5%. В отложениях горизонта 9.2 количество пеструшки возрастает до 8,7%, а в осадках горизонта 9.1 достигает 17,9%. Важно отметить, что подобная тенденция отмечена в динамике численности по сборам 1994 г. Сходство проявляется и в заметном падении численности в интервале горизонтов 13.4- 12.3. Это сходство придает особую достоверность полученным результатам. В целом ископаемые пеструшки представлены видомLagurus lagurus. Специальные исследования показали, что пеструшки плейстоценовых популяций морфологически отличаются от современных.
Серые лемминги трибы Lemmini представлены единичными находками по всей толще отложений от уровня 10 горизонта 22.2 до горизонта 11.1. При этом только в отложениях слоя 20 количество останков лемминга достигает 3,2%. В остальных случаях оно не превышает 1 %.
Узкочерепная полевка Stenocranius gregalis - один из массовых видов плейстоценовых отложений. В каждом из слоев количество Mj узкочерепной полевки превышает количество Mj любого другого вида полевок в 2 - 3 раза. В среднем количество ее останков колеблется от 7 до 10%: в отложениях горизонтов 13.4 и 13.1 оно превышает 14%, а в горизонте 12.2 -15%. Относительная численность узкочерепной полевки возрастает вверх по разрезу до горизонта 12.2, а затем начинает снижаться. В целом похожую тенденцию отражают сборы 1994 г., хотя полного совпадения отдельных пиков по материалам разных квадратов не наблюдается.
Полевка-экономка Microtus oeconomus отмечена по всей толще плейстоценовых осадков. Однако ее численность редко достигает 3%. Только в отложениях горизонта 14.3 количество этой полевки возрастает до 5,9%. Правда в этом горизонте обнаружено небольшое количество костного материала и данный показатель может иметь случайный характер.
Северосибирская полевка Microtus hyperboreus -редкий представитель тафоценоза. Останки этой полевки встречаются спорадически, главным образом в средней и верхней частях разреза.
Темная полевка Microtus agrestis - редко встречается в отложениях плейстоценовой толщи. Отдельные зубы отмечены в слое 19, а также в горизонтах 14.2,14.1,13.4,12.1 и 11.4. Темная полевка - эволюционно молодой вид. Его выделение из общего ствола серых полевокMicrotini произошло в первой половине среднего неоплейстоцена. Современные морфологические черты этот вид приобрел во второй половине среднего неоплейстоцена.
Обыкновенная полевка Microtus arvalis тоже редка в плейстоценовых отложениях пещеры. Ее малочисленные останки зафиксированы от слоя 21 до горизонта 11.2.
Водяная полевка Arvicola sp. - редкий вид плейстоценовых осадков. Однако ее останки встречаются почти по всему разрезу: от уровня 5 горизонта 22.1 до горизонта 9.3. Водяная полевка представлена эволю-ционно продвинутой формой, близкой современнойArvicola terrestris.
Алтайский цокор Myospalax myospalax является одним из самых характерных элементов тафоценоза плейстоценовых отложений. Его останки встречаются практически на всех горизонтах разреза. Их относительная численность колеблется от 5 до 7%, а в слое 20 достигает 13%. Останки цокора являются важным палео-климатическим показателем. Они свидетельствуют об относительной стабильности природной среды на протяжении всего периода накопления плейстоценовых осадков. Примечательно, что в двух колонках 1994 и 1995 гг. фиксируется одинаковое падение численности цокора в осадках горизонта 22.3, а также в отложениях слоя 17 и нижней части слоя 14.
Пищуха Ochotona sp. представляет немногочисленный, но постоянно встречающийся компонент тафоценоза. Относительная численность пищухи, как правило, не достигает 3%. Однако на уровне 3 горизонта 22.1 она превышает 4,8%, а в слое 20 достигает 5,2%. В отложениях пещеры присутствует два вида пищухи: алтайскаяOchotona alpina и степнаяО. pusilla. Первый вид и сегодня обитает в бассейне Ануя. Горный Алтай является одним из районов его возникновения и формирования в процессе эволюционного становления. Степная пищуха - типичный обитатель равнинных степных ландшафтов Евразии. Его проникновение в бассейн Ануя могло произойти только в специфических природных условиях, т.е. при увеличении доли степных сообществ. Останки двух видов пищух подчеркивают своеобразие и пестроту плейстоценовых ландшафтов в окрестностях пещеры.
Заяц Lepus sp. редко встречается в костном материале мелкой фауны. Однако разрозненные зубы этого животного отмечены в плейстоценовой толще от уровня 8 горизонта 22.2 до горизонта 9.3. При этом их относительное количество не превышает 1-1,5%. Судя по размеру фрагментов и целых зубов, основная их часть принадлежит зайцу-толаю.
Мелкие куньи рода ласки Mustela sp. в небольшом количестве присутствуют почти по всему разрезу. В связи с фрагментарностью и малым количеством определимого материала вопрос о механизме их попадания в пещерные отложения пока остается открытым.
Птицы Aves. Кости птиц и скорлупа птичьих яиц часто встречаются в отложениях пещеры. В среднем их относительная численность составляет 5 - 7%, возрастая до 19,7% в слое 20. В толще слоя 22 число останков птиц резко снижается до 1,5- 2%. Возможно, это связано с большей залесенностью и закрытостью биотопов в эпоху накопления слоя 22. Подобная тенденция отмечена и в сборах 1994 г., хотя там она выражена менее отчетливо.
Рептилии Reptilia. Редко встречаются в плейстоценовых отложениях пещеры. Они представлены разрозненными фрагментами челюстей ящериц преимущественно в средней и верхней частях разреза.
Амфибии Amphibia. Кости конечностей лягушек спорадично встречаются по всему разрезу. Наибольшее количество останков амфибий отмечено в стратиграфическом интервале горизонтов 19.3 - 13.2.
Рыбы Pisces. Один из наиболее многочисленных компонентов тафоценоза пещеры. В среднем останки рыб составляют 10 - 12%. В отложениях горизонтов 21.1 и 9.3 их количество достигает более 28%. В пределах слоя 22 относительная численность костных остатков рыб снижается до 3 - 5%.
Динамика численности отдельных таксонов по сборам 1995 г. позволяет проследить общие закономерности изменения сообщества мелких млекопитающих за время накопления толщи плейстоценовых осадков Денисовой пещеры. В целом облик микротериофауны определяют типичные обитатели сухих и высокогорных степей — Stenocranius gregalis иAlticola strelzovi. Третьим важным компонентом является степная пеструшкаLagurus lagurus. От нижнего уровня слоя 22 до кровли плейстоценовой толщи встречаются останки характерных представителей лесных биотопов - полевок родаClethrionomys. Практически на всех уровнях разреза отмечены останкиAsioscalops altaica иMyospalax myospalax. Оба вида - специализированные землерои, которые не могут существовать при глубоком промерзании почвы. К видам, встречающимся в небольших количествах, но по всему разрезу, относятсяCricetulus barabensis иMicrotus oeconomus. Первый из них обитатель степей, второй - пойменных биотопов.
Высокая численность в составе фауны степных и нивальных видов предполагает существование значительных участков открытых ландшафтов в период накопления плейстоценовой толщи. Вместе с тем присутствие останков Clethrionomys, единичные находкиSciurus, Pteromys volans, Eutamias sibiricus свидетельствуют о постоянном наличии лесных массивов.
Обитатели леса наиболее многочисленны в слое 22, где доля Clethrionomys составляет на разных уровнях от 5 до 11% общего состава. Удельный весAlticola колеблется от 3,8 до 15%. Достаточно заметным элементом сообщества являетсяStenocranius - 3 - 5%. Интересно отметить обилие в этом слое костных остатков летучих мышей. На отдельных уровнях останки Chiroptera составляют более 60%. В слоях 20 - 9 их показатель не превышает 3 - 6%, что, очевидно, свидетельствует о возрастании фактора беспокойства со стороны человека. В целом накопление слоя 22, видимо, происходило в период широкого развития лесной растительности при умеренных климатических режимах.
В отложениях слоя 21 численность Clethrionomys существенно не меняется, удельный весAlticola увеличивается до 16,4%, а доляStenocranius составляет 4,3%. С этого уровня в ископаемой фауне начинают постоянно встречаться костные остаткиLagurus lagurus.
Своеобразные изменения в составе мелких млекопитающих отмечены в слое 20. Здесь резко сокращается численность Clethrionomys, снижается до минимума количествоAlticola иLagurus, а доляStenocranius gregalis возрастает до 7,2%. Для этого слоя характерны наиболее высокие значенияMyospalax myospalax (13%) иAsioscalops altaica (5,2%). Видимо, в период накопления слоя 20 происходило одновременное сокращение лесных массивов и участков горно-степных петрофильных ассоциаций. В это время наибольшего развития достигли лугово-степные ассоциации и ли-ственнично-березовые леса. Это была эпоха лугового и лесного разнотравья с минимальным участием злаков. Климат отличался теплыми зимами и дождливым летним периодом.
Выше по разрезу численность основных групп мелких млекопитающих восстанавливается в прежних соотношениях и сохраняет относительную стабильность до кровли слоя 14. В отложениях слоев 19-14 доля Clethrionomys варьирует в пределах 5 - 7%,Alticola -9 - 20%,Stenocranius - 5 - 10%,Lagurus - 3 - 5%.
В отложениях горизонта 13.2 отмечено резкое падение численности Clethrionomys иAlticola, количествоStenocranius gregalis снижается до 3,6%, исчезаетAsioscalops, Lagurus иMicrotus oeconomus. Одновременно увеличивается численностьLemmini. Вероятно, формирование этого горизонта связано с эпохой господства горно-степных петрофильных и нивальных злаково-осоковых ассоциаций в условиях относительно холодного и сухого климата.
Общий облик тафоценоза восстанавливается на уровне горизонта 13.1. Здесь численность Clethrionomys достигает 6,5%,Alticola - 13,3%,Stenocranius gregalis - 14,4%, появляютсяAsioscalops иMicrolus оесопотш. Однако такая стабилизация природной обстановки была кратковременной.
В интервале слоев 12-9 наблюдается заметная перестройка в составе плейстоценовой биоты. На фоне относительно высокого удельного веса Alticola (12-17%) иStenocranius (8 - 15%) удельный весClethrionomys снижается до 1 - 2%, aLagurus - резко возрастает, достигая своего максимума (17,8%) в отложениях горизонта 9.1. Эти данные свидетельствуют о значительных изменениях ландшафтной обстановки. Очевидно, накопление осадков этой части разреза проходило в период сокращения участков леса и увеличения площади открытых биотопов, занятых в основном горно-степными петрофильными ассоциациями. Особенно ярко эта тенденция выражена в отложениях слоя 9.
Диаграммы состава мелких позвоночных имеют незавершенный характер. Нарастание остепнения, ухудшение климатической обстановки не достигают здесь своего максимума. Исходный материал не демонстрирует перехода от плейстоценовой биоты к го-лоценовой. Эти данные указывают на отсутствие в толще пещерных отложений осадков финального этапа плейстоцена.
Все мелкие млекопитающие, представленные в та-фоценозе пещеры, по своей морфологии мало отличаются от аналогичных современных таксонов. Вместе с тем данное сообщество по структуре тафоценоза, количественному соотношению входящих в его состав видов, напротив, принципиально отличается от современных сообществ мелких млекопитающих бассейна Ануя.