

Джефферсон - Сборник упражнений и задач - 2000 / Student Guide12
.pdf85 |
Глава 12: Регуляция метаболизма углеводов и жиров |
Глава 12
Регуляция метаболизма углеводов и жиров
Резюме главы
В этой главе описывается механизм регуляции двух главных энерго-метаболизирующих систем углеводов и жиров. На ферментативном уровне рассматривается регуляция обмена углеводов и жиров в зависимости от избытка или недостатка пищевых продуктов в организме, а также в критических ситуациях срочной потребности в энергии. Эти процессы должны координировано протекать в различных типах тканей, участвующих в запасании и использовании этих пищевых молекул. В главу включено обсуждение модели МихаэлисаМентен ферментов с аллостерической регуляцией, роли фосфорилирования белка в гормональной регуляции с участием вторичного мессенджера.
Цели урока
♦Суммарное направление метаболического потока в условиях изобилия и голода.
♦Проблема “холостого” (субстратного) цикла и системы его преодоления.
♦Термодинамические различия между реакциями, “выбранными” в качестве пунктов регуляции.
♦Два основных механизма обратимого изменения скорости ферментативной реакции.
♦Факторы, влияющие на количество фермента, присутствующего в системе, и концепция времени полужизни белков.
♦Важность модели ферментативной кинетики Михаэлиса-Ментен и
влияние величин [S], Km è Vmax на скорость катализируемой ферментом реакции.
♦Концепция регуляции по принципу обратной связи.
♦Значение аллостерической регуляции ферментативных реакций и графическая интерпретация данных о ее влиянии на ферментативную кинетику.
♦Две модели, предложенные для описания механизма аллостерической регуляции.

86 |
Глава 12: Регуляция метаболизма углеводов и жиров |
♦Значение фосфорилирования и дефосфорилирования в регуляции активности ферментов, в которой могут участвовать боковые группы аминокислот.
♦Роль киназ и фосфатаз в ковалентной модификации белков.
♦Различия между внутренней и внешней регуляцией ферментативной активности и роль гормонов и нейромедиаторов.
♦Важные регуляторные звенья в процессах расщепления и синтеза гликогена.
♦Регуляция пируватдегидрогеназы - фермента, осуществляющего взаимосвязь между расщеплением жирных кислот, гликолизом и циклом лимонной кислоты.
♦Концепция энергетического заряда, нашедшая свое отражение в концентрациях ATP/ADP/AMP и NADH/NAD+.
♦Важные звенья регуляции расщепления и синтеза жирных кислот и необходимость координирования двух этих процессов.
♦Влияние гормонов глюкагона, инсулина и адреналина на мышцу, печень и жировую ткань и координация их эффектов.
♦Значение вторичного мессенджера сАМР и роль рецепторов клеточной мембраны в регуляции фермента аденилатциклазы.
♦Значение сАМР-зависимых протеинкиназ (ПКА).
♦Значение усиления сигнала в процессе функционирования киназ.
♦Механизм снятия активации системы под действием сигнала.
♦Значение нового метаболита фруктозо- 2,6-дифосфата.
Прогулка по главе
Регуляция ферментативной активности
Обмен углеводов и жиров определяет поток метаболитов в большинстве систем. Направление этого потока веществ часто изменяется, поскольку изобилие и голод присущи многим живым организмам. Сбалансированное состояние является той целью, к достижению которой направлено совместное функционирование различных метаболических путей и в этом участвуют механизмы передачи информации.
Говоря простым языком, если реакции протекают в одном направлении, то реакции, идущие в противоположном должны быть ………………(1). Различные системы могут нуждаться в
…………….(2) и доступных …..………(3), поэтому необходимо преодолеть возможную проблему
……………….(4) циклов или субстратных циклов (аналогично короткому замыканию в электросети). Реципрокная регуляция заключается в том, что …………….(5) сигнал …….............….(6) один путь и ……………..(7) другой. Напомним, что такая регуляция происходит
Глава 12: Регуляция метаболизма углеводов и жиров |
87 |
|
|
осуществляется на уровне термодинамически …………….. (8) этапов (большие величины DG), контроль за которыми может осуществляться раздельно на каждом из путей. Легко обратимая реакция, катализируемая одним и тем же ферментом в ……………………(9), не может подвергаться реципрокной регуляции в двух направлениях.
Для систем, нуждающихся в изменении направления ответа, обратимая регуляция является более приемлемой, чем необратимая. Есть два основных пути обратимого изменения активности фермента: изменение количества ...…………….(10) или ..…………….(11) катализируемой им реакции. Количество любого фермента определяется скоростью …………….(12) (и активации) данного белка и его ..………………..(13). Время полужизни белка определяется тем (средним) промежутком времени, который необходим для деградации ..……………………(14) его молекул.
………………..(15) представляют собой относительно короткоживущие молекулы с временем полужизни, варьирующим от одного часа да нескольких дней. Это .........…………..(16) тип регуляции, медленно реагирующий как на стимулирующие, так и на тормозные воздействия.
Ответы: (1) выключены; (2) энергии; (3) пищевых продуктах; (4) “холостых”; (5) один и тот же;
(6) выключает; (7) включает; (8) необратимых; (9) в обоих направлениях; (10) фермента; (11) скорости; (12) синтеза; (13) деградации; (14) половины; (15) Белки; (16) долговременный.
Модель Михаэлиса-Ментен
Активность синтезированных ферментов может очень быстро и тонко регулироваться. Наше понимание такой модели регуляции основано на механизме действия ферментов, предложенном
.......……...................…………..(1). Согласно этому механизму, фермент (Е) и субстрат (S) обратимо соединяются с образованием комплекса (ES), из которого возникает и продукт реакции (Р), а фермент вступает в новый цикл. Концентрация субстрата (S) оказывает влияние на скорость реакции, сдвигая равновесную концентрацию ……………….(2); зависимость скорости реакции от [S] характеризуется ...……………(3) кривой с насыщением. Это означает, что выше определенной концентрации субстрата [S] фермент функционирует с ………………(4) скоростью и не может работать быстрее. В этом случае фермент .....………….(5) субстратом и функционирует при …….………..(6). Чем ……………….(7) фермент связывается с субстратом, тем меньшая концентрация субстрата необходима для достижения ……………….(8). Величина, характеризующая прочность фермент-субстратного комплекса, называется …………….(9) и определяется как концентрация субстрата [S], при которой фермент работает со скоростью, составляющей ………….(10). Поэтому, …………………(11) значения Кm указывают на существование прочного ES комплекса. Концентрации субстратов в клетке обычно сопоставимы со значениями Кm.
Ответы: (1) Михаэлисом и Ментен; (2) ES комплекса; (3) гиперболической; (4) максимальной; (5) насыщен; (6) Vmax; (7) прочнее; (8) Vmax; (9) Êm; (10) 1/2Vmax; (11) малые.

88 |
Глава 12: Регуляция метаболизма углеводов и жиров |
Аллостерическая регуляция
Обычно объектом регуляции в метаболическом пути является один, часто ……………(1) фермент. Регуляция по принципу обратной связи (называемая также ингибированием конечным продуктом) включает торможение первой реакции данного метаболического пути высокими концентрациями конечного метаболита. Двумя главными механизмами регуляции ферментативной активности без изменения количества фермента, являются аллостерический контроль и ковалентная модификация. В аллостерической регуляции участвуют молекулы ………………(2), которые связываясь с ферментом либо ускоряют, либо тормозят реакцию. Наиболее общий механизм аллостерической регуляции включает изменение сродства фермента к субстрату; этот механизм известен как ……………..(3) эффект. Ферменты такого типа часто являются олигомерными белками
èобычно содержат две и более каталитических субъединиц. Взаимодействие этих субъединиц изменяет чувствительность фермента к концентрации …………..(4). Графически это выражается
………………….(5), “S”-образной кривой. Активаторы сдвигают кривую ………………(6), и снижают ……………..(7), при этом образуется ……………….(8) ES-комплекс. Ингибиторы
(дезактиваторы) сдвигают кривую вправо и поэтому …………….(9) значение Кm, в этом случае фермент образует менее прочный …………….(10) комплекс.
Для объяснения аллостерического эффекта предложено две модели. Согласно …….....................……(11), субъединицы олигомерного белка-фермента могут существовать в состоянии с высоким или с низким сродством к субстрату (иначе говоря, в высокоили низкоаффинном состоянии), причем все они должны находиться в одном и том же конформационном состоянии. Эти два состояния обозначаются соответственно как расслабленное
èнапряженное. Связывание молекулы субстрата вызывает переход фермента из ………….(12) в
……………..(13) аффинное состояние. Активные центры каждой молекулы фермента могут находиться в одном из двух этих состояний либо в состоянии с высоким сродством, либо - с низким. Обе формы комплекса при этом находятся в динамическом равновесии, которое сдвинуто в сторону белка с …………..(14) аффинностью субъединиц. Поэтому при ……………..(15) концентрации субстрата большая часть фермента переходит в ……………..(16) аффинное состояние. Связывание аллостерических модуляторов сдвигает положение равновесия между низко- и высокоаффинными конформациями фермента путем предпочтительного взаимодействия с молекулами фермента, находящихся в одном из двух состояний. При кооперативном связывании субстратов ферменты переходят из напряженного состояния в расслабленное.
Согласно последовательной модели, все молекулы фермента или субъединицы исходно находятся в низкоаффинном состоянии и равновесия нет, однако по мере связывания субстрата
..............………………(17) происходит изменение конформации фермента от Т (напряженного) к R (расслабленному), что облегчает аналогичное изменение ..……………(18) субъединицы. Такой переход фермента от Т к R конформации описывается сигмоидной кривой. Этот процесс распространяется на все субъединицы. Обе модели дают удовлетворительное объяснение механизму
Глава 12: Регуляция метаболизма углеводов и жиров |
89 |
|
|
аллостерической регуляции, который в действительности может представлять их комбинацию или некий промежуточный вариант.
Аллостерический контроль может осуществляться молекулами, структура которых обычно не похожа ни на субстрат, ни - продукта реакции. Это позволяет очень тонко координировать функционирование метаболических путей. Метаболиты одного пути обмена могут служить регуляторами другого и ферменты могут иметь более одного эффектора.
Ответы: (1) первый; (2) лиганда; (3) Кm; (4) субстрата; (5) сигмоидной; (6) влево; (7) Кm; (8) более прочный; (9) повышают; (10) ES; (11) модель согласованного механизма; (12) низко-; (13) высоко-; (14) низкой; (15) увеличении; (16) высоко; (17) каждой субъединицей; (18) соседней.
Регуляция фосфорилированием
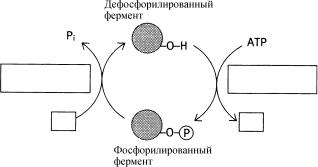
Во избежание “химического хаоса”, приводящего к избытку или недостатку метаболитов, каждый метаболический путь нуждается в информации о функционировании другого. Фосфорилирование представляет собой форму ковалентной модификации фермента с целью изменения его активности. Реакцию переноса фосфата с АТР на другую молекул катализируют ферменты киназы; фосфорилирование белка часто вызывает изменение активности в результате конформационных изменений его структуры. Процесс химически обратим и фосфат удаляется в реакции, которую катализирует фермент фосфатаза. Для образования высокоэнергетической фосфоэфирной связи часто используются гидроксильные группы аминокислот серина, треонина и иногда тирозина.
Заполните следующее
Внутренние и внешние уровни регуляции
Регуляция активности киназ и фосфорилаз, которые соответственно присоединяют или удаляют фосфатные группы, часто осуществляется извне клетки под действием ……………(1) и
……………..(2). Внутренний контроль достигается посредством ……………….(3) механизмов и осуществляется автоматически на уровне саморегуляции. В задачу внешних механизмов входит координация “усилий” различных ………………(4). Взаимопревращение …………….(5) гликогена
90 |
Глава 12: Регуляция метаболизма углеводов и жиров |
|
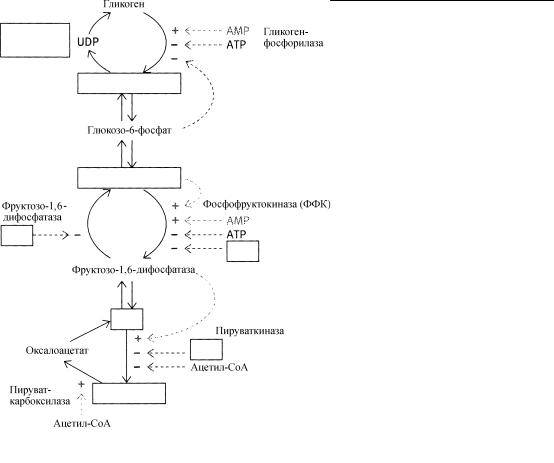
|
и фосфорилированных …………………(6) глюкозы катализируют соответственно синтаза и фосфорилаза. Гликоген ……………….(7), регулируется внешними механизмами в зависимости от поступления пищевых молекул. …………………….(8), катализирующая образование глюкозо- 1-фосфата, существует в двух формах: менее активной фосфорилазы b и более активной фосфорилазы ……………(9). Однако неактивная фосфорилаза ……………….(10) может частично активироваться под действием аллостерического регулятора АМР, высокие концентрации которого накапливаются в условиях дефицита энергии. Этой частичной активации фосфорилазы b под действием ……………….(11) противодействуют ………(12) и …………………(13). Регуляторные эффекты этих соединений также проявляются при высоких концентрациях, что указывает на отсутствие потребности системы в генерируемой при гликолизе энергии. Присутствие обеих b и а форм фосфорилазы позволяет ……………..(14) механизмам регуляции превалировать над
…………………(15).
Ответы: (1) гормонов; (2) нейромедиаторов; (3) аллостерических; (4) клеток; (5) полимеров; (6) мономеров; (7) синтаза; (8) Фосфорилаза; (9) а; (10) b; (11) АМР; (12) АТР; (13) глюкозо-6-фосфат; (14) внешним; (15) внутренними.
При накоплении АМР (указывающего на потребность клеток в энергии) активность фосфофруктокиназы ……………….(1), а фруктозо-1,6-дифосфатазы ……………….(2). Фруктозо- 6-фосфат активирует фосфофруктокиназу, которая вызывает повышение уровня фруктозо-1,6- дифосфата и активирует ……………..(3) ……………..(4). Активирующий эффект АМР и ФФК уравновешивается ингибирующим действием ………………..(5). Лимонная кислота - это название в большинстве случаев используется наравне с цитратом, - также накапливается при увеличении концентрации АТР и также ингибирует активность ………………….(6), так что гликолиз подавляется при поступлении “сырья” в ……………….(7). Аналогичным образом, ацетил-СоА (поступающий в цикл лимонной кислоты) ингибирует активность ………………….(8) - фермента, приводящего к образованию пирувата, который затем может превратиться в ацетил-СоА. АцетилСоА активирует пируват …………(9). Это приводит к увеличению образования
……………….(10), конденчация которого с ацетил-СоА способствует нормальной работе цикла лимонной кислоты.
Ответы: (1) стимулируется; (2) ингибируется; (3) пируват; (4) киназу; (5) АТР; (6) ФФК; (7) цикл лимонной кислоты; (8) пируваткиназы; (9) карбоксилазу; (10) оксалоацетата.
Глава 12: Регуляция метаболизма углеводов и жиров |
91 |
Заполните следующее
Пируватдегидрогеназное звено
Пируватдегидрогеназа, катализирующая превращение ………………….(1) в ……………..(2), играет стратегическую регуляторную роль, соединяя гликолиз с …………….(3) и синтезом жира. Продукты реакций - ацетил-СоА и …………(4), - ингибируют ………………(5), а субстраты
………………(6) è NAD+ ………………….(7) активность. Эта дегидрогеназа также является объектом регуляторного влияния …………(8), осуществляемого в ходе фосфорилирования киназой. Последняя является частью целого дегидрогеназного ферментного комплекса. Функционирование цикла лимонной кислоты и электронтранспортной системы фосфорилирования АТР во многом зависит от обеспеченности ………………….(9) NAD+ и ADP. Как правило, высокий энергетический заряд системы, оцениваемый по высокой концентрации ………(10) и
…………………(11), блокирует функционирование образующих энергию путей, в то время как при …………….(12) энергетическом заряде (когда накапливаются высокие концентрации ADP и NAD+, происходит ………….(13) системы для «подзарядки метаболической батареи».
92 |
Глава 12: Регуляция метаболизма углеводов и жиров |
|
|
Ответы: (1) пирувата; (2) ацетил-СоА; (3) циклом лимонной кислоты; (4) NADH; (5) фермент; (6) CoASH; (7) стимулируют; (8) АТР; (9) субстратами; (10) АТР; (11) NADH; (12) низком; (13) стимуляция.
Запасание жиров
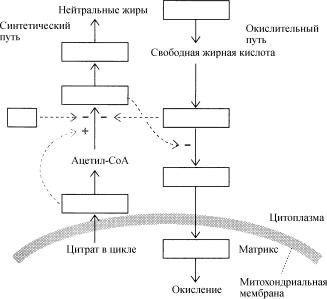
Расщепление и синтез жирных кислот скоординированы таким образом, что они взаимно подавляют друг друга. Жирные (длинноцепочечные) ацил-СоА ингибируют активность
…………………(1), которая катализирует первый обязательный этап в синтезе жиров. В свою очередь первый продукт синтеза жиров, ……………..(2), ингибирует перенос ацильных СоА производных жирных кислот на ……………..(3). Ацетил-СоА-карбоксилаза активируется цитратом. Когда уровень последнего достаточно высок он транспортируется из ………………(4),
что служит сигналом к запасанию жиров. Основным механизмом регуляции активности ацетил- СоА-карбоксилазы является ее инактивация под действием …………………..(5), стимулируемого
……………….(6). Реакция дефосфорилирования, снимающая ……………………(7) активности ацетил-СоА-карбоксилазы регулируется гормонами. Информация о физиологических потребностях целого организма передаются отдельным клеткам при помощи гормонов и нейромедиаторов.
Ответы: (1) карбоксилазы; (2) малонил-СоА; (3) карнитин; (4) митохондрий; (5) фосфорилирования; (6) АМР; (7) ингибирование.
Заполните следующее

Глава 12: Регуляция метаболизма углеводов и жиров |
93 |
Роль гормонов
Основными гормонами, участвующими в регуляции обмена углеводов и жиров, являются
……………….(1), …………………(2), а также катехоламины ……………..(3) и ………………(4). Глюкагон сигнализирует о …………..(5) уровне сахара в крови, инсулин - о ……………….(6), а адреналин и норадреналин - об острой потребности организма в ……………..(7). Инсулин секретируется специализированными клетками .......……................……..(8) и характеризуется коротким временем полужизни в крови. Поэтому он действует в течение короткого времени при повышении уровня сахара в крови выше нормы (90 мг%).
Гормоны представляют химические сигнальные молекулы, которые высвобождаются в кровь
èдействуют только на…………………(9). Гормоны можно разделить на два типа: …………….(10)
растворимые и ………..(11) растворимые. Три рассматриваемых в этой главе гормона относятся к числу водорастворимых; они взаимодействуют с ………………….(12) на внешней поверхности клеточной мембраны. Только …………(13) имеют соответствующий рецептор, воспринимающий сигнал данного гормона. Связываясь с рецептором, гормон передает гуморальный сигнал клетке. При этом изменяется уровень .......…………………(14) - внутриклеточной регуляторной молекулой. В случае глюкагона и адреналина вторичным мессенджером служит циклический АМР (сАМР), который образуется, когда это необходимо, из ………(15) под действием фермента …………….(16). сАМР аллостерически активирует …………(17) и эти киназы фосфорилируют ферменты. Эта группа киназ получила называние сАМР-зависимые протеинкиназы (ПКА). В отсутствие сАМР протеинкиназа А представляет собой ……….(18), построенный из двух каталитических и двух регуляторных …………..(19). сАМР связывается с ……………..(20) субъединицами, в результате чего и каталитические субъединицы становятся полностью активными. ……...………..(21) гидролизует сАМР, что приводит к снижению его уровня и возвращению ПКА в неактивное состояние. Фосфорилированные белки теряют фосфатную группу под действием …………….(22)
èсистема возвращается в исходное состояние. Количество рецепторных белков на наружной поверхности клетки находится под контролем и может снижаться при чрезмерной стимуляции клетки. Активность рецептора может изменяться в результате его фосфорилирования, происходящего внутри клетки. Под активностью здесь понимается способность рецептора передавать сигнал в клетку.
Ответы: (1) глюкагон; (2) инсулин; (3) адреналин; (4) норадреналин; (5) низком; (6) высоком; (7) энергии; (8) поджелудочной железы; (9) клетки-мишени; (10) водо-; (11) жиро-; (12) белком рецептором; (13) клетки-мишени; (14) вторичного мессенджера; (15) АТР; (16) аденилатциклазы; (17) киназы; (18) тетрамер; (19) субъединиц; (20) регуляторными; (21) Фосфодиэстераза; (22) фосфатазы.

94 Глава 12: Регуляция метаболизма углеводов и жиров
Координация запасания и использования
Обычно, глюкагон и инсулин выполняют противоположные и дополняющие друг друга функции, хотя механизм действия инсулина пока полностью непонятен. Клетка поглощает глюкозу в результате …………………(1) при помощи специального мембранного транспортного белка в зависимости потребности в ………..………(2). Поглощение глюкозы клетками мышц и жировой ткани зависит от уровня ......………(3), а в ………………(4) инсулин не оказывает влияния на поглощение глюкозы. Тот факт, что фермент головного мозга …………….(5) имеет более низкое значение Кm для глюкозы (более прочное связывание), чем фермент печени ……………(6), уже обсуждался ранее. Он обусловлен более важной ролью, которую выполняет глюкоза в головном мозге, по сравнению с печенью. …………….(7) повышает активность глюкокиназы, индуцируя синтез этого белка в ответ на ……………(8) уровень сахара в крови. Что касается жиров, то свободные жирные кислоты проникают через мембрану …………….(9) за счет их гидрофобности. Поэтому, уровень жирных кислот, поступающих в …………….(10) и в ………….(11) регулируется посредством одинаковых механизмов.
Ответ: (1) облегченной диффузии; (2) энергии; (3) инсулина; (4) печени и головном мозге; (5) гексокиназа; (6) глюкокиназа; (7) Инсулин; (8) высокий; (9) самостоятельно; (10) клетку; (11) кровь.
Расщепление гликогена
Расщепление гликогена инициируется ферментом гликогенфосфорилазой, которая существует в двух формах - “а”, которая …………….(1) в отсутствии …..……..(2), и “b”, которая активна только в присутствии АМР. Гликогенфосфорилаза b превращается в а форму под действием
……………..(3), которая ее фосфорилирует. Адреналин оказывает такой же эффект при участии
………….(4), выполняющего функцию вторичного мессенджера. Адреналин - гормон, участвующий в реализации организмом экстремальных реакций (борьбы и бегства), он подает сигнал метаболической тревоги, вызывая синтез большого количества АТР. В печени адреналин индуцирует расщепление гликогена, координированное с высвобождением ……………..(5) в
……………(6) и таким образом повышает снабжение мышц энергией. Глюкагон оказывает на клетки печени такой же эффект, как и адреналин. Прохождение сигнала от вторичного мессенджера сАМР до инициации расщепления гликогена является опосредованным и включает активацию киназы, которая активирует другую киназу, а та в свою очередь активирует расщепляющий гликоген фермент. Киназа, расположенная между ПКА, активируемой сАМР и гликогенфосфорилазой b, расщепляющей гликоген, называется ……………..(7). Этот непрямой путь приводит к …………..(8) усилению сигнала. Одна молекула гормона может вызывать образование нескольких молекул сАМР и каждая из них способна активировать следующий ферментативный процесс, вызывая многократное усиление на каждом этапе.
Механизм выключения расщепления гликогена включает ………….(9) высокоэнергетической фосфатной связи фосфатазой. В печени активность этого фермента стимулирует свободная глюкоза,