

5. Ферменты
Вещества белковой природы, которые катализируют химические превра-щения в живых клетках, называют ферментами или энзимами. Здесь не просто лингвистическое расхождение. На начальном этапе развития био-химии шел спор между Л.Пастером с одной стороны и М.Бертло и Ю.Либихом с другой. Л.Пастер был убежден в том, что процесс брожения неотделим от дрожжевых клеток и осуществляющие это превращение катализаторы – организованные ферменты (от лат. fermentum – закваска) – от клеток неотделимы. М.Бертло и Ю.Либих оперировали понятием энзим (от гр. en - в, zyme – закваска, то есть то, что в закваске), относя его к секретируемым клетками катализаторам пищеварительных процессов – пепсину, трипсину, амилазе и др. Заблуждение Пастера было основано на том, что он проводил стерилизацию клеточных культур нагреванием, при котором шла и денатурация ферментов, а М.Бертло и Ю.Либих работали с ферментами из пищеварительных соков, не проводя их термическую обра-ботку. В соответствии с этим биокатализаторы вне продуцирующей их клетки принято назвать энзимами, то есть ферменты в составе стирального порошка, в названии которого есть приставка «био», или в таблетке фестала – это энзимы. В биохимии, рассматривающей обменные процессы в клетках, речь обычно идет о ферментах, хотя относящийся к ним раздел биохимии называют энзимологией для того чтобы понятие было состав-лено только из греческих слов.
Работу ферментов выбрал для иллюстрации особенности биохимических процессов известный биохимик Сент-Дьердь. Во введении к своей книге по биохимии он напомнил, что в гробнице Тутанхамона несколько тысяч лет пролежала высохшая трапеза фараона, оставленная ему для загробной жиз-ни. Будучи съеденной, она превратилась бы в диоксид углерода, воду и другие продукты метаболизма в считанные часы.
Надо сказать, что начало биохимии в нашем сегодняшнем понимании было положено работами Э.Бухнера, показавшего, что после длительного пере-мешивания дрожжевых клеток с инфузорной землей происходит их разру-шение, но полученный бесклеточный сок некоторое время сохраняет спо-собность к сбраживанию глюкозы. На самом деле аналогичные опыты ста-вились намного раньше, например, за 25 лет до публикации Э.Бухнера появилась статья на эту тему М.М.Манасеиной (Коркуновой) на русском и немецком языках, были и более ранние сообщения.
Что же отличает ферменты, как катализаторы? В общем можно выделить три особенности:
Высокая эффективность. Под этим подразумевается, что реакции, ко-торые в обычных условиях идут очень медленно или вообще не идут, в присутствии ферментов протекают с очень высокой скоростью. Правда, скорость при этом понятие относительное. Если одна молеку-ла пепсина вызывает расщепление примерно десяти пептидных связей в секунду, то молекула каталазы, разлагающая пероксид водорода на воду и кислород, осуществляет десятки тысяч превращений в секунду.
Ферменты высокоспецифичны по типу реакций, то есть каждый фер-мент катализирует только одну реакцию, например, глютаминовая кислота действием соответствующей декарбоксилазы превращается в -аминомасляную кислоту, а в присутствии фермента, относящегося к трансаминазам, из нее образуется кетоглутаровая кислота.
Ферменты чаще всего специфичны, избирательны, по субстрату. На-пример, превращение гидроксильной группы в карбонильную осу-ществляют ферменты, относящиеся к дегидрогеназам, но этанол превращается в ацетальдегид алкогольдегидрогеназой, а молочная кислота в пировиноградную – лактатдегидрогеназой, хотя, конечно, есть и ферменты с невысокой избирательностью, например, пищева-рительные ферменты.
Из этого следует, что ферментов, осуществляющих все многообразие метаболических превращений, должно быть очень много. В 1930 году было известно всего лишь 80 ферментов, а в 1994 году – 3200. Долгое время безуспешно искали ферменты не только среди белков, но и среди других классов биополимеров, и лишь в последние годы было показано, что превращения определенных субстратов могут катализировать и неко-торые рибонуклеиновые кислоты, названные рибозимами.
Ферменты, выделенные и охарактеризованные на ранней стадии развития биохимии, получали имена собственные (трипсин, пепсин, тромбин), затем названия ферментов стали образовывать по названию субстратов с оконча-нием -аза (аргинин аргиназа, аспарагин аспарагиназа, мочевина уреаза и т.д.), но чаще всего название субстрата дополняют типом осу-ществляемого ферментом химического превращения (лактатдегидрогеназа, холинэстераза, супероксиддисмутаза). Обилие таких названий может на-пугать приступающего к изучению биохимии, но этот испуг легко преодо-лим, так как в конце концов даже специалисты имеют обычно дело с огра-ниченным числом биохимических превращений. Если выделять из биохи-мии раздел энзимологии, то такая путаница в названиях становится слиш-ком обременительной, и поэтому была принята цифровая классификация ферментов, включающая четыре группы цифр: класс, подкласс, подпод-класс и номер фермента в подподклассе. Исторически выделилось шесть классов ферментов.
Первый класс представлен оксидоредуктазами. В их число входят дегид-рогеназы, то есть ферменты, катализирующие дегидрирование спиртовых групп до карбонильных групп, окисление карбонильных групп до карбок-сильных, дегидрирование с образованием этиленовых соединений, дегид-рирование групп СН-NН2, приводящее к образованию иминов (C=NH), и так далее. Реакции дегидрирования протекают по схеме:
![]()
Чаще всего в роли акцепторов водорода выступают NAD(P)+ или флави-новые коферменты дегидрогеназ. Если атомы водорода или электроны непосредственно переносятся на кислород, то такие ферменты относятся к подклассу оксигеназ.
Второй класс представлен трансферазами – это ферменты, катализирую-щие перенос многоатомных групп с одной молекулы на другую. Сюда вхо-дят, например, ферменты, переносящие фосфатные остатки с аденозинтри-фосфата на молекулы сахаров (фосфофруктокиназа), ферменты, перено-сящие одноуглеродные фрагменты (фолат-зависимые ферменты). К транс-феразам относятся также трансаминазы, обменивающие аминогруппы на карбонильные функции.
Ферменты третьего класса называют гидролазами. Они катализируют перенос двух фрагментов молекулы на молекулу воды, то есть реакцию гидролиза. Если они гидролизуют пептидные связи в белках, то это проте-азы или пептидазы. Отщепление С‑концевой аминокислоты катализирут карбопептидазы, а N-концевой – аминопептидазы. Гидролитическое рас-щепление нуклеиновых кислот проводят нуклеазы (ДНК-аза, РНК-аза).
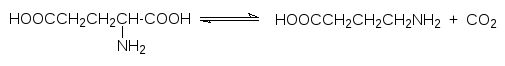
Четвертый класс ферментов – это лиазы. Они катализируют расщепление субстрата с образованием кратной связи или, в обратном направлении, – присоединение по кратной связи (в этом варианте обратной реакции их называют обычно синтазами). В качестве примера можно привести карбо-ангидразу, катализирующую реакцию образования бикарбоната из диок-сида углерода и воды:
![]()
Еще один пример – декарбоксилазы:
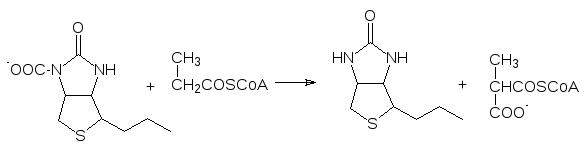
К пятой группе относятся изомеразы. Уже их название характеризует тип катализируемых этими ферментами превращений. Можно только доба-вить, что наряду с простыми изомеризациями, связанными, например, с переносом ацильной (фосфорильной) группы с одной гидроксильной груп-пы на другую, изомеразы катализируют и более сложные реакции. Напри-мер, к ним относятся рацемазы и эпимеразы (если речь идет о получении другого диастереоизомера), а кобаламинзависимые изомеразы по свобод-норадикальному механизму катализируют и перестройку углеродного ске-лета молекул. С их участием, например, метилмалоновая кислота в виде ее тиоэфира с коферментом А превращается в сукцинил-кофермент А:
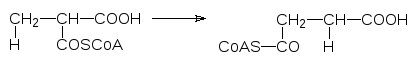
Шестая группа ферментов – лигазы (синтетазы). Главная особенность этих ферментов состоит в том, что энергию, требуемую для протекания катализируемых ими превращений, поставляют макроэргические вещест-ва, главным из которых является АТФ. Примером катализируемых лигаза-ми реакций служит образование C-N-, C-S- и С-С-связей (например, пеп-тидных связей, тиоэфиров и т.д.).
Набор функциональных групп в белковых аминокислотах невелик и с их помощью белковая молекула не может обеспечить катализ всех требуемых для протекания метаболических процессов реакций. Вследствие этого во многих случаях к работе фермента подключаются вещества небелковой природы, которые могут быть связаны с белковой молекулой ковалентны-ми связями, солевыми связями или же они могут образовывать более или менее прочные комплексы с белковой частью фермента, которую называ-ют апоферментом. Связанные с апоферментами ковалентной связью не-белковые составляющие называют простетическими группами, а связан-ные с апоферментами водородными связями, солевыми, Ван-дер-Ваальсо-выми или гидрофобными взаимодействиями небелковые компоненты на-зывают коферментами (в последнее время входит в употребление термин косубстрат, то есть дополнительный, вспомогательный субстрат). Облада-ющий каталитической активностью комплекс кофермента и апофермента, если это надо подчеркнуть, называют холоферментом. Кофермент или простетическая группа отвечают за химизм катализируемого превращения. Образование холофермента стабилизирует апофермент, поскольку при этом реализуется наиболее выгодная в энергетическом отношении струк-тура, поэтому апофермент легче денатурируется, чем холофермент. Но белковая компонента не только определяет специфичность фермента по субстрату, обеспечивает связывание с субстратом и отход от каталитичес-кого центра продуктов реакции. В отдельных случаях, как говорилось выше, она определяет и химизм катализируемой реакции. Кроме кофер-ментов или простетических групп в состав ферментов могут входить и кофакторы – чаще всего это ионы металлов, известные нам как микро-элементы.
Существуют, конечно, и ферменты, каталитический центр которых состав-лен только функциональными группами аминокислот, но и они чаще всего наиболее активны при наличии в их структуре кофакторов. Так, например, в работе каталитического центра сериновых гидролаз принимает участие ион цинка.
Особую роль в метаболических процессах играют аллостерические фер-менты, молекулы которых, кроме каталитических центров, имеют еще и регуляторные участки. В результате связывания с ними веществ, называе-мых эффекторами, активность фермента может возрастать (положитель-ный эффектор) или снижаться (отрицательный). Аллостерические фермен-ты имеют более сложное строение и более высокую молекулярную массу.
Строение многих коферментов и простетических групп было установлено еще в тридцатые годы прошлого века. Оказалось, что чаще всего они обра-зуются в результате более или менее сложных превращений витаминов. Так, например, витамин В1 или тиамин, входит в состав лиаз, декарбокси-лирующих -кетокислоты (пируватдекарбоксилаза, пируватдегидрогеназа, кетоглутаратдегидрогеназа) в виде эфира с пирофосфорной кислотой – тиаминпирофосфата:

Кофермент пиридоксальфосфат, принимающий участие в реакциях пере-аминирования (трансаминазы) и декарбоксилирования аминокислот (лиа-зы), точно так же образуется в результате монофосфорилирования витами-на В6 – пиридоксаля:
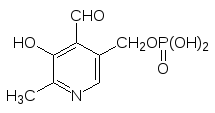 и
и
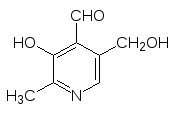
Более сложная цепочка превращений ведет от витамина В3 – ниацина (так называют никотиновую кислоту и никотинамид) к коферменту реакций гидрирования-дегидрирования – никотинамидадениндинуклеотиду NAD+:
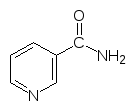 и
и

Еще одна группа ферментов, принимающих участие в реакциях гидрирова-ния-дегидрирования, представлена флавопротеинами (FMN и FAD – фла-винмононуклеотид и флавинадениндинуклеотид), у которых донором-акцептором атомов водорода является рибофлавин (витамин В2):
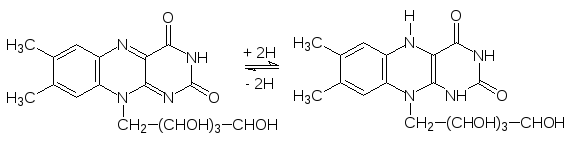
Важную роль в окислительно-восстановительных превращениях в биоло-гических системах играют цитохромы (цито – от греч. kytos – вмещающее, клетка; хрома – цвет, окрашивание). В их состав входят железопорфирины, которые представляют собой макроцикл из четырех связанных метиновы-ми мостиками пиррольных колец с атомом железа в центре. В качестве примера здесь можно привести структуру гема – железопорфирина, вхо-дящего в состав гемоглобина и предназначенного для переноса молекуляр-ного кислорода. Гем соединен с белковыми молекулами тиоэфирными свя-зями, образовавшимися в результате присоединения сульфгидрильных групп из остатков цистеина по винильным группам гема.
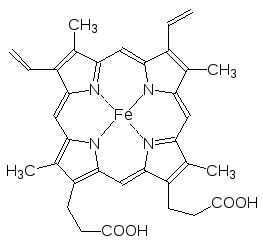
Содержащие гемовые структуры цитохромы оксидоредуктаз, окрашенные в цвета от красного до коричневого, участвуют в переносе электронов (Fe+2 Fe+3 + e), например, когда образовавшиеся в реакциях дегидриро-вания атомы водорода отдают соответствующим акцепторам свои элект-роны и превращаются в протоны. Цитохромы обозначают буквами латинского алфавита с цифровыми индексами, например а, а3, b, c1, с, f6, всего их известно более двадцати. В частности в цитохроме с феррипор-фириновый фрагмент в отличие от гема вместо остатков пропионовой кислоты содержит пропильные группы. Цитохромы участвуют также в реакциях окислительного гидроксилирования органических соединений, а пара цитохромоксидаз а-а3 переносит четыре электрона на молекулу кислорода (аэробные процессы).
Иногда в учебниках по биохимии в качестве универсального кофермента реакций гидрирования-дегидрирования рассматривается и убихинон (ubi – по латыни значит везде, то есть вездесущий хинон). На самом деле су-ществует несколько соединений этого класса, соответствующих формуле
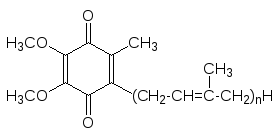
В частности убихинон с n = 10 выделен из печени млекопитающих (ко-фермент Q, CoQ), однако, это растворимое в клеточных липидах вещество не предназначено для связывания с белками и поэтому не может претен-довать на роль истинного кофермента. На самом деле небольшой пул этого соединения растворен в липидном слое клеточных мембран и служит для одно- или двухэлектронного переноса между более жестко встроенными в мембрану переносчиками электронов.
Фолиевая кислота (витамин Вс)
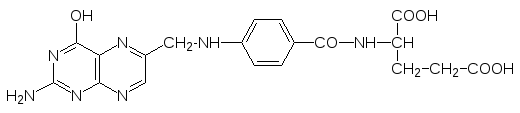
после ее превращения в тетрагидрофолиевую кислоту участвует в работе ферментов, переносящих одноуглеродные фрагменты (метильную, гидрок-симетильную, формильную группу):
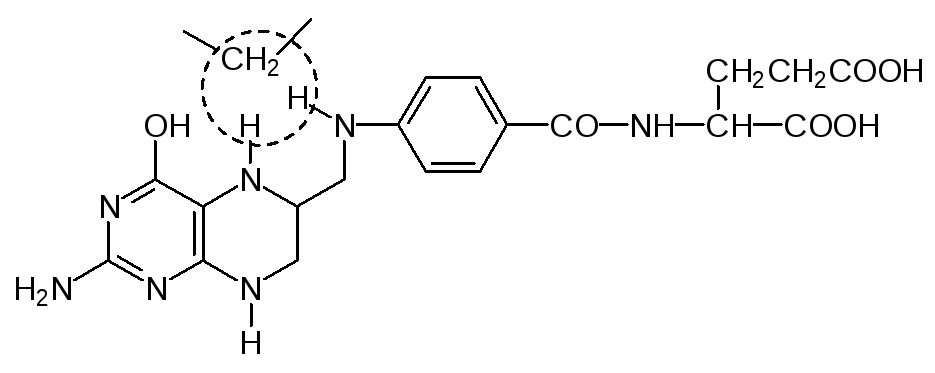
Среди ацилтрансфераз чаще всего встречаются ферменты, в работе кото-рых принимает участие кофермент А, обозначаемый часто CoA-SH, В этом соединении пирофосфатный фрагмент 3-фосфоаденозиндифосфата соеди-нен эфирной связью с пантотеновой кислотой, которая является обязатель-ным фактором питания, иногда ее называют также витамином В5.
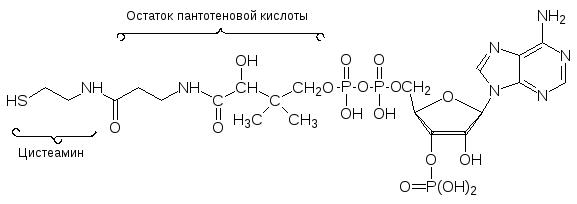
С участием АТФ или в результате обменной реакции с другим тиоэфиром это соединение ацилируется по сульфгидрильной группе с образованием соответствующего S‑ацильного производного, например ацетил-кофермен-та А (СоА-S-СОСН3) и снова может переносить ацильные группы на дру-гие молекулы по двум механизмам: с разрывом связей С-S- или С-Н в ‑положении к тиокарбоксилатной группе.
В группе лигаз одно из главных мест принадлежит карбоксилазам, у кото-рых простетическая группа представлена биотином или витамином Н:
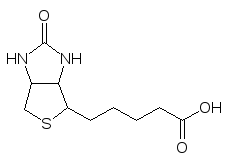
Витамин Н достаточно стабилен и авитаминоз по нему встречается только у людей, которые едят много сырых яиц, так как яйца содержат белок, который связывает биотин в прочный комплекс. Биотин присоединен к белковой составляющей фермента амидной связью за счет ацилирования концевой аминогруппы лизинового франмента в молекуле белка. Это связано с тем, что мочевинный участок бициклической системы простети-ческой группы должен перемещаться на таком поводке из десяти атомов с минимальной структурной перестройкой из одного участка фермента в другой, так как механизм его действия основан на двух реакциях. Сначала в одном участке каталитического центра фермента протекает его карбокси-лирование бикарбонатом с участием аденозинтрифосфата:

После этого уже в другом участке фермента карбоксильная группа перено-сится на другую молекулу, например, так образуется метилмалонил-СоА:
Одним из наиболее изучаемых витаминов является витамин С – аскорби-новая кислота, однако ее роль в функционировании ферментов до сих пор не установлена во всех ее проявлениях. Предполагается, что она участвует в образовании входящих в состав коллагена (белок соединительной ткани) аминокислот 4-гидроксипролина и 5-гидроксилизина и поэтому ее присут-ствие важно для заживления ран. Не исключено и ее участие в ассимиля-ции неорганического железа.
Сложное строение отличает группу веществ, называемых витамин В12, чаще всего это цианкобаламин.
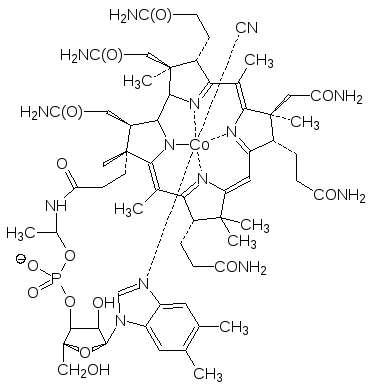
Витамин В12 представляет собой кобальторганическое соединение, раствор которого в воде окрашен в пурпурный цвет. Он выполняет роль кофермен-та изомераз, функционирующих по свободнорадикальному механизму. Ис-точниками кобаламина являются бактерии, в частности бактериальная флора пищеварительного тракта; в растениях он отсутствует. Наиболее ярко авитаминоз по этому фактору проявляется в нарушении процесса кроветворения.
Из других обязательных факторов питания можно назвать витамин А или ретинол, витамин Е и витамины группы D и группы К. Витамин А в орга-низме выполняет несколько функций. Наиболее известна его роль в зри-тельном процессе, где он в виде ретиналя, образующегося из ретинола,

участвует в образовании реагирующего на свет пигмента родопсина. Но дефицит по витамину А не ограничивается только «куриной слепотой», это всего лишь первый симптом авитаминоза по этому веществу. При серьез-ной недостаточности по витамину А появляется сухость кожи, глаз (ксеро-фтальмия), задержка развития и роста, стерильность самцов. Но и гиперви-таминоз опасен, особенно для беременных женщин.
Витамин Е или токоферол (от гр. токос – потомство, феро – несу)
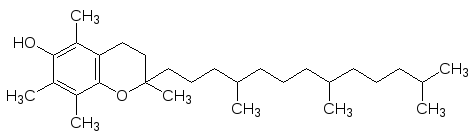
не участвует в работе ферментов, он является ингибитором свободнора-дикальных реакций. Его недостаток в рационе вызывает перерождение ре-продуктивных органов, нарушение функций мембран, дегенерацию пече-ни, но точная биологическая роль витамина Е не установлена. Много этого витамина содержится в растительном масле, тем более, что синтетический витамин Е добавляют сейчас в пищевые продукты специально для замед-ления процессов полимеризации и окисления.
Витамины группы D (они, как отмечалось выше, образуются из эргосте-рина) относятся к гормонам, так как они регулируют обмен кальция и фос-фата
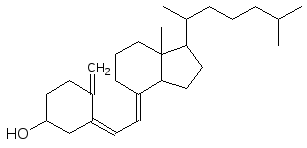 ,
,
а витамин К – замещенный метилнафтохинон – участвует в работе нес-кольких факторов свертывания крови, правда, коферментная роль его не установлена.
Кинетическое описание ферментативных реакций было предложено Миха-элисом и Ментен. В 1913 году эти исследователи предположили, что фер-мент (Е) и субстрат (S) образуют комплекс (ES, комплекс Михаэлиса),
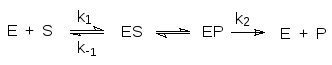
который превращается в комплекс фермента с продуктом превращения субстрата (ЕР), распадающийся далее на фермент и продукт (Р).
Основа катализа – снижение энергии активации реакции, и ферменты по способности снижать ее превосходят все известные катализаторы. Так, на-пример, разложение пероксида водорода без катализатора требует энергии активации 18000 кал/моль, самый активный неорганический катализатор – коллоидная платина – снижает ее до 11700 кал/моль, а фермент каталаза обеспечивает снижение энергии активации до менее 2000 кал/моль. Этило-вый эфир масляной кислоты гидролизуется в присутствии кислот с энерги-ей активации (Е) 16800 кал/моль, в присутствии щелочей 10200 кал/моль, а панкреатическая липаза (пищеварительный фермент, секретируемый поджелудочной железой) характеризуется энергией активации по этому субстрату равной 4500 кал/моль. Очень медленно гидролизуется водой са-хароза, в присутствии кислот этот процесс ускоряется (Е=25600 кал/моль), а дрожжевая инвертаза снижает этот показатель реакции до 8000-10000 кал/моль. Эта различие в абсолютных величинах выглядит не очень серь-езным, но энергия активации в уравнении Аррениуса входит в экспонен-циальный множитель
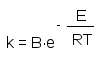
и тогда константа скорости ферментативной реакции (k с индексом е) в логарифмическом виде представляется уравнением
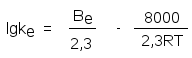 ,
,
а константа скорости реакции, катализируемой протонами (индекс h), уравнением
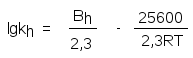
Постоянные реакций Вe и Вh примерно равны и тогда для температуры 37С получаем
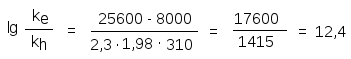
что соответствует соотношению ke/kh = 2,51012 (двенадцать порядков!).
Что же лежит в основе уникальной избирательности и высокой каталити-ческой активности ферментов? Прежде всего, это особенность пространст-венного строения белковой молекулы, лежащей в основе этих катализа-торов. Форма глобулярного белка во многом определяется гидрофобными взаимодействиями между структурными элементами входящих в его со-став аминокислот. Эти взаимодействия слабы, но когда их много – они дают серьезный выигрыш в энергии. Гидрофобные структурные элементы заполняют внутреннюю среду белковой глобулы, однако слабость каждого отдельного взаимодействия позволяет белку в ответ на воздействие на внешнюю сферу в результате изменения рН или присоединения другой мо-лекулы легко перестраиваться, принимая новую форму и меняя местами различные функциональные группы боковых цепей аминокислотных фраг-ментов. Еще Э.Фишер выдвинул гипотезу о взаимодействии субстрата с активным центром фермента по принципу «ключ-замок». Очень часто и сейчас используют принцип «ключ-замок» для схематического представ-ления ферментативного катализа, но этот подход слишком упрощен, он не отражает того, что в ответ на связывание с субстратом происходит пере-стройка белковой молекулы фермента, и тогда образующемуся комплексу соответствует минимум энергии уже при ином расположении составляю-щих фермент структурных элементов, а по мере протекания ферментатив-ной реакции такие перестройки белковой молекулы могут происходить неоднократно до отхода продукта реакции от реакционного центра.
В разделе, посвященном строению белков, уже были представлены данные по миоглобину и рибонуклеазе. В дополнение к ним можно привести при-мер защитного белка с гидролазной активностью лизоцима (лизис – раз-рыв, растворение). Этот фермент, в котором, как у большинства гидролаз, отсутствует простетическая группа, содержится в яичном белке, в слезах, в слюне, а его антимикробная активность основана на расщеплении олигоса-харидных фрагментов, участвующих в образовании клеточной стенки многих бактерий. В ‑структуры молекулы лизоцима включено 12 % ами-нокислотных составляющих, в -спирали – 40 %, а остальные образуют различные изгибы и иррегулярные витки. Активный центр лизоцима пред-ставлен только -спиралями. Расстояние, равное шагу -спирали (0,54 нм) или кратное ему, разделяет атомы с повышенной или с пониженной элект-ронной плотностью во многих биологически активных веществах.
Одной из главных особенностей ферментов является реализуемая с их по-мощью возможность использования свободной химической энергии, за-ключенной в таких макроэргических веществах как аденозинтрифосфат, для осуществления эндотермических реакций, а также синтез аденозин-трифосфата за счет использования энтропийного фактора. Однако, и без этого ферменты представляют собой уникальные катализаторы, специфи-ку которых объясняют перечисленными далее механизмами.