

10. Основные анаболические процессы
10.1. Глюконеогенез
Главным быстро мобилизуемым источником энергии в клетках является глюкоза, запасы которой находятся в виде продуктов ее поликонденсации крахмала, гликогена или других полисахаридов. В крови человека поддер-живается постоянный уровень глюкозы около 5 ммоль/л, но небольшие отклонения от этого нормального уровня возможны при интенсивной рабо-те или после еды. Глюкоза поступает в клетки по градиенту концентраций, но есть специальный мембранный белок, облегчающий ее транспорт. Уро-вень глюкозы в крови поддерживается с помощью гормона поджелудочной железы инсулина и ряда других гормонов. Инсулин имеет пептидную при-роду, он состоит из 51 аминокислоты, но секреторные клетки (островки Лангерганса) производят проинсулин, который у разных животных содер-жит от 78 до 86 аминокислотных компонент в виде одной цепочки с тремя дисульфидными связями. Активация с образованием самого инсулина про-исходит за счет гидролитического расщепления молекулы проинсулина с элиминированием из нее центрального участка (С‑цепь) с сохранением дисульфидных связей в активном двухцепочечном полипептиде (А- и В‑цепи). Инсулин не проходит через клеточную мембрану, но он связыва-ется с белком переносчиком глюкозы и ускоряет ее диффузию в клетку. При избытке глюкозы она превращается в гликоген, а при недостатке глю-козы она образуется из гликогена. При недостатке инсулина или при на-рушении механизма поступления глюкозы в клетки организм повышает ее концентрацию в крови для того, чтобы увеличить движущую силу диффу-зии за счет градиента концентраций. Однако, это приводит и к интенсив-ному выделению глюкозы почками (сахарный диабет). Испытывающие дефицит по глюкозе клетки начинают синтезировать ее из аминокислот сократительных белков, подключая к этому окислительные превращения жирных кислот. В результате этого мышечные ткани атрофируются и разрастается жировая ткань.
Для клеток нервной системы, почек, семенников, тканей эмбриона глюкоза является единственным поставщиком энергии. Важно, что для транспорта глюкозы в нейроны инсулин не нужен и мозг диабетиков, как и мозг здо-ровых людей, расходует около 120 г глюкозы в сутки.
Роль глюкозы в метаболизме очевидна, но возможности депонирования ее в виде гликогена ограничены: в организме взрослого человека содержится всего лишь несколько сот граммов этого полисахарида. Поступление глю-козы с пищей (в основном в виде крахмала) не всегда соответствует дейст-вительным потребностям и поэтому организм животных в значительной мере зависит от образования глюкозы из поступающих с пищей веществ. У теплокровных избыток пищевых веществ закладывается в жировую ткань, а при недостатке глюкозы в пище на ее биосинтез расходуются белки, по-скольку образующийся из жирных кислот ацетил-СоА не может превра-щаться в глюкозу. Ключевое вещество цикла Кребса – оксалоацетат – так-же не может быть получен из ацетилкофермента А, поэтому правило диетологии гласит: жиры сгорают в пламени углеводов.
Напомним три основных положения, относящиеся к течению метаболи-ческих процессов:
Катаболические и анаболические пути одного вещества различны. Это значит, что есть по крайней мере одна стадия, по которой они не совпадают.
Катаболический и анаболический путь превращения одного вещества контролируется различными регуляторными ферментами, но регуляция биосинтеза и биодеградации одного вещества взаимосвязаны. Актива-ция биосинтеза приводит к подавлению катаболического превращения.
Анаболические процессы расходуют АТФ, который поставляется ката-болическими. Поскольку КПД всегда менее 100 %, затраты АТФ оказы-ваются выше, чем это следует из термодинамических расчетов. Это обеспечивает необратимость метаболических превращений.
При изучении процесса гликолиза было установлено, что конечный про-дукт анаэробного превращения глюкозы – молочная кислота – может сно-ва превращаться в глюкозу. При этом около 1/5 молочной кислоты окис-ляется до диоксида углерода, поставляя энергию для глюконеогенеза.
В процессе гликолиза глюкозы образованию молочной кислоты предшест-вует катализируемое пируваткиназой образование пирувата из фосфоенол-пирувата с генерированием АТФ (АТР):
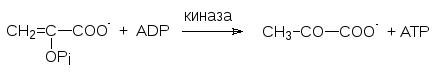
По энергетическим соображениям обращение этого пути невозможно, так как прямая реакция сопровождается выделением 7,5 ккал/моль. Поэтому для глюконеогенеза образование фосфоенолпирувата из пирувата вклю-чает обходной путь. Сначала пируват переносится в матрикс митохондрий и карбоксилируется биотинзависимым ферментом с образованием оксало-ацетата. Для переноса оксалоацетата в цитозоль, где идет биосинтез глю-козы, оксалоацетат восстанавливается в малат, для которого существует специальная транспортная система, и уже в цитозоле малат дегидрируется с образованием оксалоацетата. Анион щавелевоуксусной кислоты фосфо-рилируется действием ГТФ в присутствии фосфоенолпируваткарбокси-киназы с одновременным декарбоксилированием:
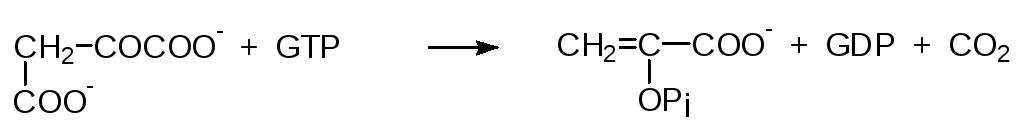
Хотя изменение стандартной свободной энергии при этом равно + 0,2 ккал/моль, в реальных условиях клетки этот процесс сопровождается выделением 6 ккал/моль, что делает его необратимым.
В обращение процессов гликолиза фосфоенолпируват гидратируется и восстанавливается до фосфоглицеринового альдегида, половина которого изомеризуется в фосфат диоксиацетона, после чего под действием альдо-лазы из фосфоглицеринового альдегида и фосфата диоксиацетона обра-зуется 1,6-дифосфат фруктозы. При катаболическом гликолизе идет фос-форилирование глюкозы и фосфофруктозы аденозинтрифосфатом, но об-ратный процесс образования АТФ из дифосфата фруктозы или фосфо-глюкозы и АДФ по термодинамическим соображениям невозможен, поэто-му при глюконеогенезе фосфатные группы отщепляются гидролитически.
Если суммарный процесс гликолиза представляется схемой
![]() ,
,
то на обратный процесс затрачивается уже втрое больше АТФ, чем обра-зуется в прямом (поэтому 15 % молочной кислоты идет на энергетическое обеспечение глюконеогенеза):
![]()
В начале глюконеогенеза лежит пируваткарбоксилаза – аллостерический фермент, активируемый ацетилкоферментом А, то есть при накоплении в клетке неиспользуемого ацетилкофермента А начинается усиленный био-синтез глюкозы из пирувата.
В глюкозу и далее в гликоген могут превращаться все промежуточные продукты цикла Кребса. Большинство аминокислот также относятся к глю-когенным. В их числе аланин, глютаминовая, аспарагиновая кислота и их амиды, превращающиеся (амиды после гидролиза) трансаминазами в пи-руват, кетоглутарат и оксалоацетат, кроме того, эти кетокислоты образу-ются и из других аминокислот. Особенно интенсивно идет превращение аминокислот в глюкозу при недостатке инсулина.