

Глава 5 экология популяций (демэкология) 105
щшшш, 137
ПРИКЛАДНАЯ ЭКОЛОГИЯ 160
Продуценты — автотрофные организмы, способные про- В изводить органические вещества из неорганических, I используя фотосинтез или хемосинтез (растения и авто- : щ; трофные бактерии). I
Консументы (макроконсументы, фаготрофы) — гетеро- ш трофные организмы, потребляющие органическое веще- І ство продуцентов или других консументов (животные, * гетеротрофные растения, некоторые микроорганизмы). I Консументы бывают первого порядка (фитофаги, сапро- 1 фаги), второго порядка (зоофаги, некрофаги) и т.д. 1
Редуценты (микроконсументы, деструкторы, сапротрофы, ® осмотрофы) — гетеротрофные организмы, питающиеся I; органическими остатками и разлагающие их до минераль- I ных веществ (сапротрофные бактерии и грибы). 1
БИОСФЕРА 24
т 44
т 44
т 44
т 44
г 44
т 44
т 44
т 44
т 44
т 44
*жтт 86
ОБЩАЯ ЭКОЛОГИЯ 86
ЭКОЛОГИЯ ОРГАНИЗМОВ (АУТЭКОЛОГИЯ) 86
др-); 89
Глава 5 экология популяций (демэкология) 105
щшшш, 137
ПРИКЛАДНАЯ ЭКОЛОГИЯ 160
 В
экосистеме пищевые и энергетические
связи идут в направлении: продуценты
-> консументы -» редуценты.
В
экосистеме пищевые и энергетические
связи идут в направлении: продуценты
-> консументы -» редуценты.
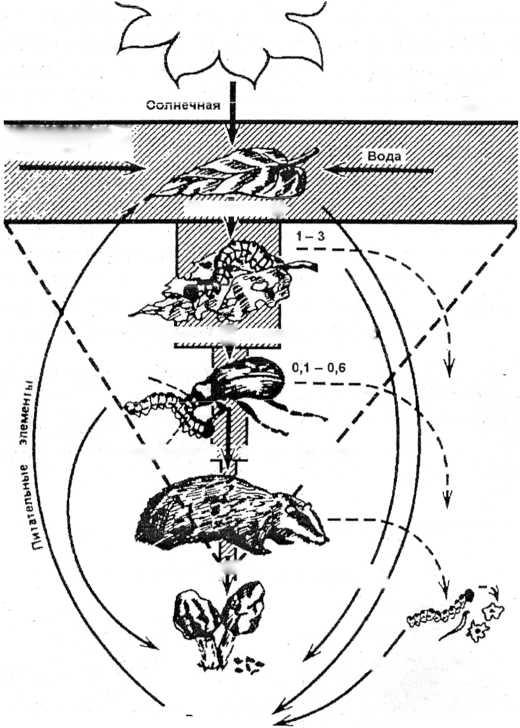
Пищевые цепи и сети. Питаясь друг другом, живые организмы образуют цепи питания. Цепь питания — последовательность организмов, по которой передается энергия, заключенная в пище, от ее первоначального источника. Каждое звено цепи называется трофическим уровнем (рис. 19). Первый трофический уровень — продуценты (автотрофные организмы,
энергия
щшшш,
^ Углекислый газ
Продуценты^/
Консументы I .^порядка (фитофаги;
Паразиты
фитофагов
Консументы
II
01 - 0,1
Паразиты
хищников
Консументы III р&Гпорядка (вторичные хищники) / J
/ Паразиты хищников высоких порядков и надпаразиты
преимущественно зеленые растения). Второй трофический уровень — консументы первого порядка (растительноядные животные и паразиты продуцентов). Третий трофический уровень — консументы второго порядка (первичные хищники, питающиеся растительноядными животными, и паразиты первичных консументов). Четвертый трофический уровень — консументы третьего порядка (вторичные хищники, питающиеся плотоядными животными, и паразиты вторичных консументов). В пищевой цепи редко бывает больше 4—5 трофических уровней. Последний трофический уровень — редуценты (сапротрофные бактерии и грибы). Они осуществляют минерализацию — превращение органических остатков в неорганические вещества. Редуценты могут представлять любой трофический уровень, начиная со второго.
Различают два типа пищевых цепей (рис. 20). Цепи выедания (или пастбищные) — пищевые цепи, начинающиеся с живых фотосинтезирующих организмов. Например, фитопланктон —> зоопланктон —» рыбы микрофаги -» рыбы макрофаги -> птицы ихтиофаги. Цепи разложения (или детритные) — пищевые цепи, начинающиеся с отмерших остатков растений, трупов и экскрементов животных. Например, детрит —> детрито- фаги —> хищники микрофаги —> хищники макрофаги. Таким
Пастбищная
пищевая цепь
Детритная
пищевая цепь
Рис.
20.
Ү-образная
модель
потока энергии, показывающая связь
между пастбищной и детритной пищевыми
цепями (Ю. Одум, 1986)
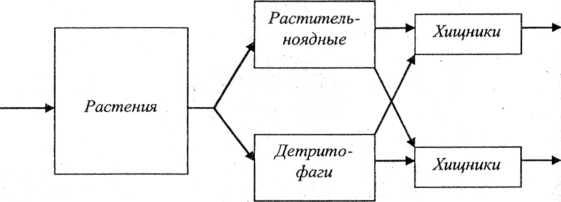
образом, поток энергии, проходящий через экосистему, разбивается как бы на два основных направления. Энергия к консу- ментам поступает через живые ткани растений или через запасы мертвого органического вещества. Цепи выедания преобладают в водных экосистемах, цепи разложения — в экосистемах суши.
В сообществах пищевые цепи сложным образом переплетаются и образуют пищевые сети. В состав пищи каждого вида входит обычно не один, а несколько видов, каждый из которых в свою очередь может служить пищей нескольким видам. С одной стороны, каждый трофический уровень представлен многими популяциями разных видов, с другой стороны, многие популяции принадлежат сразу к нескольким трофическим уровням. В результате благодаря сложности пищевых связей выпадение какого-то одного вида часто не нарушает равновесия в экосистеме.
Круговорот веществ и поток энергии в экосистеме. В экосистеме органические вещества синтезируются автотрофами из неорганических веществ. Затем они потребляются гетеротро- фами. Выделенные в процессе жизнедеятельности или после гибели организмов (как автотрофов, так и гетеротрофов) органические вещества подвергаются минерализации, то есть превращению в неорганические вещества. Эти неорганические вещества могут быть вновь использованы автотрофами для синтеза органических веществ. Так осуществляется биологический круговорот веществ.
В то же время, энергия не может циркулировать в пределах экосистемы. Поток энергии (передача энергии), заключенной в пище, в экосистеме осуществляется однонаправленно от автотрофов к гетеротрофам.
На рис. 21 представлена упрощенная схема потоков энергии на трех трофических уровнях пищевой цепи. Трофические уровни изображены как резервуары, размер которых соответствует энергии, заключенной в их биомассе (В), размер соединяющих их каналов — величине потоков энергии. На входе в систему поступает поток солнечной энергии (L). На первом трофическом уровне зелеными растениями поглощается около 50% солнечной энергии (Ьа). Только часть ее в процессе
g
й
1
ЈJS
X
tt>°S
5
л
у 3
^
^ сх; ^
*
£
1 “§ ж
к р .
р*
і>>
^
' • 3 s
Н
a
S
^
р
оЗ
о Н СХ Н
о,
g
а,
о гч
HfflOC
1
I I 1
Ч
»5
«-, й
►« 8 «
« £ - "
§§
^
8
* к з
§
,
О
! * !
CQ
ё
£ &S
a
«
g
g.
аЗ
$ S
СХ
^
g
I §
g
§So|^s
1
«-£§
ft*
§
I
s
a
“ Ј
о
„ «Я о О °
а
°ч я к с »
ЧАС'1'b
II.
ОБЩАЯ
ЭКОЛОГИЯ
У
О и-Ө"
О
а>
X
</
лЧ
Оч
О
сх
^
03
оЗ
«л»
а > с-
£2^^
>» о
.
й
И
S
О*
“
0,0
§ с
L
П
Һ
*
,
«
«
оо
а\
§
>,
к
S
<D
i—
1
О
оЗ
о
сх
2
аЗ
X
р,
Er<
So4
с
ftVl
сс
£
о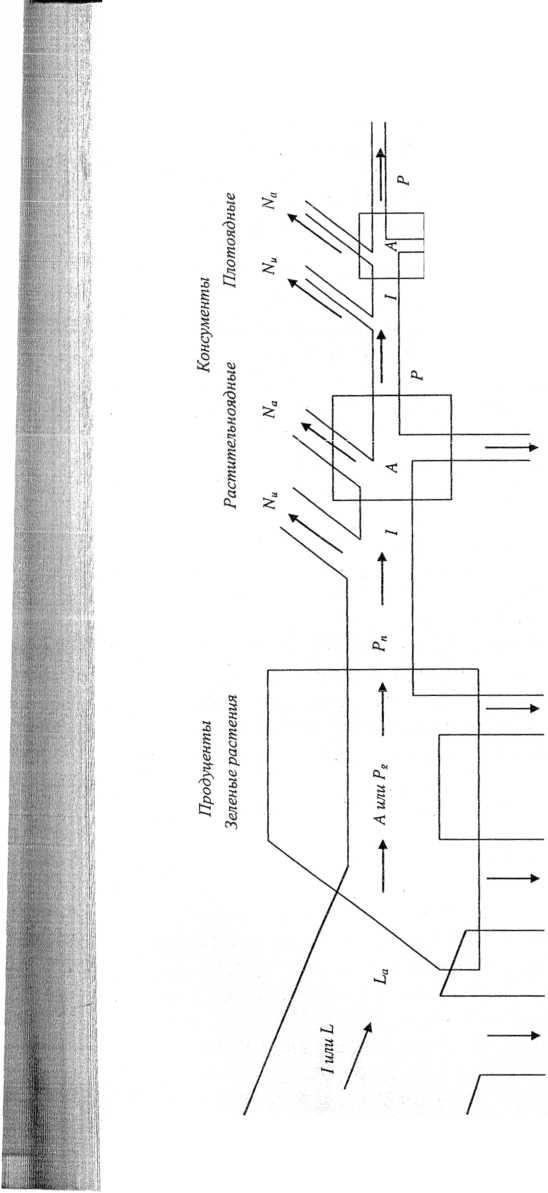
фотосинтеза преобразуется в энергию химических связей органических веществ. Это валовая первичная продукция (Р ). Большая часть поглощенной растениями, но не усвоенной энергии, рассеивается в окружающую среду в виде тепловой энергии. Часть образованных органических веществ окисляется, а высвобождающаяся энергия расходуется на поддержание всех метаболических процессов. Это так называемые траты на дыхание (R). Эта энергия, в конечном счете, также рассеивается в виде тепла. Оставшаяся часть новообразованных органических веществ составляет прирост биомассы растений (АВ) и называется чистой первичной продукцией (PJ. В чистую первичную продукцию превращается только 1% поглощенной растением энергии. Таким образом, Рп= Р — R.
До второго трофического уровня доходит только часть чистой первичной продукции. Некоторая ее часть (Nu) не используется консументами первого порядка. Она может накапливаться или экспортироваться за пределы системы. Та часть, которую ассимилировали (потребили) консументы (А), частично тратится на дыхание (R), частично выделяется с экскрементами (JV), а остальное накапливается в виде вторичной продукции (Р).
Вторичная продукция на каждом последующем трофическом уровне консументов (Р2, Р3 и т.д.) составляет около 10% предыдущей (хотя на уровне хищников может быть выше — около 20%). В результате, чем длиннее пищевая цепь, тем меньше остается к ее концу накопленной в органическом веществе энергии. Поэтому число трофических уровней никогда не бывает слишком большим.
Таким образом, при передаче энергии с одного трофического уровня на другой большая часть энергии рассеивается в виде тепла (в соответствии со вторым законом термодинамики), и только около 10% от первоначального количества передается по пищевой цепи.
В результате, пищевые цепи можно представить в виде экологических пирамид. Различают три основных типа экологических пирамид (рис. 22).
Пирамида чисел (пирамида Элтона) отражает уменьшение численности организмов от продуцентов к консументам.
2
х Ю'
Телята
Люцерна
10
1
4,5
Мальчик
Телята
Люцерна
|
|
|
— 1 Мальчик 48 кг ] |
|
|
|
Телята 1035 кг | ||
|
Люуе |
пип 8211 кг | ||
1 10 100 I I !
10
100
_|
б
Ткани человека 34,75 кДж
4,98x10 кДж
6,24x10 кДж
| 7 1 1 Использованная солнечная энергия 1 ? _'
100
Рис. 22. Пирамиды чисел (а), биомасс (б) и энергии (в), представляющие упрощенную экосистему: люцерна — телята — мальчик 12 лет (по Ю. Одуму, 1959)
Пирамида чисел (а) показывает, что если бы мальчик питался в течение одного года только телятиной, то для этого ему потребовалось бы 4,5 теленка, а для пропитания телят необходимо засеять поле в 4 га люцерной, что составит 2 х Ю7 растений. В пирамиде биомасс (б) число особей заменено их биомассой. В пирамиде энергии (в) учтена солнечная энергия. Люцерна использует 0,24% солнечной энергии. Для накопления продукт! и телятами в течение года используется 8% энергии, аккумулированной люцерной. На развитие и рост ребенка в течение года используется 0,7% энергии, аккумулированной телятами. В результате чуть более одной миллионной доли солнечной энергии, падающей на поле в 4 га, используется для пропитания ребенка в течение одного года.
Пирамида биомасс показывает изменение биомасс на каждом следующем трофическом уровне: для наземных экосистем пирамида биомасс сужается кверху, для экосистемы океана — имеет перевернутый характер (сужается книзу), что связано с быстрым потреблением фитопланктона консументами.
Пирамида энергии (продукции) имеет универсальный характер и отражает уменьшение количества энергии, содержащейся в продукции, создаваемой на каждом следующем трофическом уровне.
Биологическая продуктивность экосистем
І Прирост биомассы в экосистеме, созданной за единицу времени, называется биологической продукцией (продуктивностью). Различают первичную и вторичную продукцию сообщества.
Первичная продукция — биомасса, созданная за единицу времени продуцентами. Она делится на валовую и чистую. Валовая первичная продукция (общая ассимиляция) — это общая 1 биомасса, созданная растениями в ходе фотосинтеза. Часть ее расходуется на поддержание жизнедеятельности растений — траты на дыхание (40—70 %). Оставшаяся часть составляет чистую первичную продукцию (чистая ассимиляция), которая в |ч дальнейшем используется консументами и редуцентами, или 1 накапливается в экосистеме.
Вторичная продукция — биомасса, созданная за единицу времени консументами. Она различна для каждого следующе- I' го трофического уровня.
Масса организмов определенной группы (продуцентов, кон- I сументов, редуцентов) или сообщества в целом называется био- |. массой. Самой высокой биомассой и продуктивностью обла- Р дают тропические дождевые леса, самой низкой — пустыни и Һ тундры (табл. 17).
Если в экосистеме скорость прироста растений (образова- рния первичной продукции) выше темпов переработки ее кон- Всументами и редуцентами, то это ведет к увеличению биомас- сы продуцентов. Если при этом присутствует недостаточная J утилизация продуктов опада в цепях разложения, то происхо- | дит накопление мертвого органического вещества. Это прояв-
|
Тип экосистем |
Площадь, 106 км |
Биомасса растений, кг/м2 |
Мировая величина | ||
|
колеба ния |
в среднем |
Биомасса растений, 109т |
Биомасса животных, 106 т | ||
|
Влажные тропические леса |
17,0 |
6-80 |
45 |
765 |
330 |
|
Тропические сезоннозеленые леса |
7,5 |
6-80 |
35 |
260 |
90 |
|
Вечнозеленые леса умеренного пояса |
5,0 |
6-200 |
35 |
175 |
50 |
|
Листопадные леса умеренного пояса |
7,0 |
6-60 |
30 |
2)0 |
110 |
|
Тайга |
12,0 |
6-40 |
20 |
240 |
57 |
|
Лесо-кустарниковые сообщества |
8,5 |
2-20 |
6 |
50 |
40 |
|
Саванна |
15,0 |
0,2-15 |
4 |
60 |
220 |
|
Лугостепь |
9,0 |
0,2-5 |
1,6 |
14 |
60 |
|
Тундра и высокогорье |
8,0 |
0,1-3 |
0,6 |
5 |
35 |
|
Пустыни и полупустыни |
18,0 |
0,1-4 |
0,7 |
13 |
8 |
|
Сухие пустыни, скалы, ледники и т. п. |
24,0 |
0-0,2 |
0,02 |
0,5 |
0,02 |
|
Культивируемые земли |
14,0 |
0,4-12 |
1 |
14 |
6 |
|
Болота и марши |
2,0 |
3-50 |
15 |
30 |
20 |
|
Озера и водотоки |
2,0 |
0,01 |
0,02 |
0,05 |
10 |
|
Материковые экосистемы в целом |
149 |
- |
12,3 |
1837 |
1005 |
|
Открытый океан |
332,0 |
0-0,005 |
0,003 |
1,0 |
800 |
|
Зоны апвеллинга |
0,4 |
0,005-0,1 |
0,02 |
0,008 |
4 |
|
Континентальный шельф |
26,6 |
0,001- 0,04 |
0,01 |
0,27 |
160 |
|
Заросли водорослей и рифы |
0,6 |
0,04-4 |
2 |
1,2 |
12 |
|
Эстуарии |
1,4 |
0,01-6 |
1 |
1,4 |
21 |
|
Морские экосистемы в целом |
361 |
- |
0,01 |
3,9 |
997 |
|
Общая биомасса Земли |
510 |
- |
3,6 |
1841 |
2002 |
Таблица 17
Биомасса разных типов экосистем
(Н.Ф. Реймерс, 1990)
ляется в заторфовывании болот, образовании мощной лесной подстилки и т.п. В стабильных экосистемах биомасса остается постоянной, так как практически вся продукция расходуется в цепях питания.
Динамика экосистем
Изменения в сообществах могут быть циклическими и поступательными.
Циклические изменения — периодические изменения в биоценозе (суточные, сезонные, многолетние), при которых биоценоз возвращается к исходному состоянию.
Суточные циклы связаны с изменением освещенности, температуры, влажности и других экологических факторов в I течение суток. Суточные ритмы проявляются в изменении со- : стояния и активности живых организмов в течение суток. Например, чередование световой и темновой фаз фотосинтеза у растений, сна и бодрствования у животных.
I Сезонная цикличность связана с изменением экологических факторов в течение года и наиболее сильно выражена в высоких широтах, где велик контраст зимы и лета. Сезонная из- 1 менчивость проявляется не только в изменении состояния и активности, но и количественного соотношения отдельных I видов. На определенный период многие виды выключаются ; из жизни сообщества, впадая в спячку, оцепенение, перекочевывая или улетая в другие районы.
Многолетняя изменчивость связана с флуктуациями климата или другими внешними факторами (степень разлива рек), I либо с внутренними причинами (особенности жизненного | цикла растений-эдификаторов, повторения массового размножения животных).
■Е; Поступательные изменения — изменения в биоценозе, в конечном счете приводящие к смене этого сообщества дру- ! гам. Сукцессия — последовательная смена биоценозов (экоси- I стем), выраженная в изменении видового состава и структуры І сообщества (рис. 23). Последовательный ряд сменяющих друг друга в сукцессии сообществ называется сущессионной серией.
I К сукцессиям относятся опустынивание степей, зарастание озер 1 и образование болот и др.