

1. Клітини рослинних організмів
Цитологія
(від гр. κύτος – «вмістилище», тут:
«клітина» і λόγος – «вчення», «наука»)
Цитологія– розділ біології, який вивчає живі клітини, їх походження, їхню будову, функціонування, процеси клітинного розмноження, старіння и смерті.
Клітина – це найменша структурна та функціональна одиниця живого організму. Усі клітини мають спільне походження, схожу будову і подібне функціонування (в них відбуваються схожі процеси). Усі клітини утворюються в результаті поділу материнської (вихідної) клітини. Із материнської клітини спадкова інформація (програма функціонування клітин і всього організму в цілому) передається у знов утворені дочірні клітини.
Хімічний склад клітини (біологічних об’єктів). Клітина, з фізичної точки зору, є одним з рівнів організації матерії, який включає в себе атомарний і молекулярний рівні, і складається з хімічних елементів. За співвідношенням хімічних елементів та їх ізотопним складом біологічні об’єкти (клітини організмів) відрізняються від об’єктів неживої природи (земної кори). Умовно усі елементи клітини (організму), за масовою часткою, можна розділити на три групи:
макроелементи(> 0,001%): кисень (О) – 65-75%, вуглець (С) – 15-18%, водень (Н) – 8-10%, азот (N) – 2,0-3,0%, фосфор (Р) – 0,2-1,0%, калій (К) – 0,15-0,40%, сірка (S) – 0,15-0,20%, хлор (Cl) – 0,05-0,10%, магній (Mg) – 0,02-0,03%, натрій (Na) – 0,02-0,03%, кальцій (Са) – 0,04-2,00%, залізо (Fe) – 0,01-0,015%;
мікроелементи (0,001% - 0,000001%): ванадій (V), германій (Ge), йод (І), кобальт (Со), марганець (Mn), нікель (Ni), рутеній (Ru), селен (Se), фтор (F), мідь (Cu), хром (Cr), цинк (Zn);
ультрамікроелементи (<0,000001%): золото (Au), срібло (Ag), ртуть (Pb), платина (Pt), цезій (Cs).
М
Силове
поле у фізиці – це векторне поле у
просторі, у кожній точці якого на пробну
частку діє певна за величиною і напрямом
сила (вектор сили).
Організм
(пізнолат. organis-mus
від пізнолат. organizo –
засвоюю, створюю упорядкований вид,
від давньогр. ὄργανον – знаряддя) –
живе тіло, властивості якого відмінні
від не живої матерії.
Програма
(від гр. program-ma – об’ява, розпорядження;
алгоритм дій)
Утворені у певних силових (енергетично-інформаційних) полях простору молекули, за програмами більш високих рівнів, об’єднуються у інформаційні самовідновні структури – організми. (біологічні об’єкти) Клітини організму, як відмічалось раніше, представляють собою програмовані (керовані) структурно-функціональні одиниці організмів. У своїй будові вони складаються з молекулярних спеціалізованих комплексів – органел клітини, які функціонально об’єднані між собою обміном речовин.
Молекули, які організують (будують) внутрішню самовідновну структуру органел клітин, називаються конституційними. Молекули які синтезуються живими організмами у процесі життєдіяльності і які забезпечують взаємодію органел клітин між собою і навколишнім середовищем, називаються ергастичними.
Конституційні молекули виконують будівельну та інформаційно-транскрипційну функції, у той час як ергастичні речовини виконують інформаційно-трансляційну, захисну, запасну, енергетично-транспортну та інші функції, які забезпечують існування (функціонування) клітин.
Ж
Транскрипція
(від лат. transcriptio – переписування)
запис програми (інформації) на мобільні
молекули-носії
Трансляція
(від лат. trans-latio – передача) перенесення
програми (інформації) від місця збереження
до місця її виконання
Хімічні сполуки, що утворюються поза межами організмів називаються мінеральними (неорганічними). Вуглецевмісні сполуки, синтезовані організмами із неорганічних сполук називаються органічними.
На молекулярному рівні біологічні об’єкти (клітини) складаються з обох типів сполук, які у своїх співвідношеннях дуже варіюють (змінюються):
неорганічні сполуки: вода (Н2О) – 70-80%, мінеральні солі – 1,0-1,5%;
органічні сполуки: білки – 10-20% (до 35%), ліпіди – 2,0-5,0% (до 50-90%), вуглеводи – 1,0-5,0% (до 70%), нуклеїнові кислоти – 1,0-5,0%, інші речовини – 0,1-0,5%.
О
Метаболізм
(від гр. μεταβολή – «перетворення,
зміна»)
Анаболізм
(від гр. ἀναβολή – «підйом») або
пластичний обмін
Катаболізм
(від гр. καταβολή – «основа») або
енергетичний обмін
анаболізм (анаболістичні реакції) – реакції синтезу (утворення) складних речовин із простих, які відбуваються з поглинанням енергії (ендогенні реакції),
катаболізм (катаболістичні реакції) – реакції розщеплення складних речовин на прості, які відбуваються з виділенням енергії (екзогенні реакції).
Будова рослинної клітини. Функціонально рослинну клітину можна розділити на дві складові частини (рис. 1.):
протопласт – «жива» конституційна частина, яка зберігає інформацію і структуру клітини.
похідні протопласту – «нежива» ергастична частина, через яку інформація клітини реалізується (виконується).
|
|
|
Рослинна клітина |
|
| |||||||||||||||||||||||||||||||
|
|
|
|
| ||||||||||||||||||||||||||||||||
|
|
|
|
|
| |||||||||||||||||||||||||||||||
|
|
Протопласт |
|
Похідні протопласту | ||||||||||||||||||||||||||||||||
|
|
|
|
|
|
| ||||||||||||||||||||||||||||||
|
|
|
|
|
|
| ||||||||||||||||||||||||||||||
|
Плазмалема |
|
Цитоплазма |
|
Органела Ядро |
|
|
Клітинна стінка | ||||||||||||||||||||||||||||
|
| |||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
| ||||||||||||||||||||||||||||
|
|
|
Гіалоплазма |
|
Органели |
|
Клітинний сік вакуоль | |||||||||||||||||||||||||||||
|
|
| ||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
| ||||||||||||||||||||||||||||
|
|
Ендоплазматична сітка |
|
|
Рибосоми |
|
|
| ||||||||||||||||||||||||||||
|
|
|
|
Речовини запасу | ||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
| ||||||||||||||||||||||||||||
|
|
Апарат Гольджі |
|
|
Мікротрубочки |
|
|
| ||||||||||||||||||||||||||||
|
|
|
|
|
Фізіологічно активні речовини | |||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
| |||||||||||||||||||||||||||||
|
Вакуолі |
|
|
Мікрофіламенти |
|
| ||||||||||||||||||||||||||||||
|
|
|
|
|
| |||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
| ||||||||||||||||||||||||||||
|
Пластиди |
|
|
Мітохондрії |
|
|
| |||||||||||||||||||||||||||||
|
|
|
| |||||||||||||||||||||||||||||||||
Рис. 1. Структурний взаємозв’язок компонентів клітини
Протопласт і його складові частини
П
Протопласт
(від гр. πρῶτος (protos) – перший і
πλαστός (plastos) –
виліплений, утворений)
Плазмалема
(від гр. πλάσμα (plásma) – буквально
виліплене, оформлене і (lémma) –
оболонка, шкірка)
Цитоплазма
(від гр. κύτος (kytos) –
вмістилище, тут – клітина і πλάσμα
(plasma) –
виліплене, оформлене)
Гіалоплазма
(від гр. (hyalinos) –
скловидний, прозорий і πλάσμα (plasma) –
виліплене, оформлене)
Цитозоль
(від гр. κύτος (kytos) –
вмістилище, тут – клітина і англ.
sol від лат. solutio – розчин)
Мезоплазма
від гр. μέσος –
середній і πλάσμα (plásma) – буквально
виліплене, оформлене)
Плазмалема (плазматична мембрана) – зовнішня мембрана протопласту, що оточує цитоплазму. За суттю плазмалема це мембранний мішок заповнений цитоплазмою. Плазмалема приймає участь у обміну речовин між клітиною і зовнішнім середовищем.
Цитоплазма –позаядерна внутрішня частина протопласту клітини. Складається із гіалоплазми, у якій містяться органели та інші включення. Цитоплазма здатна до росту і відтворення, при частковому видаленні може відновлюватись. Однак нормально функціонує цитоплазма тільки в присутності ядра. Без нього довго існувати цитоплазма не може, як і ядро без цитоплазми.
Гіалоплазма (цитозоль, мезоплазма) – прозора частина цитоплазми у якій розташовані внутріклітинні структури – органели (включно з ядром клітини), включення, фізіологічно активні речовини (продукти метаболізму). Гіалоплазма постійно рухається, перетікає всередині живої клітини, переміщуючи разам із собою різні речовини, включення і органоїди (органели). Цей рух називається циклозом і генерується мікрофіламентами. Гіалоплазма об’єднує всі клітинні структури (компоненти) і забезпечує їх хімічну взаємодію.
|
|
|
Лейкопласт |
|
|
Плазмалема | |
|
Гіалоплазма
|
Апарат Гольджі | |
|
Ядро |
Рибосоми | |
|
Агранулярна ЕПС |
Центріоль | |
|
Плазмодесма |
Мітохондрія | |
|
Скорочувальна вакуоля |
Хлоропласт | |
|
Тонопласт |
Хромопласт | |
|
|
Клітинний сік |
 Гранулярна
ЕПС
Гранулярна
ЕПС










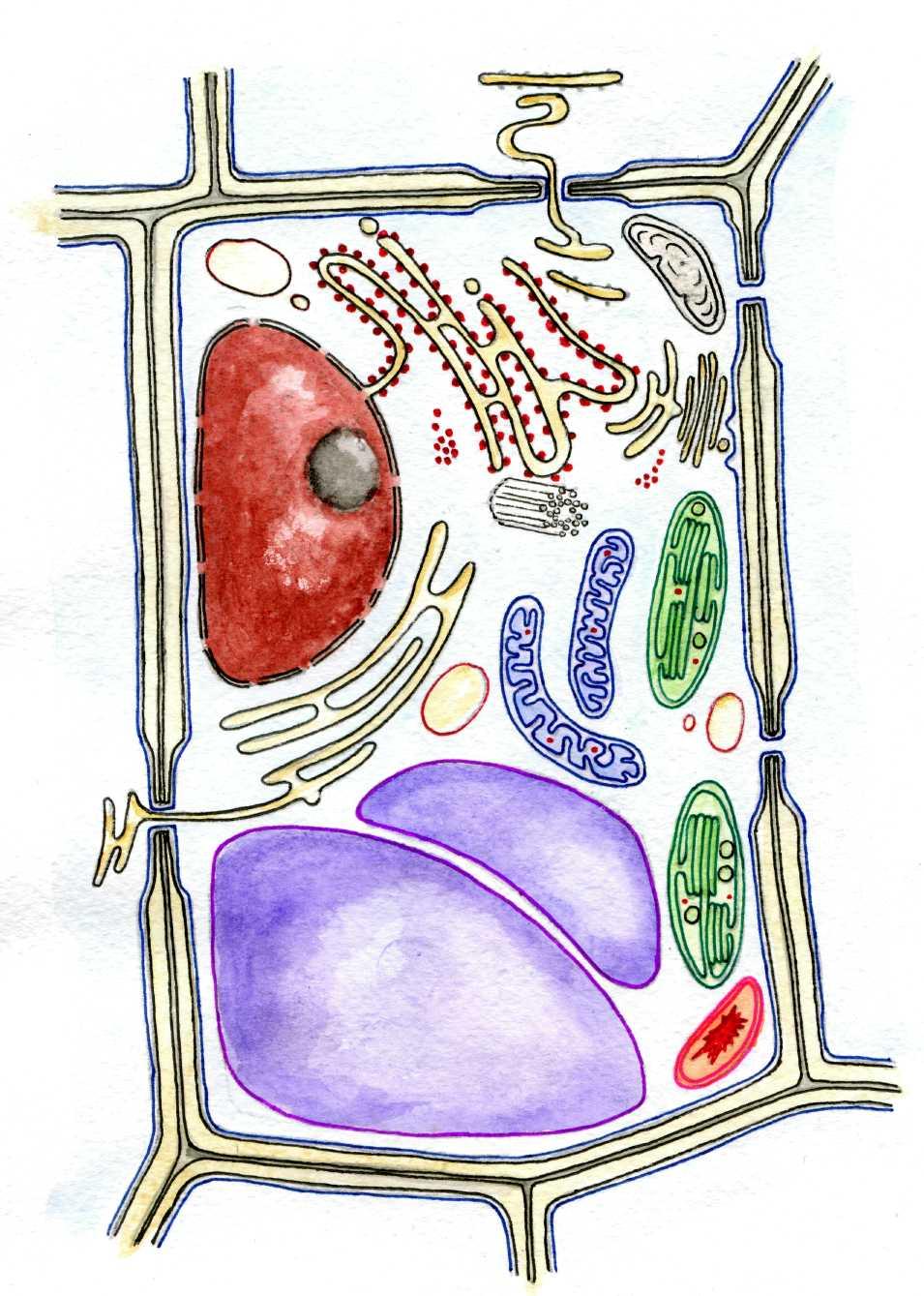


 Вакуоля
сферосома
Вакуоля
сферосома (мезоплазма)
(мезоплазма) Клітинна
стінка
Клітинна
стінкаРис. 2. Будова рослинної клітини
О
Органоїд
(від пізнолат. organizo –
засвоюю, упорядковую, від давньогр.
ὄργανον (organon) –
знаряддя і εἶδος (eidos) – вид)
ядро(не в усіх клітинах),
хромосоми (на період поділу клітини),
мітохондрії,
пластиди(тільки в клітинах рослин),
рибосоми,
ендоплазматичну сітку (ендоплазматичний ретикулум),
апарат (комплекс) Гольджі,
вакуолі,
мікротрубочки,
мікрофіламенти.
1
Мембрана
(від латинської membrana – пергамент)плівка,
напівпроникний роздільник середовищ,
оболонка
Глобулярний
(від лат. globus –
куля,
круглий)
Сферичнний
(від гр. σφαῖρα (sphaira) – куля)
Будова. Мембрани складаються із подвійного шару молекул фосфоліпідів у який занурені глобулярні (сферичні, кулясті) молекули білків (рис.3.). Мембрани в клітині утворюють органели різної форми, вірніше оболонки (стінки) органел, а саме мембрани згортаються у пухирці різних форм і трубочки. Мембрани органел клітини і плазмалеми періодично змінюють свою площу і загальну форму самих органел.
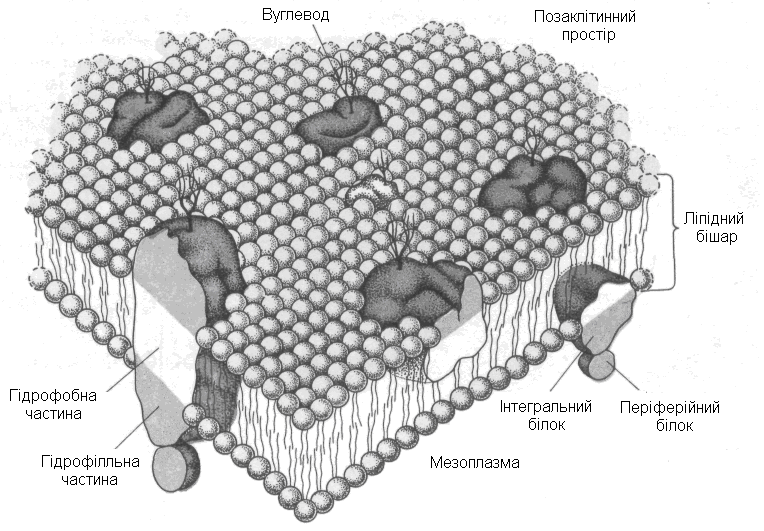
Рис. 3. Рідинно-мозаїчна модель мембрани
Основною властивістю живих біологічних мембран є вибіркова проникність, яка порушується хімічними (кислоти та ін.) і фізичними (температура та ін.) чинниками.
Функціїмембран в клітині визначаються білками, що входять до їхнього складу. Мембрани:
виконують бар’єрну функцію,
пропускають вибірково крізь себе за допомогою білкових каналів і ланцюгів молекули різних речовин, іони та електрони (транспортна функція),
отримують і передають сигнали з навколишнього середовища у протопласт (сигнальна функція).
Ендоплазматичний
ретикулум (від гр. (endon) –
усередині, πλάσμα (plasma) –
виліплене, оформлене і від лат. reticulum –
сітка)
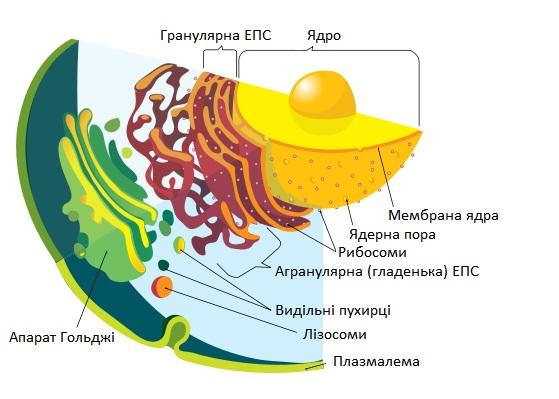
1.1.2. Ендоплазматична сітка (ЕПС) або ендоплазматичний ретикулум (ЕПР) – органела клітини у вигляді системи з’єднаних одномембранних трубочок (каналів), пухирців і плоских цистерн (порожнин). Діаметр трубочок ЕПС дорівнює 0,05 - 0,10 мкм (іноді до 0,30 мкм). Товщина мембранних стінок ЕПС складає біля 5 нм, 0,005 мкм. ЕПС не є стабільною структурою і часто змінює свою форму.
Розрізняють два типи ЕПС:
гранулярна або шорсткувата (на її поверхні містяться гранули рибосом),
агранулярна або гладенька (без рибосом).
Гранулярна ЕПС транспортує переважно білки, які синтезують рибосоми на її поверхні, агранулярна ЕПС – ліпіди та вуглеводи. Для тимчасового або довготривалого зберігання запасних поживних речовин на мембранній трубочці ЕПС видувається пухирець вакуолі у який завантажуються необхідні речовини. Після чого пухирець вакуолі від’єднується від трубочки ЕПС. Таким чином ЕПС формує не тільки вакуолі, але і пухирці апарату Гольджі – диктіосоми.
|
б а
Рис. 4. Взаємозв’язок мембранних органел клітини: а. Будова ендоплазматичної сітки, б. Мембрана ЕПС згорнута у трубку |
ФункціїЕПС:
синтез клітинних мембран;
утворення мембранних органел: диктіосом апарату Гольджі, вакуоль, оболонок ядра;
синтез і транспорт речовин по системі трубок ЕПС
Диктіосома
(від гр. (diktyon) – сітка і (soma) –
тіло)
Гольджі
(Goldgi) Каміло (1844-1926) італійський гістолог,
іноземний член-кореспондент Петербургської
академії наук (1905). Створив фундаментальні
праці з гістології та морфології
нервової системи. Розробив (1873) метод
приготування препаратів нервових
клітин. Установив два типи нервових
клітин. Описав так званий Гольджі
комплекс та
Нові
мембрани синтезуються на гранулярному
ендоплазматичному ретикулумі. Дрібні
пухирці відбруньковуються від гладенької
поверхні ЕПС і переносять мембрани з
речовинами в середині до диктиосоми,
що формується. Секреторні пухирці
відокремлюються від цистерн диктиосом,
що дозрівають, мігрують до плазматичної
мембрани і зливаються з нею, додаючи
нову порцію мембран у плазмолему і
будівельних матеріалів у клітинну
оболонку.
1.1.3. Апарат (комплекс) Гольджі – органела клітини у вигляді набору (стопки) із 4-20 плоских пухирців – диктіосом, що постійно оновлюються. Дископодібні пухирці (цистерни) диктіосом з країв розгалужуються у складну систему трубочок. Нові диктіосоми апарату Гольджі утворюються в результаті злиття проміжних пухирців, які відокремлюються від ЕПС. У диктіосомах відбувається перетворення (дозрівання) речовин, що були виділені з ЕПС. Диктіосоми поступово переміщуються у бік плазмалеми, де крайня диктіосома розпадається на секреторні пухирці. Секреторні пухирці підходять до плазмалеми, стикуються з нею своєю мембранною оболонкою (у пухирці утворюється отвір назовні плазмалеми), після чого пухирець вивертається наізнанку. Усі речовини, що містились у секреторному пухирці, виштовхуються назовні плазмалеми протопласту, а мембрана пухирця вбудовується у плазмалему. Речовини, що виділяються протопластом клітини за допомогою секреторних пухирців апарату Гольджі, нашаровуються на клітинну стінку, яка оточує протопласт. Найбільша активність апарату Гольджі спостерігається під час формування нових клітинних стінок після поділу клітин, а також на етапах росту клітин, коли молоді клітинні стінки розтягуються під тиском протопласту, що росте і збільшується у розмірах.
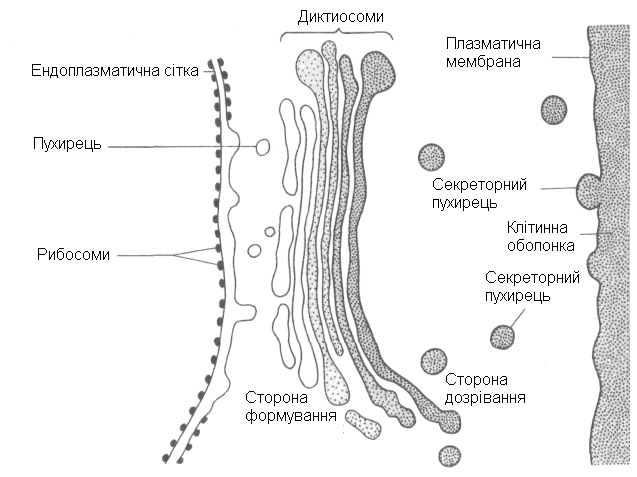
Рис. 5. Схематична ілюстрація ендомембранної концепції
(утворення і робота диктіосом апарату Гольджі)
Функціїапарату Гольджі:
секреторна або видільна, виводить за межи протопласту непотрібні в середині клітини речовини;
переміщує мембрани ЕПС до плазмалеми;
нарощує клітинну стінку;
створює, як і ЕПС, різного типу вакуолі.
1
Вакуоля
(від французької vacuole, від лат. vacuus –
пустий)
Тургор
(пізнолат. turgor – здуття, наповнення,
від лат. turgere – бути набряклим,
наповненим)
Плазмоліз
(від гр. πλάσμα – виліплене, оформлене
і λύσις - розкладання, розпад)
Градієнт
концентрації (від лат. gradi, gradu, gradus –
хід, рух, течія, наближення; con – з,
разом, сумісно + centrum – центр)
Гіпертонічний
(від гр. (hypér) – над, зверх і лат. tonus,
гр. (tоnos) – напруга)
Гіпотонічний
(від гр. ὑπο – під, внизу і лат. tonus,
гр. (tоnos) – напруга)
Ізотонічний
(від гр. ισος – рівний і лат. tonus, гр.
(tоnos) – напруга)
скорочувальні або пульсуючі,
лізосоми,
пероксисоми,
сферосоми.
1.1.4.1. Скорочувальні (пульсуючі) вакуолі – одномембранні пухирці утворені ендоплазматичною сіткою і апаратом Гольджі і заповнені водним розчином солей – клітинним соком. Молодим клітинам притаманна велика кількість маленьких вакуоль. Внаслідок поглинання води дрібні вакуолі меристематичних (с. 123) клітин збільшуються, зливаються і сприяють розтягненню молодої клітини. Часто пульсуючи вакуолі зливаються у одну велику вакуолю, яка притискає гіалоплазму з іншими органелами до клітинної стінки.
Скорочувальні вакуолі створюють в клітині осмотичний (тургорний) тиск і підтримують водно-сольовий баланс через явища плазмолізу і деплазмолізу.
Плазмоліз – процес виходу води із вакуолі клітини за градієнтом (перепадом) концентрації у зовнішній гіпертонічний розчин до встановлення ізотонічної рівноваги. Внаслідок плазмолізу:
клітина втрачає тургор,
із-за виходу води із вакуолі концентрація солей клітинного соку збільшується;
вакуоля скорочується – зменшується у розмірах і стягує за собою протопласт, що призводить до відокремлення плазмолеми протопласту від клітинної стінки (крім ділянок плазмодесм) і стискування протопласту у центрі або куті клітини.
Рис.
6. Плазмоліз у епідермальній клітині.
А.
При нормальних умовах протопласт
заповнює простір, обмежений клітинною
стінкою. Б. Якщо клітину розмістити у
концентрованому розчині сахарози, вода
починає виходити із клітини у гіпертонічну
середу, плазматична мембрана скорочується.
Перенесена у більш концентрований
розчин сахарози, клітина втрачає ще
більше води, и її протопласт стискається
сильніше.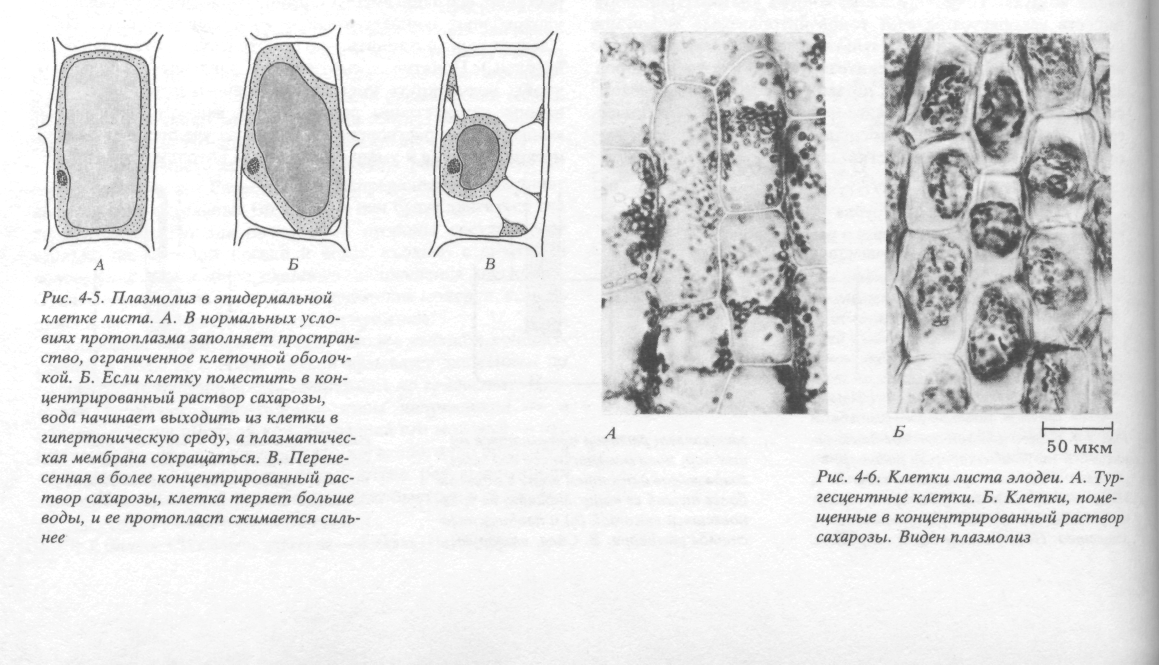
Деплазмоліз – зворотній плазмолізу процес з відновленням тургору – поверненням води у скорочувальну вакуоль клітини за градієнтом концентрації із гіпотонічного розчину до встановлення ізотонічної рівноваги.
1
Лізосома
(від гр. λύσις –
розчиняю, і σῶμα (sōma) – тіло)
Аутофагія
(від давньогр. αὐτός – сам і φαγεῖν –
«їсти»)
Авто́лиз,
ауто́лиз, (від давньогр. αὐτός – сам
і λύσις – розкладення, розпад)
Пероксисома
(від лат. per – посилення, надлишок
чого-небудь; гр.
(oxys) – кислий, гр. (soma) – тіло) –
органела в якій відбувається найвищий
ступінь окислення елементів
Сферосома
(від гр. (sphaira) – куля і σῶμα (soma) –
тіло)
Мітохондрія
(від гр. μίτος (mitos) –
нитка та χόνδρος (chondros) –
зернинка, крупинка)
Кристи
(від лат. crista – гребінь)
Матрикс
(від лат. mater – мати, основа)
перетравлення захоплених клітиною чужорідних тіл;
аутофагія– знешкодження непотрібних клітині структур, наприклад підчас заміни старих органоїдів новими або перетравленні білків та інших речовин утворених всередині самої клітини;
автолізу– програмованого самоперетравлення клітини, що призводить до її загибелі (процес супроводжує розвиток організму або диференціювання деяких спеціалізованих клітин).
1.1.4.3. Пероксисоми (мікротільця) – вакуолі розміром від 0,2 до 1,5 мкм, що містять велику кількість ферментів, які каталізують (прискорюють) окисно-відновні реакції (оксидази D-амінокислот, уратоксидази і каталази). Набор функцій пероксисом відрізняється в клітинах різних типів. Серед них: окислення жирних кислот, фотодихання, руйнування токсичних сполук та ін. Наряду с мітохондріями пероксисоми є головними споживачами кисню (O2) в клітині.
1.1.4.4. Сферосоми – дрібні сферичні вакуолі (пухирці) утворені гладкою (агранулярною ЕПС), які виконують функцію синтезу і накопичення ліпідів.
1.1.5. Мітохондрії – напівавтономні двомембранні органели клітини. Мають форму видовжених цистерн довжиною 1-2 мкм (до 7 мкм) і діаметром 0,3-1,0 мкм.
Будова. Мітохондрії складаються із двох мембран: зовнішньої та внутрішньої. Внутрішня мембрана більша за площею, ніж зовнішня, тому має складчастий вигляд. Складки внутрішньої мембрани мітохондрій називаються кристами. У середині мітохондрії заповнені дрібнозернистою напіврідкою речовиною – матриксом. У матриксі мітохондрій знаходяться метаболічні ферменти, мітохондріальні РНК і ДНК, рибосоми. Нові мітохондрії утворюються шляхом поділу материнської мітохондрії перетяжкою навпіл (амітозом).
Функції мітохондрій:
аеробне дихання – поступове розщеплення вуглеводів до вуглекислого газу (СО2), з утворенням води (Н2О) внаслідок приєднання до кисню (О2) протонів (Н+) та електронів (е-) водню. У процесі дихання енергія розщеплення вуглеводів забезпечує відновлення (зарядження) молекул-енергоносіїв, наприклад: нуклеотид АДФ (аденозиндифосфат) відновлюється до АТФ (аденозинтрифосфата) шляхом приєднання залишку однієї фосфорної кислоти. Молекули-енергоносії приймають участь у окисно-відновних реакціях, що проходять у клітині, де віддають енергетичний заряд (розщеплюються) для синтезу різних сполук, активації осмосу та інших процесів. Після цього повертаються для відновлення у мітохондрії.
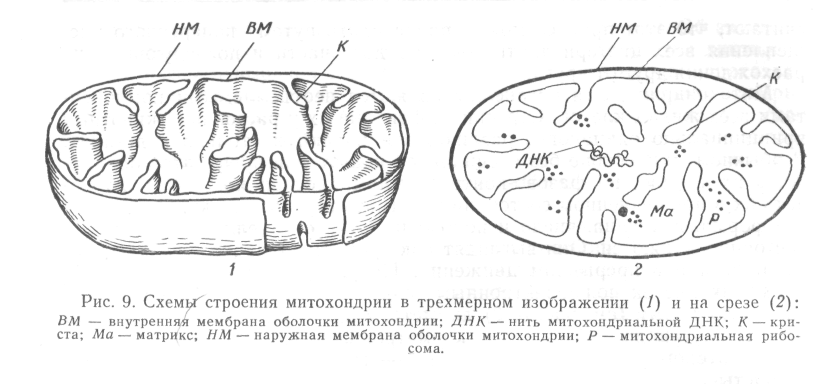
Рис. 7. Схеми будови мітохондрії:
1 – тривимірне зображення, 2 – поперечний зріз, ВМ – внутрішня мембрана оболонки мітохондрії; ДНК – нитка мітохондріальної ДНК; К – криста; Ма – матрикс; НМ – зовнішня мембрана оболонки мітохондрії; Р – мітохондріальна рибосома
Пластид
(від гр. πλαστός (plastos) –
виліплений,
утворений)
Строма
(від гр. στρῶμα (stroma) –
підстилка),
у біологічному значенні – основна
опорна структура органів і органоїдів
(органел)
Пропластиди
(від гр. (pro) –
до, перед
і
πλαστός (plastos) –
виліплений,
утворений)
1.1.6. Пластиди – напівавтономні двомембранні органели клітини переважно дископодібної форми.
Будова. Пластиди мають зовнішню гладеньку мембрану і внутрішню – різного ступеню розвиненості. У середині пластиди заповнений колоїдним розчином – стромою. У стромі знаходяться: пластидні РНК і ДНК, рибосоми, відповідні метаболічні ферменти, енергетичні молекули (АТФ, НАДФ·Н та ін.), а також продукти метаболізму вуглеводи, ліпіди і білки. Залежно від ступеню розвиненості внутрішньої мембрани пластидів і функцій, які вони виконують, пластиди поділяють на чотири типи:
пропластиди,
хлоропласти,
лейкопласти,
хромопласти.
1
Етіопласт
(від гр. (aitia) – причина і πλαστός
(plastos) –
виліплений, утворений)
1
Хлоропласт
(від гр. χλωρός (chloros) –
зелений і πλαστός
(plastos) –
виліплений, утворений)
Тілакоїд
(від гр. (thylakos) – мішок)
Ламела
(lamella) – підстилка
Грана
(від лат. granum – зернинка
Пігменти
(від лат. pigmentum – фарба) –
складні молекули які сприймають світло,
енергія якого використовується у
світлових реакціях фотосинтезу.
Хлорофіл
(від гр. χλωρός (chloros) –
зелений і φύλλον (phillon) –
листок)
Каротин
(від лат. carota – морква)
Ксантофіл
(від гр. (xanthós) – жовтий і φύλλον
(phillon) –
листок)
Мезофіл
(від гр. μέσος (mesos) –
середній, проміжний і φύλλον (phillon) –
листок), середня м’ясиста частина
листка
Особливості будови. Внутрішня мембрана хлоропластів добре розвинена, має видуті плоскі пухирці – тилакоїди і ламели, які представляють собою єдину взаємозв’язану систему. Тилакоїди розташовуються один над одним формуючи так звані стопки – грани. У одному хлоропласті може бути декілька гран, які з’єднані між собою довгими тилакоїдами строми - ламелами. У мембранах тилакоїдів розміщені фотосинтезуючі пігменти: хлорофіл (зелений), каротин (жовтогарячий), ксантофіл (темно-червоний) та ін. Зелений колір хлоропластів визначає пігмент хлорофіл який міститься у великій кількості і заглушає кольори інших пігментів.
Функція хлоропластів:
утворення вуглеводів (глюкози) у процесі фотосинтезу із молекул вуглекислого газу (СО2) і атомів водню (Н) води (Н2О) під дією світлової енергії сонця:
6СО2 + 12Н2О ―енергія світла→ 6О2↑ + С6Н12О6 + 6Н2О
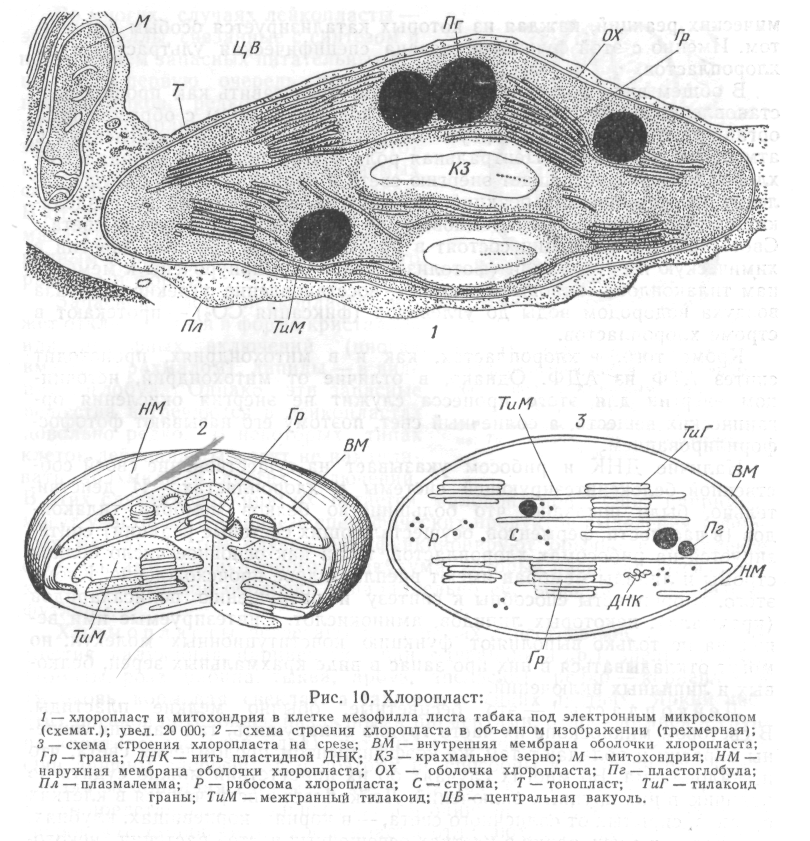
Рис.8. Хлоропласт:
1 – хлоропласт в клітині під електроним мікроскопом; 2, 3 – схеми будови хлоропласту; ВМ – внутрішня мембрана оболонки хлоропласту; Гр. – грана; ДНК – нитка пластидної ДНК; КЗ – крохмальне зерно; М – мітохондрія; НМ – зовнішня мембрана оболонки хлоропласту; ОХ – оболонка хлоропласту; Пг – пластоглобула; Пл – плазмалема; Р – рибосома хлоропласта; С – строма; Т – тонопласт; ТиГ – тилакоїд грани; ТиМ – міжгранний тилакоїд; ЦВ – центральна вакуоль
Хлоропласти знаходяться переважно у клітинах листків рослин і однорічних зелених пагонів на які попадає сонячне світло. У одній клітині мезофілу може знаходитись до 40-50 хлоропластів. Без отримання світла у хлоропластах поступово зникають пігменти.
1
Лейкопласт
(від гр. λευκός (leukos) –
білий і πλαστός (plastos) –
виліплений,
утворений)
Амілопласт
(від гр. (ámylon) – крохмаль і πλαστός
(plastos) –
виліплений, утворений)
Протеїнопласти
(від новолат. protein –
простий білок і гр. πλαστός (plastos) –
виліплений, утворений)
Елайопласти
(від гр. (elaion) – оливкове масло (олія)
і πλαστός (plastos) –
виліплений, утворений)
Ліпід
(від гр. λίπος (lípos) – жир)
Особливості будови. Лейкопласти мають слабо розвинену внутрішню мембранну систему. На пухирцях і виступах внутрішньої мембрани відсутні пігменти, тому лейкопласти виглядають безбарвними (рис 9).
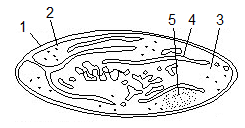
Рис. 9. Будова лейкопласту:
1 – зовнішня мембрана, 2 – внутрішня мембрана,
3 – строма; 4 – ламела; 5 – крохмальне зерно
Функції лейкопластів – накопичення запасних поживних речовин (включень): вуглеводів (крохмалю), ліпідів і білків.
Залежно від типу запасних поживних речовин які накопичують лейкопласти їх поділяють на:
амілопласти(накопичують крохмаль – запасній вуглевод у вигляді крохмальних зерен),
протеїнопласти (накопичують запасний білок у вигляді алейронових зерен),
елайопласти (накопичують запасні ліпіди у вигляді ліпідних крапель).
Лейкопласти розташовуються в клітинах органів рослини, які скриті від потрапляння прямих променів сонця (середині стебел, підземних органах: бульбах, коренеплодах, кореневищах, цибулинах, коренях. Під дією світла лейкопласти перетворюються на хлоропласти.
1
Хромопласт
(від гр. χρῶμα (chroma) –
колір, фарба і πλαστός
(plastos) –
виліплений, утворений)
Особливості будови. Внутрішня мембрана хромопластів не має майже ні яких виростів. Пігмент хлорофіл у хромопластах відсутній, а пігменти каротиноїди (каротин, ксантофіл та ін.) знаходяться у стромі цих пластидів, де збираються у формах:
глобул (кульок) – глобулярні хромопласти,
фібрил (волокон, ниток) – фібрилярні хромопласти,
кристалів – кристалічні хромопласти.
Рис.
10. Будова хромопласту:
1 –
подвійна
мембрана,
2 –
строма,
3 –
фібрили пігментів;
4 –
ліпідні краплі
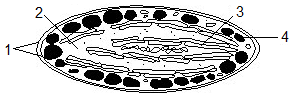
Функціяхромопластів, накопичення пігментів каротиноїдів.
Хромопласти знаходяться у клітинах пелюсток квітів, плодів, а також утворюються із хлоропластів у клітинах листків восени. Цей процес призводить до зміни кольору листка з зеленого на жовтий або червоний.
1.1.7. Рибосоми – безмембранні органели, діаметром 20 нм.
Будова. Рибосоми складаються із молекули білка і молекули р-РНК (рибосомальної рибонуклеїнової кислоти).
Рибосома
(від лат. ribes – струм і гр. σῶμα
(sōma) – тіло)
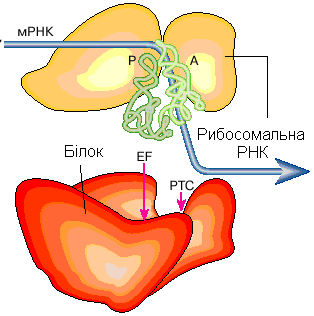
Рис. 9. Будова рибосоми:
а – загальний вигляд;
б – шлях проходження матричної РНК через рибосому під час синтезу білка
Функція рибосом – синтез молекул білка у процесі трансляції із молекул амінокислот. Рибосоми розміщуються у ядрі клітини, мітохондріях, на поверхні гранулярної ЕПС, вільно у гіалоплазмі. Рибосоми в клітині можуть розташовуватись поодиноко, тоді їх називають моносомами, або ланцюгами по 4-40 шт. – полісомами.
Мікрофіламент
(від гр. μικρός (micros) –
малий і пізнолат. filamentum – ниткоподібне
утворення, нитка)
1.1.8. Мікрофіламенти – безмембранні органели клітини у вигляді білкових тяжів.
Будова. Мікрофіламенти утворені сферичним білкомактином, який здатен до скорочення (рис. 10). Актинові нитки білку закручені спірально, надаючи мікрофіламентам форму тонких кручених (витих) канатів. У гіалоплазмі вони створюють загальну для клітини мережу – мікротрабекулярну решітку – мобільну (рухому) арматурну (опорною) конструкцію для інших органел клітини, яка постійно змінює свою форму (рис. 11).
2
1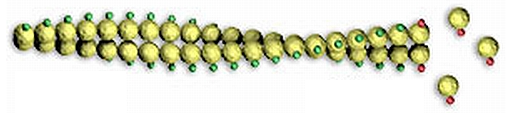

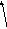
Рис. 10. Структура мікрофіламенту: 1 – F-актин (полімер), 2 – G- актин (мономер)
Рис.
11. Цитоскелет клітини:
1 –
мікротрабекулярна
решітка; 2 – мікротрубочка;
3 –
ендоплазматичний ретикулум;
4 –
клітинна мембрана;
5 –
мітохондрія;
6 –
рибосоми (полісоми);
7 –
окремі мікрофіламенти.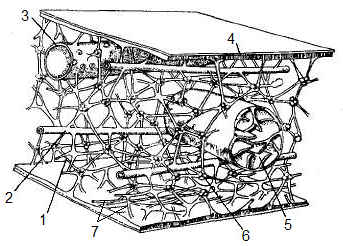
Функціїмікрофіламентів:
створюють внутрішньоклітинну опору протопласту, та органелам у ньому, у вигляді мікротрабекулярної решітки,
за рахунок скорочення актинових ниток мікрофіламентів, відносно дна одної, генерується (утворюється) тік (рух) гіалоплазми протопласту через мікротрабекулярну решітку.
1
Мікротрубочка
(від гр. μικρός (micros) –
малий і трубочка)
Будова. Мікротрубочки утворені сферичним білком тубуліном, який спірально закручується у форму трубочки (рис. 12). Органела виконує для інших органел функцію рейок, для переміщення їх по клітині. Мікротрубочки утворюються (самоскладаються із молекул білка) на тих ділянках протопласту, де необхідно, за умов невикористання вони знов розбираються на окремі молекули.
Рис.
12. Структура
мікротрубочки:
1 –
α-Тубулин,
2 –
β-Тубулин,
3 –
гетеродімер,
4 -
протофіламент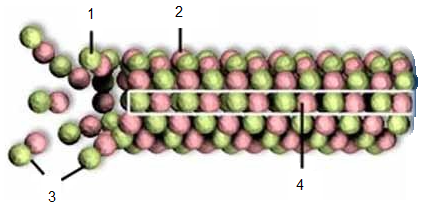
Функціїмікротрубочок:
проведення (транспорт) органел по гіалоплазмі,
утворення опорної структури – веретена поділу під час поділу клітини мітозом або мейозом для переміщення хромосом,
утворення фрагмопласту – системи спрямовуючих рейок, для переміщення секреторних пухирців апарату Гольжі до центру клітини, де під час цитокінезу (розділення материнської клітини навпіл) формується нова клітинна стінка,
орієнтування відкладання целюлозних мікрофібріл на клітинній стінці,
разом з мікрофіламентами створює внутрішній цитоскелет (опорну арматуру) протопласту, який підтримує його форму,
входить у склад джгутиків і війок клітин, забезпечує їхній рух, а також є складовим компонентом центріолей.
1
Цетнріоль
(від лат. centrum, гр. κέντρον (kentron) –
середина точка, центр, вістря, осереддя)
Будова. Центріолі утворені дев’ятьома групами триплетів мікротрубочок (по три мікротрубочки у кожній групі), що розташовуються по колу формуючи полий циліндр (рис. 13). Центріолі зазвичай розташовуються біля ядра, оточені зоною більш прозорої гіалоплазми, від якої відходять тонкі фібрили (мікротрубочки) – центросфера. Сукупність центріолей і центросфери називають клітинним центром. Зазвичай протягом клітинного циклу центріоль подвоюється один раз. Поряд з кожною «материнською» центріоллю добудовується дочірній циліндрик; відбувається це, як правило, протягом S-періоду інтерфази.
Цетнросома
(від лат. centrum, гр. κέντρον (kentron) –
середина точка, центр, вістря, осереддя
і σῶμα (sōma) – тіло)
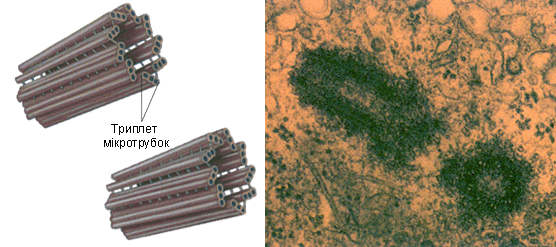
Рис. 13. Модель центріолі. Зображені дев’ять триплетів мікротрубочок
Функція центріолей –бути базою (початковою, опорною структурою) для формування мікротрубочок. Центріолі є центрами формування мікротрубочок джгутиків і війок клітин, а також веретена поділу у метафазі мітозу або мейозу в тваринних клітинах.
Евкаріот
(від гр. εύ (eu) – добре, повністю та
κάρυον (karyon) – ядро) – організми
які у складі своїх клітин мають повністю
сформоване ядро
Каріоплазма
(від гр. κάρυον (karyon) – ядро і πλάσμα
(plásma) – буквально виліплене, оформлене)
Хроматин
(від гр. χρῶμα (chroma) – колір, фарба і
(nitos) –
нитка)
Нуклеоль
(від лат. nucleus – ядро)
Генетичний,
ген (від гр. γενητως
(gentos,
genos) –
рід, походження)
ДНК –
дезоксирибонуклеї́нова кислота́ –
ядерна кислота з безкисневим цукром
рибоза (від араб. робб –фруктовий
сироп)
РНК –
рибонуклеї́нова кислота́ – ядерна
кислота з цукром (рибоза)
1.1.10. Ядро – головна органела евкаріотичної клітини.
Будова. Ядро має подвійну оболонку, яка формується ЕПС і з’єднана з нею. Середина ядра заповнена каріоплазмою – ядерним соком. У каріоплазмі знаходиться хроматин (хроматинові нитки), основу якого складають молекули ДНК, а також РНК і білок. Молекули РНК і білок (рибонуклеопротеиди) формують у ядрі ядерце (нуклеоль), яке є складовою частиною однієї з хроматинових ниток (рис. 14).

Рис. 14. Будова ядра
Функція ядра – бути командним центром клітини, задавати програму дій усім структурним компонентам клітини, зберігати генетичну (спадкову) інформацію про розвиток організму, у вигляді молекул ДНК, і передавати її у дочірні клітини під час поділу. Функцією ядерець є утворення молекул РНК для рибосом.
1
Нуклеопептид
(від лат. nucleus – ядро і гр. πεπτος
(peptos) –
поживний) – група речовин, молекули
яких побудовані із залишків α-амінокислот,
з’єднаних у ланцюг пептидними (амідними)
зв’язкам (–CO–NH–)
ДНК – дезоксирибонуклеїнова кислота представляє собою макромолекулу, мономерами (блоками) якої є нуклеотиди (рис. 15). Кожен нуклеотид складається із азотистої основи, цукру (дезоксирибози) і фосфатної групи (або гомологічної арсеноїдної). В молекулах ДНК нуклеотиди утворюють довгий ланцюг у якому з’єднані між собою фосфатна група одного нуклеотиду з дезоксирибоза іншого. ДНК має у своєму складі два спірально закручені нуклеотидні ланцюги з’єднуються азотистими (пуриновими і піримідиновими) основами. Так, піримідинова гетероциклічна основа (сполука) тимін з’єднується з пуриновою основою аденіном, а цитозін – з гуаніном. Послідовність розташування азотистих основ у молекулі ДНК і є кодом через який реалізується програма життя біологічних організмів.
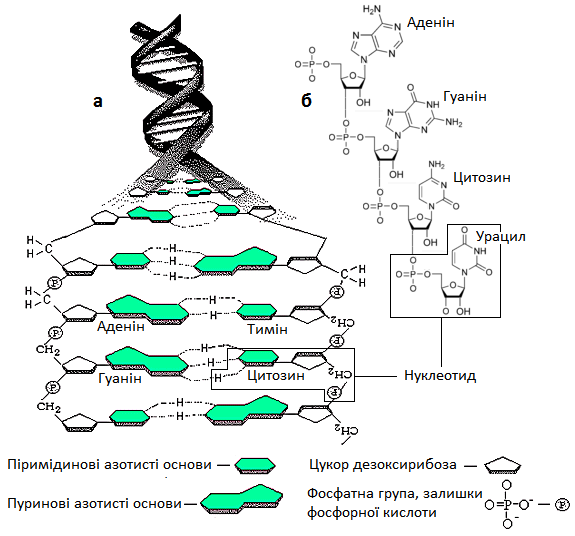
Рис. 15. Схеми нуклеїнових кислот: а – молекули ДНК, б – молекули РНК
РНК – рибонуклеїнова кислота є одноланцюговою молекулою, яка утворюється із нуклеотидів у процесі транскрипції (перезапису інформації) на певних ділянках ДНК. Тільки замість тиміну у склад РНК входить піримідинова основа – урацил.
Якщо молекули ДНК э сховищем інформації (креслень усіх елементів і процесів у клітинах організму), то РНК є трансляційними молекулами, за кодами яких на рибосома створюються молекули білків, з певною послідовністю розташування в них амінокислот. Послідовність розміщення в білках амінокислотних залишків визначає їх властивості і функції, які використовуються у житті організму.
Похідні протопласту
Ергастичний
(від гр. (ergastikos) – діяльний) –
вироблений
Похідні протопласту – ергастичні речовини – продукти життєдіяльності (метаболізму) живих компонентів клітини (протопласту). До похідних протопласту відносять:
фізіологічно активні речовини (ферменти, фітогормони, вітаміни та ін.);
продукти обміну речовин – видимі включення (крохмальні та алейронові зерна, ліпідні краплі, кристали та ін.) і розчинені у клітинному соку та гіалоплазмі;
клітинну стінку.
1.2.1. Фізіологічно активні речовини – органічні речовини різної хімічної природи, які стимулюють або пригнічують біохімічні процеси у клітині. Для них властива висока активність у дуже малих концентраціях (0,001 – 0,0001 мкг) і велика специфічність дії. До фізіологічно активних речовини належать:
ферменти і вітаміни – каталізатори хімічних реакцій,
фітогармони– ростові речовини,
фітонциди– захисні отруйні речовини.
1
Фермент
(від лат. fermentum –
закваска)
Ензим
(від гр. ἔνζυμον, ἔν (en) – в (у),
всередині і ζύμη (zyme) – закваска)
Ферменти
діють за принципом автоматичної зброї.
При контакті з молекулою субстрату
(патроном), зведений фермент розряджається,
передаючи свою енергію на субстрат,
тим самим зміщуючи електронні хмари
хімічних елементів цієї молекули і
надаючи їй змогу взаємодіяти з іншими
компонентами реакції. У ході реакції
частина енергії повертається до ферменту
перезаряджаючи його, тобто зводить.
Ферменти за своєю будовою поділяють на:
прості ферменти - складаються з молекул білку;
складні ферменти– складаються з молекули білку (апоферменту) і небілкової молекули (коферменту), які у каталітичних реакціях взаємодіють разом.
Ферменти приймають участь в усіх реакціях метаболізму в клітині, і забезпечують взаємодію усіх компонентів протопласту та його похідних.
1
Вітамін
(від лат. vita – життя та
minimum,
min – найменша,
маленька кількість)
Відомо 40 видів вітамінів які позначаються латиницею:
провітамін А (каротин) – жиророзчинний пігмент (забарвлена речовина), приймає участь у окисно-відновних процесах, фотореакціях, регуляції синтезу білків, обміні речовин, функціонуванні клітинних мембран, необхідний для росту нових клітин, уповільнює процес старіння;
в
Аскорбінова кислота (від гр. (a) – не, без і (scorbutus) – цинга) – кислота яка запобігає хворобі цингі
ітаміни групи В(В1, В2, В3 та ін.) – водорозчинні речовини; приймають участь у клітинному диханні і виробленні енергії;вітамін С (аскорбінова кислота) – водорозчинна органічна сполука, виконує біологічні функції відновника і коферменту деяких метаболічних процесів, є антиоксидантом;
вітаміни групи D – розчиняються в жирах, складаються із феролів, які активізуються під дією ультрафіолетових променів, регулюють засвоєння мінералів кальцію и фосфору;
вітаміни групи Е – жиророзчинні біологічно активні речовини (токофероли і токотриеноли), проявляють антиоксидантні властивості, є просторовими стабілізаторами фосфоліпідного шару біологічних (клітинних) мембран;
вітаміни групи К – жиророзчинні сполуки, похідні 2-метил-1,4-нафтохинона, використовується у синтезі деяких білків, засвоєнні кальцію і забезпеченні взаємодії кальцію з вітамінами групи D;
в
Фітогормони (від гр. φυτóν (phyton) – рослина і оρμόνη (hormáo) – приводжу до рух, збуджую)
Ауксин (від гр. αὔξω (áuxō) – збільшую, росту)
Фототропізм (від гр. φῶς (phos) – світло і τρόπος (trоpos) – поворот)
Геотропізм (від гр. γῆ (ge) – Земля і τρόπος (trоpos) – поворот)
Гіберелін (від лат. назви гриба Gibberella fujikuroi Sow., який вражає рослини рису і викликає їх надмірний ріст)
Цитокінін (від гр. κύτος – «вмістилище», тут: «клітина» і κινειν (kinéin) – рухатись)
Інгібітор (від (лат. inhibeo – стримую)
ітаміни групи Р –група флавоноїдів у яку входять: гесперидин, кумарини (ескулин), антоциани, катехіни та інші, приймають участь разом з вітаміном С у окисно-відновних реакціях.
1.2.1.3. Фітогормони (ростові речовини) – низькомолекулярні хімічні речовини, що утворюються в невеликих концентраціях в клітинах молодих пагонів і коренів, і регулюють швидкість їх росту і розвитку. До фітогормонів зазвичай відносять:
ауксини – хімічні сполуки на основі індолу (основний представник індолілоцтова кислота), які регулюють на різних етапах життя рослини її ріст, диференціювання тканин, ростові реакції на світло (фототропізм) і силу тяжіння (гравітації) (геотропізм);
гібереліни – сполуки дитерпенових поліциклічних кислот (основний представник гіберелова кислота), що стимулюють ріст і розвиток рослин;
цитокініни (кініни) – група гормонів рослин, похідні азотистої сполуки пурину (основний представник кінетин), які підвищують швидкість поділу клітин.
інгібітори– хімічні речовини, які пригнічують активність ферментів. До інгібіторів росту належить, наприклад, абсцизова кислота.
Фітогормони, на відміну від гормонів тварин, менш специфічні і утворюються переважно на місці безпосередньої дії (без транспортування).
Фітонцид
(від гр. φυτóν (phyton) – рослина і лат.
caedo –
вбиваю)
Антибіотик
(від
гр. ἀντί (anti) – проти і βίος (bios) –
життя)
1.2.1.4. Фітонциди (антибіотики) – різні за хімічним складом біологічно активні речовини, які вбивають або пригнічують ріст і розвиток мікроорганізмів (вірусних, бактеріальних і грибкових хвороб). Фітонциди розчинені у клітинному соку і цитоплазмі. Деякім з цих сполук притаманна летючість, що дає змогу діяти фітонцидам на мікроорганізми на відстані.
1.2.2. Клітинний сік – рідина яку виділяє протопласт (цитоплазма) у вакуолі. Водний розчин клітинного соку у своєму складі містить вуглеводи, таніни, алкалоїди, глікозиди, органічні кислоти і солі, пігменти, мінеральні солі.
1.2.2.1. Вуглеводи – група органічних сполук, первинних продуктів фотосинтезу. У клітинному соку рослин вуглеводи представлені:
моносахаридами(глюкоза, фруктоза) – містяться у вакуолях клітин плодів, наприклад винограду;
дисахаридами(сахароза) – накопичуються у коренеплодах цукрового буряку, стеблах цукрового очерету;
полісахаридами(інулін – ізомер крохмалю розчинний у воді) – знаходяться у бульбах жоржини, земляної груші (топінамбуру).
1.2.2.2. Таніни (дубильні речовини) – складні безазотисті сполуки – фенольні похідні, близькі до глікозидів. Вони мають в’яжучий смак і антисептичні властивості, які захищають рослини від пошкоджень мікроорганізмів. Таніни накопичуються в усіх клітинах, можуть просочувати клітинні стінки або міститись у окремих вмістилищах. Зазвичай клітини, які містять таніни, пов’язані з провідними пучками (с. 124).
1.2.2.3. Алкалоїди – високотоксичні азотовмісні сполуки лужної природи, які виконують у рослинах захисну функцію. Алкалоїди неоднорідні за складом і містяться у різних частинах рослин. У корі хінного дерева знаходиться хінін, у коренях і листках блекоти – атропін, у насінні і молочному соку маку – папаверин, морфін і кодеїн, у листках тютюну і махорки – нікотин.
1.2.2.4. Глікозиди – це похідні вуглеводів, які є сполученням цукрів із спиртами, альдегідами, фенолами та іншими безазотистими органічними речовинами. Глікозиди відкладаються у клітині, у якості як запасних, так і отруйних речовин. Під дією ферментів і при контакті з повітрям глікозиди легко розпадаються утворюючи ароматичні сполуки. Із отруйних глікозидів:
амигдалін (при гідролізі дає синильну кислоту), накопичується у насінні мигдалю, персика, абрикоса та інших кісточкових;
соланінзнаходиться у клітинному соку рослин родини пасльонові;
кумаринконцентрується у вегетативних і генеративних органах буркуна;
сапонін міститься у кореневищі мильнянки (Saponaria), у насінні куколю (Agrostemma) та інших гвоздичних, у розчинах піниться.
1.2.2.5. Органічні кислоти – яблучна (малат), лимонна, щавлева, винна, янтарна та інші – карбонові кислоти, органічні сполуки,що містять одну або декілька карбоксильних груп ( СООН).
Особливо багато органічних кислот у недозрілих плодах, де вони у процесі дозрівання перетворюються на ті або інші характерні для даного виду запасні або ароматичні сполуки. Органічні кислоти виконують частково роль фітонцидів (антибіотиків) і приймають участь у процесах фотосинтезу і дихання.
Кристал
(від гр. κρύσταλλος
(krystallos) –
лід)
Оксалат
(від
лат.
назви рослин роду кислиця –
Oxalis)
Рафід
(від гр. (rhaphis) –
голка)
Друза
(від
нім.
Druse –
щітка)
Ідіобласт
(від гр. (idios) – особливий і (blastos) -
росток)
1.2.2.6. Кристали – твердий фізичний стан сполук, атоми яких утворюють певну упорядковану періодичну (симетричну) структуру – кристалічну решітку, якій відповідає зовнішня симетрія форми. Солі багатьох органічних і неорганічних кислот знаходяться у клітинному соку у вигляді кристалів. У клітинах звичайні солі щавлевої кислоти або оксалати, наприклад оксалату кальцію.
Розрізняють три типи кристалів оксалату кальцію:
прості – монокристали (в лусках цибулі),
голчасті – рафіди (у традесканції, портукалу та ін.),
складні – друзи (у бегонії, інжиру та ін.).
Часто клітини, що містять великі кристали, деформуються, а клітини з рафідами, з досягненням ними граничного розміру відмирають, але зберігають у собі особливий слиз, здатний до набухання.
Кристали частіше зустрічаються у клітинах епідермісу (с. 125) і корової паренхіми (с. 126) трав’янистих і деревних рослин, а також старих секреторних паренхімних клітинах – ідіобластах. Найбільша кількість кристалів відкладається у листках перед листопадом. Частіше вони зустрічаються у рослин посушливих місць мешкання – ксерофітів, солончаків і т. д. Кристали, будучи солями, відіграють важливу роль у підтримці осмотичного тиску та кислотно-лужної рівноваги у клітині.
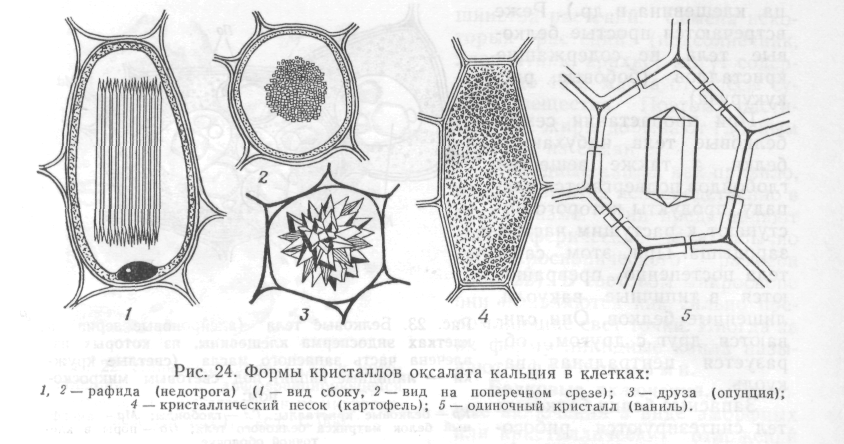
Рис. 16. Форми кристалів оксалата кальція в клітинах:
1,2 рафіди (недотрога) (1 – вид збоку; 2 – вид на поперечному розрізі); 3 – друза (опунція); 4 – кристалічний пісок (картопля); 5 – поодинокі кристали (цибуля)
1.2.2.7. Пігменти – забарвлені молекули різної хімічної природи. Для вакуоль найбільш характерні пігменти:
антоціан– глікозид за хімічною природою, яскраво-червоного, рожевого, фіолетового, теракотового, малинового, бордового кольору, який забарвлює пелюстки квітів, плоди та інші частини рослин;
антохлор – флавон за хімічною природою, жовтого кольору, який забарвлює пелюстки жовтої жоржини, льнянки, плоди цитрусових;
антофеїн– забарвлює клітинний сік у темно-бурий колір (плями на пелюстках кінських бобів).
Пігментація має велике біологічне значення для запилення квітів комахами і для розповсюдження плодів птахами та іншими тваринами.
1
Ергастичний
(від гр. (ergastikos) – діяльний)
нерозчинні– крохмальні і білкові зерна, геміцелюлози, жирні олії;
розчинні– прості вуглеводи (глюкоза, фруктоза, сахароза), деякі прості білки (альбуміни і глобуліни), вітаміни і т. д.
Розчинні речовини запасу були розглянуті у розрізі складових частин клітинного соку, тому у цьому розділі будуть розглянуті лише нерозчинні речовини запасу, крім геміцелюлоз.
1
Крохмаль
(від нім. Kraftmehl –
сильне, міцне борошно)
Асиміляційний
(від лат. assimilare –
робити подібним)
Гідроліз
(від гр. ὕδωρ (hydor) – вода і λύσις
(lysis) – розкладення, розчинення) –
розщеплення складних молекул на прості
за допомогою води
Транзитний
(від лат. transitus –
переправа)
первинний (асиміляційний) – утворюється у хлоропластах із глюкози при її надлишку, пізніше під дією ферментів гідролізуються до сахарози, яка транспортується до місць тривалого зберігання (підземних органів рослин);
транзитний– тимчасово синтезується із сахарози у пластидах клітин, через які іде транспорт вуглеводів до органів запасу, після чого знов під дією ферментів переводиться у сахарозу, яка транспортується далі;
вторинний (запасний) – полімеризується із сахарози в амілопластах органів запасу у вигляді крохмальних зерен.
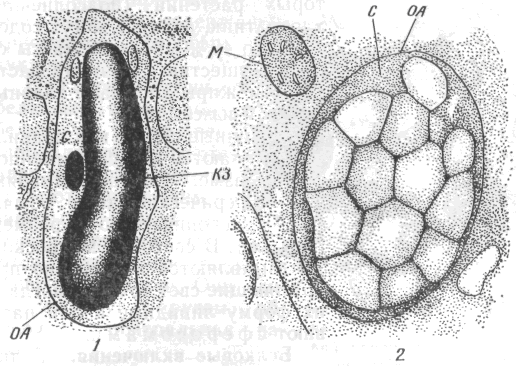
У амілопластах запасний крохмаль відкладається у стромі навколо утворювального центру у вигляді концентричних нашарувань. Форма і розміри крохмальних зерен характерні для кожного виду рослин. За своєю будовою крохмальні зерна поділяють на:
прості– з одним утворювальним центром, навколо якого розташовані шари крохмалю (пшениця, кукурудза, квасоля та ін.);
складні – мають багато утворювальних центрів із своїми шарами крохмалю (овес, гречка, рис, шпинат та ін.);
напівскладні– мають декілька (2-3 шт.) утворювальних центрів, які об’єднані загальними периферійними шарами крохмалю (картопля).
|
|
3 4 |
|
Рис. 17. Будова включень запастних поживних речовин: 1 – просте крохмальне зерно; 2 – складне крохмальне зерно; 3 – ліпідні краплі; 4 – алейронове зерно; Гл – глобулярний білок; КЗ – крохмальне зерно; Кр – білкові кристали; ЛК – ліпідна крапля; М – мітохондрія; Ма –аморфний білок матрикса білкового тіла; ОА – оболонка амілопласта; По – пора оболонки клітини. | |
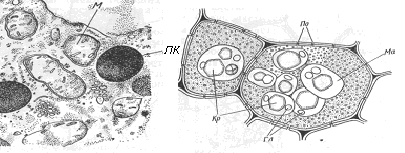
1
Конституційний
( від лат. constituere –
встановлювати; constitutio –
устрій)
конституційні– після утворення вбудовуються у органели клітин, приймають активну участь у метаболізмі;
запасні (ергастичні) – призначені для довготривалого зберігання у неактивному стані, виконують функцію енергетичного і будівельно-матеріального резерву в клітинах із зниженою активністю метаболізму.
Запасний білок відкладається у вакуолі клітин у аморфному стані, формуючи так звані алейронові зерна. Переважно білки запасаються у насінні рослин (наприклад, клейковина пшениці). Під час дозрівання насіння, яке супроводжується зневодненням, білкові вакуолі висихають і частина білка викристалізовується (перетворюється на кристалоїд алейронового зерна). У процесі зневоднення білкової вакуолі калієво-кальцієво-магнієва сіль інозитгексафосфорної кислоти виділяється на алейроновому зерні у вигляді кульки (глобули) – глобоїда. На відміну від звичайних білкових (протеїнових) зерен кристалоїди і глобоїди характерні тільки для алейронових зерен. Кристалоїди на відміну від справжніх кристалів (солей різних кислот) складаються з білків, які здатні до набухання у воді. Завдяки цій здатності алейронові зерна під час проростання насіння обводнюються, підвергаються розпаду, продукти якого поступають до зародку і перетворюються у скорочувальні вакуолі.
1.2.3.3. Жири – група органічних сполук молекули яких синтезуються з вуглеводів і є складними ефірами жирних кислот і гліцерину; входять до класу ліпідів. Жири відкладаються про запас у цитоплазмі усіх клітин та стромі пластидів у вигляді рідинних крапель. Великі запаси жиру накопичуються у насінні (соняшник, кунжут, бавовник, рицина, льон, конопля та ін.), а також у плодах (маслина) і паренхімі дерев.
Жири – найбільш економна форма запасних поживних речовин. При окисленні жирів на одиницю маси (1 г) виділяється значно більше енергії (38 кДж), ніж при окисленні вуглеводів і білків (18 кДж). Вони розчинні у органічних розчинниках (ефірі, хлороформі, толуолі, бензині, ксилолі та ін. ), слабо розчинні у спирті і нерозчинні у воді.
Плазмодесма
(від гр. πλάσμα (plasma) –
виліплене, оформлене і (desmos) – зв’язок,
зв’язка)
1.2.4. Клітинна стінка – це міжклітинна речовина (структура), продукт життєдіяльності протопласту рослинної клітини. Її будують органели – мікротрубочки і апарат Гольджі, розділяючи материнську клітину на дві дочірні. При утворенні нової клітинної стінки між протопластами двох дочірніх клітин залишається зв’язок у вигляді цитоплазматичних каналів – плазмодесм (діаметром 30-60 нм.) У середині деяких плазмодесм проходять трубочки ЕПС – десмотрубочки, які зв’язують ендоплазматичні сітки двох сусідніх клітин.
1.2.4.1. Будова клітинної стінки. Клітинна стінка складається із трьох шарів:
с
Пектин, пектинові речовини (від гр. πηκτός (pektos) – той що згорнувся, замерз)
Целюлоза (від лат. cellula, буквально – кімната, тут – клітина, кімната для протопласту) (клітковина) – полісахарид утворений залишками глюкози
ерединної пластинки,первинної клітинної оболонки,
вторинної клітинної оболонки.
1.2.4.1.1. Серединна пластинка представляє собою шар клітинної стінки який є загальним для двох сусідніх клітин. Серединна пластинка складається із пектинових речовин – полісахаридів утворених головним чином залишками галактуронової кислоти. Завдяки пектиновим речовинам серединна пластинка є еластичною і здатна до розтягування (рис. 17).
1.2.4.1.2. Первинна клітинна оболонка відкладається клітиною поверх серединної пластинки. Основу первинної клітинної оболонки складає целюлоза, яка відкладається у вигляді структурованої решітки (каркасу). Целюлозний каркас просочується:
г
Геміцелюлоза (від гр. ἦμι- (hemi-) – пів- і лат. cellula, буквально – кімната, тут – клітина) – напівцелюлоза, майже целюлоза
Глікопротеїд (від гр. γλυκός (glykys) – солодкий, πρῶτος (protos) – перший і εἶδος (eidos) – вид) – складний білок з вуглеводними компонентами
Гетероолігосахарид (від гр. ἕτερος (heteros) – інший, різний, ὀλίγος (oligos) – небагатий, незначний, (sakchar) – цукор і εἶδος (eidos) – вид)
еміцелюлозами – полісахаридами, мономерами яких є вуглеводи: галактоза, манноза, арабіноза, ксилоза та ін.;глікопротеїнами (глікопротеїдами) – складні білки, в яких білкова (пептидна) частина молекули ковалентно зєднана з однією або декількома групами гетероолігосахаридів;
пектиновими речовинами.
Глікопротеїди та пектинові речовини надають первинній оболонці еластичності та пружності, що необхідно при збільшенні (рості) клітини. Первинна клітинна оболонка відкладається навколо плазмодесм (цитоплазматичних каналів) більш тонким шаром, ніж на інших ділянках клітинної стінки, утворюючи так звані первинні порові поля. Елемент конструкції клітинної стінки із первинних порових полів сусідніх клітин і серединної пластинки між ними називається поровою мембраною.
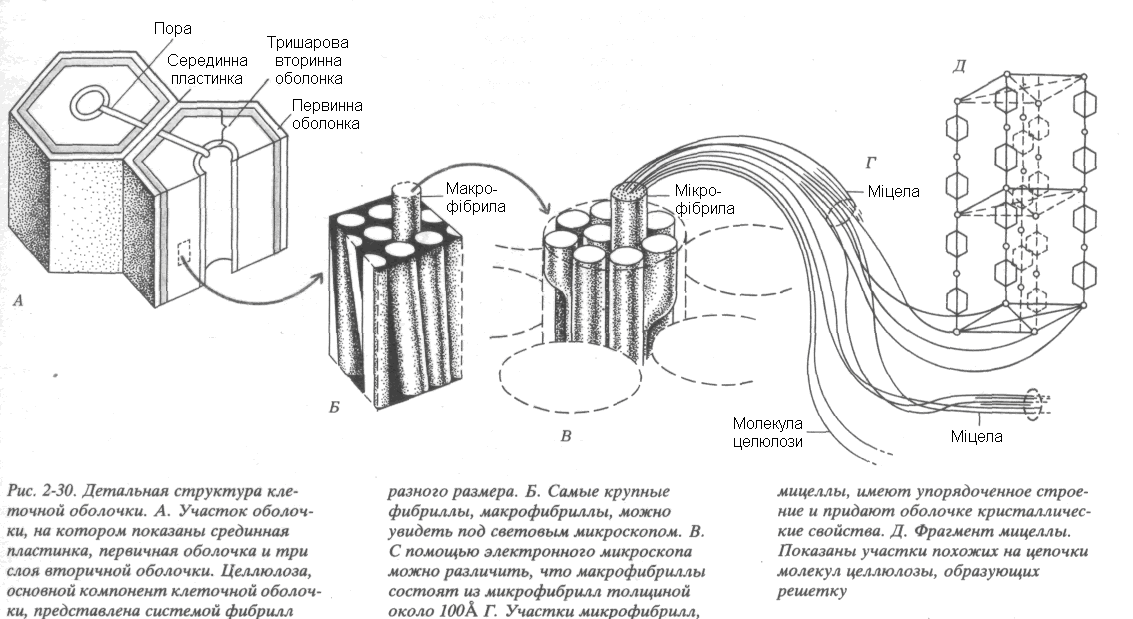
Рис. 18. Структура клітинної стінки:
А. Ділянка клітинної стінки, на якій вказані серединна пластинка, первинна оболонка і три шари вторинної оболонки. Целюлоза, основний компонент клітинної стінки, представлена системою фібрил різного розміру. Б. Найкрупніші фібрили, можна розгледіти під світловим мікроскопом. В. За допомогою електронного мікроскопу можна розрізнити, що макрофібрили складаються із мікрофібрил товщиною біля 100Å. Г. Ділянки мікрофібрил міцели, мають впорядковану будову и надають стінці кристалічні властивості. Д. Фрагмент міцели. Вказані ділянки схожих на ланцюги молекул целюлози, що утворюють решітку
|
|
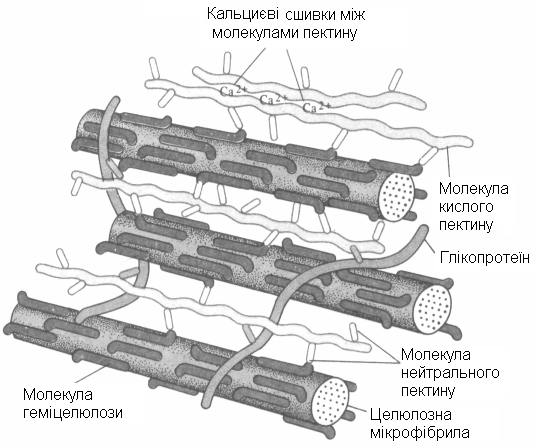
Рис.19. Схема можливого з’єднання мікрофібрил і компонентів матрикса (пектини, геміцелюлози і глікопротеїни) у первинній клітинній оболонці.
Молекули геміцелюлоиы прикріплені до поверхні целюлозних мікрофібрил водневими зв’язками. Деякі молекули геміцелюлози зв’язані з молекулами кислого пектину через молекули нейтрального пектину. Глікопротеїни, імовірно, приєднані до молекул пектину
|
1.2.4.1.3. Вторинна клітинна оболонка нашаровується поверх первинної клітинної оболонки. Вона складається з целюлози просоченої тільки геміцелюлозами. Вторинні оболонки характерні для механічних і водопровідних клітин, які завершили свій ріст і їхній протопласт поступово відмирає. У вторинної оболонки виділяють три шари – зовнішній, середній і внутрішній. Ці шари відрізняються один від одного за орієнтацією целюлозних мікрофібріл (волокон, що складають целюлозний каркас). Вторинна оболонка не відкладається на первинних порових полях, утворюючи в цих місцях пори (отвори). У вторинній клітинній оболонці розрізняють два типи пор:
прості(мають рівний край),
облямовані(мають нависаючий край над поровим полем).
|
|
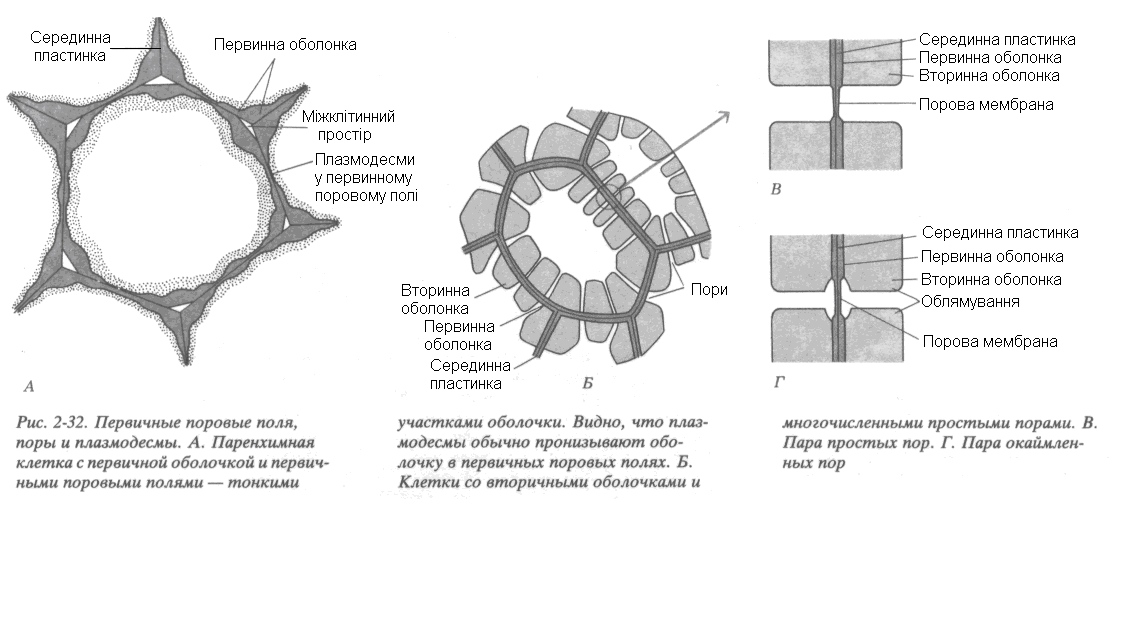
|
Рис. 20. Первинні порові поля, пори і плазмодесми: А. Паренхімна клітина з первинною оболонкою і первинними поровими полями – тонкими ділянками оболонки. Видно, що плазмодесми зазвичай пронизують оболонку у первинних порових полях. Б. Клітини з вторинними оболонками і чисельними простими порами. В. Пара простих пор. Г. Пара облямованих пор |
1.2.4.2. Перетворення (видозміни) клітинної стінки – це процеси пов’язані з відкладанням у матриксі клітинних оболонок крім пектинових речовин, геміцелюлоз і глікопротеїдів інших речовин, які надають клітинній стінці якісно нові властивості.
Лігнін
(від лат. lignum – дерево, деревина)
1.2.4.2.1. Здерев’яніння – процес просочення лігніном клітинної стінки (відкладання в матриксі клітинної стінки лігніну). Процес здерев’яніння починається з серединної пластинки, потім розповсюджується на первинну, пізніше вторинну оболонки. Лігнін – це полісахарид, який надає міцності клітинним стінкам. Здерев’яніння характерно для водопровідних клітин (судин або трахей) деревини (ксилеми).
1
Кутикула
(від лат. cuticula – шкірка)
1.2.4.2.3. Кутинізація – утворення на зовнішній поверхні стінок клітин епідермісу кутикули (восковий наліт на молодих пагонах і плодах). Кутикула представляє собою шар ліпідного полімеру кутину, у який занурений віск. Кутикула захищає органи рослин від пересихання, механічних пошкоджень, поглинає ультрафіолетові промені.
1.2.4.2.4. Мінералізація– відкладення у клітинних стінках й на їх поверхнях мінеральних речовин (кремнезему, вуглекислого кальцію та ін. у вигляді кристалів). Мінералізація характерна для клітин епідермісу (наприклад: клітини епідерми кропиви жалкої формують скловидні голкоподібні ампули заповнені мурашиною кислотою, які виконують захисну функцію).
Камедь,
гумі (лат. gummi – камедь) – клейка,
в’язка речовина
1.2.4.2.5. Ослизнення – утворення в клітинних стінках на основі пектинових речовин або целюлози сильно гідратованих полісахаридів – слизу й камеді. Слиз – суміш глікопротеїдів, утворює в’язкі водні розчини. Камедь (гумі) – високомолекулярний вуглевод близький до пектинових речовин, представляє собою клейку, в’язку речовину. Ослизнення клітинних стінок спостерігається в клітинах:
кореневого чохлика (відбувається розшарування клітин і кінчик кореня ковзає по слизу, що полегшує його просування у ґрунті),
насіннєвих оболонок (у льону й айви слиз закріплює насіння на поверхні ґрунту),
ловчих апаратів комахоїдних рослин (у росянки слиз забезпечує приклеювання комах до ловчого апарату),
травмованих тканин (у вишні та сливи камедь (деревний клей) закриває пошкоджені ділянки стебел).
Мацерація
(від лат. macero – розм’якшую)
1.2.4.2.6.Мацерація – процес роз’єднання (відокремлення) клітин між собою, внаслідок руйнування серединної пластинки (загального шару клітинної стінки для двох сусідніх клітин). Мацерація характерна для клітин перезрілих плодів, деревини.
1.3. Життєвий цикл клітини– ряд послідовних перетворень, які проходить жива клітина від моменту утворення до наступного поділу на дві клітини або відмирання. У життєвому циклі клітини виділяють два періоди:
інтерфаза(період спокою клітини),
поділ клітини (мітоз, мейоз або амітоз).
Інтерфаза
(від лат. inter –
між і від гр. phasis –
поява) – фаза у якій добре видно ядро
і нема хромасом
1
Рис.21 Клітинний цикл:
КТ –
контрольна точка початку підготовки
клітини до поділу
У періоді G1, у знов утвореній клітині, відбуваються ростові процеси, збільшуються розміри клітини і кількість органел в ній. Якщо клітина не запрограмована на подальший поділ вона переходить у період G0 – фазу спеціалізованого функціонування, у якій клітина залишається до своєї загибелі.
Період S проходить лише в клітинах які у подальшому будуть ділитись. На цьому етапі в ядрі клітини відбувається подвоєння молекул ДНК (реплікація або редуплікація хроматину). Кожна молекула ДНК роз’єднується на два ланцюги, кожен із яких за принципом компліментарності із нуклеотидів, що містяться у ядерному соку, добудовує другу половинку, створюючи дві ідентичні (однакові) молекули ДНК.
У періоді G2 клітина готується до поділу, утворюються структури і компоненти необхідні для цього (наприклад: подвоюється кількість центріолей).
1.3.2. Поділ клітин – представляє собою процес перерозподілу генетичної інформації, цитоплазматичних структур (органел), гіалоплазми та фізіологічно активних і неактивних речовин материнської клітини у дочірні, що утворюються.
1
Мітоз
(від гр. mitos – нитка)
Профаза
(від грецької pro – до, перед і phasis –
поява)
Хромосома
(від гр. χρῶμα (chroma) – колір, фарба і
σῶμα (sōma) – тіло)
Хромати́да
(від гр. χρῶμα (chroma) – колір, фарба і
εἶδος (eidos) – вид) – структурний
елемент (половинка) хромосоми
Хромонема
(від гр. χρῶμα (chroma) – колір, фарба і
(nema) – нитка)
Профаза – перша фаза мітозу:
хроматинові нитки (ДНК) дегідротуються і скручуються у хромосоми,
зникають ядерця,
руйнується ядерна оболонка, каріоплазма змішується з гіалоплазмою,
центріолі розходяться до полюсів клітини;
Під час поділу клітини хроматинові нитки скручуються у компактну транспортабельну форму – хромосоми. Кожна хромосома складається із двох ідентичних (однакових) половинок – хроматид, які після поділу потраплять у різні дочірні клітини. У складі хроматиди може знаходитись від однієї до декількох хромонем – скручених молекул ДНК. Диплоїдний та гаплоїдний набори хромосом, гамологічні хромосоми, біваленти.
Рис.
23. Схема будови хромосоми.
А.
Внутрішня структура; Б. Загальний
вигляд:
1 –
дві хроматиди, 2 – хромонеми всередині
хроматид, 3 – хромоміри, 4 – матрикс,
5 – первинна перетяжка з центомірою,
6 – ядерце, 7 – супутник хромосоми
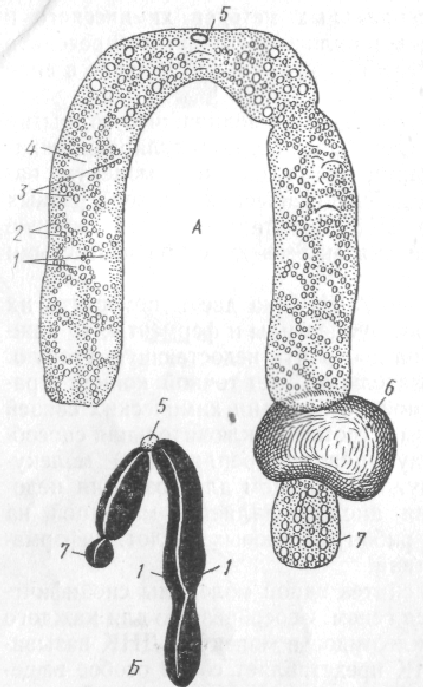
Рис. 22. Схема мітозу
М
Метафаза
(від гр. meta – між, після, через і
phasis –
поява)
Центроміра
(від лат. centrum – центр кола і від гр.
meros –
частина)
хромосоми збираються у екваторіальній площині клітини,
м
Анафаза (від гр. ana – до верху, до гори і phasis – поява)
ікротрубочки ростуть із центріолей формуючи спеціальну транспортну структуру –веретено поділу. Опорні мікротрубочки веретена поділу з’єднують центріолі на протилежних полюсах клітини, транспортні мікротрубочки з’єднуються з центромірами хроматид. Хроматиди кожної хромосоми прикріплюються до транспортних мікротрубочок, що ростуть з різних полюсів клітини;
Анафаза – третя фаза мітозу:
хромосоми діляться на дві хроматиди,
транспортні мікротрубочки скорочуються і відтягують приєднані до них хроматиди до різних полюсів клітини (половина хроматид до одного полюсу, інша – до другого);
Телофаза
(від гр.telos –
кінець і phasis –
поява)
Телофаза – четверта і заключна фаза мітозу:
половинки хромосом (хроматиди) групуються на полюсах материнської клітини і розкручуються у хроматин,
мікротрубочки веретена поділу розбираються (демонтуються),
навколо хроматину на полюсах клітини ЕПС будує ядерні оболонки (формуються два ядра),
відбувається цитокінез – розділення материнської клітини новою клітинною стінкою на дві дочірні.
Ц
Цитокінез
(від гр. kytos –
вмістилище, тут – клітина і kinesis –
рух)
Цитотомія
(від грецької kytos –
вмістилище, тут – клітина, tome -
розсічення)
Рис. 23.
Перерасподіл мікро трубочок на протязі
клітинного циклу і протягом утворення
клітинної стінки. А.
В інтерфазі, під час росту і диференціації
клітин, мікротрубочки лежать безпосередньо
під плазматичною мембраною.Б. Перед
профазою кільцевий пояс мікротрубочок
оточує ядро в екваторіальній площині
майбутнього мітотичного веретена. В.
В метафазі мікро трубочки формують
веретено поділу. Г. Коли мікро трубочки
веретена зникають, нові мікро трубочки
утворюють між двома дочірніми ядрами
фрагмопласт. Серединна пластинка
формується в екваторі фрагмопласту
між двома ядрами і росте центр обіжно,
доки не сягає оболонки клітини ,що
ділиться. Д. Кожна з дочірніх кітин
формує особисту первинну оболонку. Е.
У міру росту дочірніх клітин (тут
показана лише верхня) оболонка
материнської клітини руйнується. Д. і
Е. Мікро трубочки звов лежать під
плазматичною мембраною.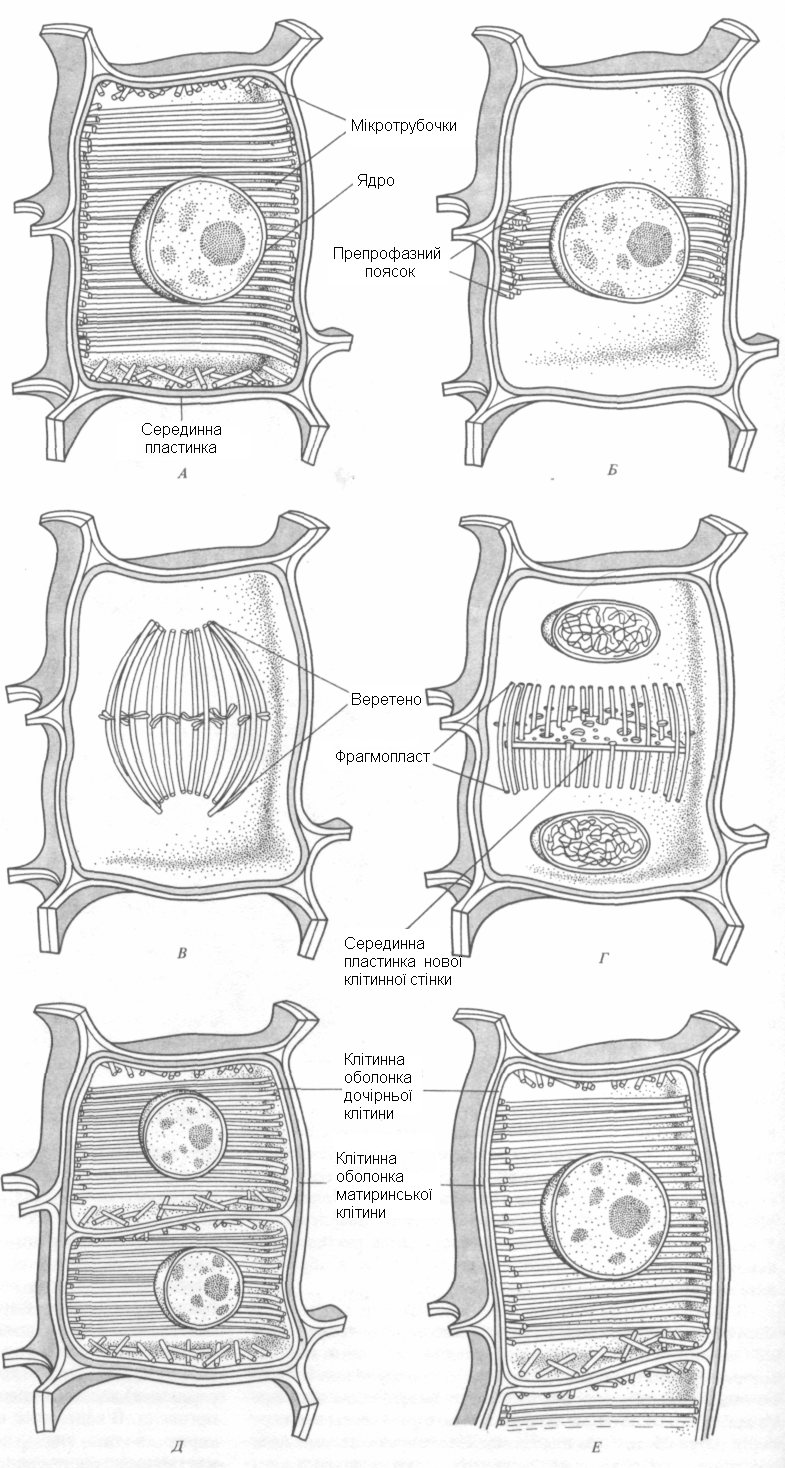
1
Мейоз
(від гр. meiosis –
зменшення)
етап І: профаза І, метафаза І, анафаза І і телофаза І;
етап ІІ: профаза ІІ, метафаза ІІ, анафаза ІІ і телофаза ІІ.
Рис. 24. Схема мейозу
(вказані не всі стадії)
Перший етапмейотичного поділу характеризується передачею у дочірні клітини половинного (гаплоїдного, зменшеного у двічі) набору хромосом. У материнській клітині за фазами поділу відбуваються наступні процеси:
Профаза І – перша фаза першого етапу мейозу:
руйнується ядерна оболонка і каріоплазма з’єднується з гіалоплазмою,
зникають ядерця,
хроматин скручується у хромосоми,
відбувається кон’югація – попарне з’єднання (переплетіння) гомологічних хромосом (пара гомологічних хромосом називається бівалентам),
між гомологічними хромосомами у бівалентах відбувається обмін ділянками (генами) – кросинговер;
Метафаза І – друга фаза першого етапу мейозу:
біваленти хромосом переміщуються у екваторіальну площину клітини,
із мікротрубочок формується веретено поділу,
до центромір гомологічних хромосом кріпляться транспортні мікротрубочки веретена поділу, що ростуть з різних полюсів клітини;
Анафаза І – третя фаза першого етапу мейозу:
біваленти роз’єднуються, транспортні мікротрубочки скорочуються і відтягують гомологічні хромосоми до різних полюсів клітини;
Телофаза І – четверта фаза першого етапу мейозу:
розбирається веретено поділу,
хромосоми розкручуються у хроматин (не у всіх клітин),
утворюються ядерця (не у всіх клітин),
ЕПС формує нові ядерні оболонки (не у всіх клітин),
відбувається цитокінез – розділення материнської клітини на дві дочірні.
Інтеркінез
(від лат. inter –
між і від гр. kinesis – рух)
Інтеркінез – період між першим і другим етапами мейозу, у якому клітина готується до другого поділу, утворюються структури необхідні для цього.
Другий етап мейотичного поділу відбувається за типом мітозу паралельно у двох дочірніх клітинах, що утворилися після першого етапу, а саме:
Профаза ІІ – перша фаза другого етапу мейозу:
зникають ядерця (як що вони були у телофазі І),
хроматин скручується у хромосоми (як що він утворився у телофазі І),
руйнуються ядерні оболонки у двох дочірніх клітинах і каріоплазма в них з’єднується з гіалоплазмою (як що вони були сформовані у телофазі І);
Метафаза ІІ – друга фаза другого етапу мейозу:
гаплоїдні хромосоми переміщуються у екваторіальну площину в двох дочірніх клітинах,
із мікротрубочок формуються веретена поділу,
до центромір хроматид гомологічних хромосом кріпляться транспортні мікротрубочки веретен поділу, що ростуть з різних полюсів клітини;
Анафаза ІІ – третя фаза другого етапу мейозу:
гомологічні хромосоми роз’єднуються на хроматиди, транспортні мікротрубочки скорочуються і відтягують хроматиди гомологічних хромосом до різних полюсів у двох дочірніх клітинах;
Телофаза ІІ – четверта фаза другого етапу мейозу:
розбираються веретена поділу,
хромосоми розкручуються у хроматин,
ЕПС формує нові ядерні оболонки (у двох дочірніх клітина утворюються по два ядра),
у дочірніх клітинах відбувається цитокінез – розділення кожної з низ на дві. Утворюється тетрада (четвірка) клітин з гаплоїдним набором хромосом.
1.3.2.3. Амітоз – простий поділ клітин, при якому ядро ділиться перетяжкою на дві частини не завжди рівномірні.