

биохимия (Восстановлен)
.pdf1.Предмет и задачи биологической химии. Биохимия как молекулярный уровень изучения структурной организации, анаболизма и катаболизма живой материи. Место биохимии среди других биологических дисциплин. Значение биохимии в подготовке врача и для медицины.
Биохимия – это наука о химическом составе живой материи, химических процессах, происходящих в живых организмах, а также связи этих превращений с деятельностью органов и тканей.
Таким образом, биохимия состоит как бы из трех частей:
1)статическая биохимия (это анализ химического состава живых организмов);
2)динамическая биохимия (изучает совокупность превращения веществ и энергии в организме);
3)функциональная биохимия (исследует процессы, лежащие в основе различных проявлений жизнедеятельности).
Главным для биохимии является выяснение функционального, то есть биологического назначения всех химических веществ и физико-химических процессов в живом организме, а также механизм нарушения этих функций при разных заболеваниях.
Современная биохимия решает следующие задачи:
1.Биотехнологическую, т.е. создание фармацевтических препаратов (гормонов, ферментов), регуляторов роста растений, средств борьбы с вредителями, пищевых добавок.
2.Проводит разработку новых методов и средств диагностики и лечения наследственных заболеваний, канцерогенеза, природы онкогенов и онкобелков.
3.Проводит разработку методов генной и клеточной инженерии для получения принципиально новых пород животных и форм растений с более ценными признаками.
4.Изучает молекулярные основы памяти, психики, биоэнергетики, питания и целый ряд других задач.
Биологическая химия изучает молекулярные процессы, лежащие в основе развития и функционирования организмов. Биохимия использует методы «молекулярных» наук — химии, физической химии, молекулярной физики, и в этом отношении биохимия сама является молекулярной наукой. Однако главные конечные задачи биохимии лежат в области биологии: она изучает закономерности биологической, а не химической формы движения материи. С другой стороны , «молекулярные изобретения» природы , открываемые биохимиками, находят применение в небиологических отраслях знания и в промышленности (молекулярная бионика, биотехнология). В таких случаях биохимия выступает в роли метода, а предметом исследований и разработок являются проблемы, выходящие за пределы биологии.
Живые организмы находятся в постоянной и неразрывной связи с окружающей средой. Эта связь осуществляется в процессе обмена веществ. Обмен веществ включает 3 этапа: поступление веществ в организм, метаболизм и выделение конечных продуктов из организма.
Поступление веществ в организм происходит в результате дыхания (кислород) и питания. В ЖКТ продукты питания перевариваются (расщепляются до простых веществ). При переваривании происходит гидролиз полимеров (белков, полисахаридов и других сложных органических веществ) до мономеров, всасывающихся в кровь и включающихся в промежуточный обмен.
Промежуточный обмен (внутриклеточный метаболизм) включает 2 типа реакций: катаболизм и анаболизм.
Катаболизм - процесс расщепления органических молекул до конечных продуктов. Конечные продукты превращений органических веществ у животных и человека - СО2, Н2О и мочевина. В процессы катаболизма включаются метаболиты, образующиеся как при пищеварении, так и при распаде структурнофункциональных компонентов клеток.
Реакции катаболизма сопровождаются выделением энергии (экзергонические реакции).
Анаболизм объединяет биосинтетические процессы, в которых простые строительные блоки соединяются в сложные макромолекулы, необходимые для организма. В анаболических реакциях используется энергия, освобождающаяся при катаболизме (эндергонические реакции).
Практически любое заболевание начинается с повреждения (нарушения) одной реакции в метаболизме клетки, а затем оно распространяется на ткань, орган и целый организм. Нарушение метаболизма ведет к нарушению гомеостаза в биологических жидкостях организма человека, что сопровождается изменением биохимических показателей.
Большое значение клинико-биохимических методов исследования биологических жидкостей велико в медицине и важно для подготовки медицинских лабораторных техников. Достаточно напомнить, что только в крови человека можно определить современными методами биохимических исследований около 1000 показателей метаболизма.
Биохимические показатели биологических сред организма человека широко используются при:
1.постановке диагноза заболевания, особенно дифференциального диагноза;
2.выборе метода лечения;
3.контроле за правильностью назначенного лечения; 4.результаты биохимических анализов служат одним из критериев излеченности патологического процесса; 5.скрининге (выявлении болезни на доклинической стадии);
6.мониторинге (контроле за течением заболевания и результатом лечения); 7. прогнозе (информации о возможном исходе заболевания).
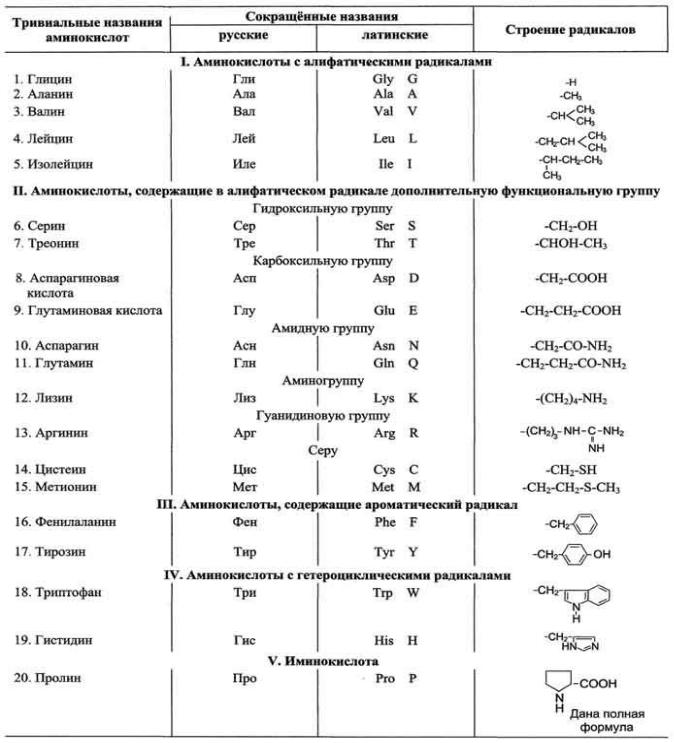
2. Аминокислоты, входящие в состав белков, их строение и свойства. Пептиды. Биологическая роль аминокислот и пептидов.
1. Общие структурные особенности аминокислот, входящих в состав белков
Общая структурная особенность аминокислот - наличие амино- и карбоксильной групп, соединѐнных с одним и тем же ?-углеродным атомом. R - радикал аминокислот - в простейшем случае представлен атомом водорода (глицин), но может иметь и более сложное строение. В водных растворах при нейтральном значении рН ?- аминокислоты существуют в виде биполярных ионов. В отличие от 19 остальных ?- аминокислот, пролин - иминокислота, радикал которой связан как с ?-углеродным атомом, так и с аминогруппой, в результате чего молекула приобретает циклическую структуру.
19 из 20 аминокислот содержат в ?-положении асимметричный атом углерода, с которым связаны 4 разные замещающие группы. В результате эти аминокислоты в природе могут находиться в двух разных изомерных формах - L и D. Исключение составляет глицин, который не имеет асимметричного ?- углеродного атома, так как его радикал представлен только атомом водорода. В составе белков присутствуют только L-изомеры аминокислот.
Чистые L- или D-стереоизомеры могут за длительный срок самопроизвольно и неферментативно превращаться в эквимолярную смесь L- и D-изомеров. Этот процесс называют рацемизацией. Рацемизация каждой L-аминокислоты при данной температуре идѐт с определѐнной скоростью. Все 20 аминокислот в организме человека различаются по строению, размерам и физико-химическим свойствам радикалов, присоединѐнных к ?-углеродному атому.
2. Классификация аминокислот по химическому строению радикалов
По химическому строению аминокислоты можно разделить на алифатические, ароматические и гетероциклические
В составе алифатических радикалов могут находиться функциональные группы, придающие им специфические свойства: карбоксильная (-СООН), амино (-NH2), тиольная (-SH), амидная (-CO-NH2),
гидроксильная |
(-ОН) |
и |
гуанидиновая группы. |
Для записи аминокислотных остатков в молекулах пептидов и белков используют трѐхбуквенные сокращения их тривиальных названий, а в некоторых случаях и однобуквенные символы
3. Классификация аминокислот по растворимости их радикалов в воде
Все 20 аминокислот в белках организма человека можно сгруппировать по способности их радикалов растворяться в воде. Радикалы можно выстроить в непрерывный ряд, начинающийся полностью гидрофобными и заканчивающийся сильно гидрофильными.
Растворимость радикалов аминокислот определяется полярностью функциональных групп, входящих в состав молекулы (полярные группы притягивают воду, неполярные еѐ отталкивают).
Аминокислоты с неполярными радикалами
К неполярным (гидрофобным) относят радикалы, имеющие алифатические углеводородные цепи (радикалы аланина, валина, лейцина, изолейцина, пролина и метионина) и ароматические кольца (радикалы фенилаланина и триптофана). Радикалы таких аминокислот в воде стремятся друг к другу или к другим гидрофобным молекулам, в результате чего поверхность соприкосновения их с водой уменьшается.
Аминокислоты с полярными незаряженными радикалами
Радикалы этих аминокислот лучше, чем гидрофобные радикалы, растворяются в воде, так как в их состав входят полярные функциональные группы, образующие водородные связи с водой. К ним относят серии, треонин и тирозин, имеющие гидроксильные группы, аспарагин и глутамин, содержащие амидные группы, и цистеин с его тиольной группой.
Аминокислоты с полярными отрицательно заряженными радикалами
К этой группе относят аспарагиновую и глутаминовую аминокислоты, имеющие в радикале дополнительную карбоксильную группу, при рН около 7,0 диссоциирующую с образованием СОО- и Н+. Следовательно, радикалы данных аминокислот - анионы. Ионизированные формы глутаминовой и аспарагиновой кислот называют соответственно глутаматом и аспартатом.
Аминокислоты с полярными положительно заряженными радикалами
Дополнительную положительно заряженную группу в радикале имеют лизин и аргинин. У лизина вторая аминогруппа, способная присоединять Н+, располагается в ?-положении алифатической цепи, а у аргинина положительный заряд приобретает, гуанидиновая группа, Кроме того, гистидин содержит слабо ионизированную имидазольную группу, поэтому при физиологических колебаниях значений рН (от 6,9 до 7,4) гистидин заряжен либо нейтрально, либо положительно. При увеличении количества протонов в среде имидазольная группа гистидина способна присоединять протон, приобретая положительный заряд, а при увеличении концентрации гидроксильных групп - отдавать протон, теряя положительный заряд радикала. Положительно заряженные радикалы - катионы .Наибольшей растворимостью в воде обладают полярные заряженные радикалы аминокислот.
4. Изменение суммарного заряда аминокислот в зависимости от рН среды
При нейтральных значениях рН все кислотные (способные отдавать Н+) и все основные (способные присоединять Н+) функциональные группы находятся в диссоциированном состоянии.
Поэтому в нейтральной среде аминокислоты, содержащие недиссоциирующий радикал, имеют суммарный нулевой заряд. Аминокислоты, содержащие кислотные функциональные группы, имеют суммарный отрицательный заряд, а аминокислоты, содержащие основные функциональные группы, - положительный заряд
Изменение рН в кислую сторону (т.е. повышение в среде концентрации Н+) приводит к подавлению диссоциации кислотных групп. В сильно кислой среде все аминокислоты приобретают положительный заряд.
Напротив, увеличение концентрации ОН- групп вызывает отщепление Н+ от основных функциональных групп, что приводит к уменьшению положительного заряда. В сильно щелочной среде все аминокислоты имеют суммарный отрицательный заряд.
5. Модифицированные аминокислоты, присутствующие в белках
Непосредственно в синтезе белков организма человека принимают участие только 20 перечисленных аминокислот. Однако в некоторых белках имеются нестандартные модифицированные аминокислоты - производные одной из этих 20 аминокислот.
Модификации аминокислотных остатков осуществляются уже в составе белков, т.е. только после окончания их синтеза. Введение дополнительных функциональных групп в структуру аминокислот придаѐт белкам свойства, необходимые для выполнения ими специфических функций.
6. Химические реакции, используемые для обнаружения аминокислот
Для обнаружения и количественного определения аминокислот, находящихся в растворе, можно использовать нингидриновую реакцию.
Эта реакция основана на том, что бесцветный нингидрин, реагируя с аминокислотой, конденсируется в виде димера через атом азота, отщепляемый от ?-аминогруппы аминокислоты. В результате образуется пигмент красно-фиолетового цвета. Одновременно происходит декарбоксилирование аминокислоты, что приводит к образованию СО2 и соответствующего альдегида. Нингидриновую реакцию широко используют при изучении первичной структуры белков Так как интенсивность окраски пропорциональна количеству аминокислот в растворе, еѐ используют для измерения концентрации ?-аминокислот.
Специфические реакции на отдельные аминокислоты
Качественное и количественное определение отдельных аминокислот возможно благодаря наличию в их радикалах особенных функциональных групп.
Аргинин определяют с помощью качественной реакции на гуанидиновую группу (реакция Сакагучи), а цистеин выявляют реакцией Фоля, специфичной на SH-группу данной аминокислоты. Наличие ароматических аминокислот в растворе определяют ксантопротеиновой реакцией (реакция нитрования), а наличие гидроксильной группы в ароматическом кольце тирозина - с помощью реакции Миллона.
Б. Пептидная связь. Строение и биологические свойства пептидов
3.Биологическая роль пептидов
В организме человека вырабатывается множество пептидов, участвующих в регуляции различных биологических процессов и обладающих высокой физиологической активностью.
Функции пептидов зависят от их первичной структуры. Ангиотензин I по структуре очень похож на ангиотензин II (имеет только две дополнительные аминокислоты с С-конца), но при этом не обладает биологической активностью.
Изменение в аминокислотном составе пептидов часто приводит к потере одних и возникновению других биологических свойств.
Так как пептиды - мощные регуляторы биологических процессов, их можно использовать как лекарственные препараты. Основное препятствие для терапевтического использования - их быстрое разрушение в организме. Одним из важнейших результатов исследований является не только изучение структуры пептидов, но и получение синтетических аналогов природных пептидов с целенаправленными изменениями в их структуре и функциях.
Открытые и изученные в настоящее время пептиды можно разделить на группы по их основному физиологическому действию:
пептиды, обладающие гормональной активностью (окситоцин, вазопрессин, рилизинг-гормоны гипоталамуса, меланоцитстимулирующий гормон, глюкагон и др.);
пептиды, регулирующие процессы пищеварения (гастрин, холецистокинин, вазоинтестиналшый пептид, желудочный ингибирующий пептид и др.);
пептиды, регулирующие тонус сосудов и АД (брадикинин, калидин, ангиотензин II);
пептиды, регулирующие аппетит (лептин, нейропептид Y, меланоцитстимулирующий гормон, (?- эндорфины);

пептиды, обладающие обезболивающим действием (энкефалины и эндорфины и другие опиоидные пептиды). Обезболивающий эффект этих пептидов в сотни раз превосходит анальгезирующий эффект морфина;
пептиды, участвующие в регуляции высшей нервной деятельности, в биохимических процессах, связанных с механизмами сна, обучения, памяти, возникновения чувства страха и т.д.
3. Первичная структура белков. Пептидная связь, ее характеристика (прочность, кратность, компланарность, цис- ,транс- изомерия). Значение первичной структуры для нормального функционирования белков (на примере гемоглобина S).
Первичной структурой белков называется линейная полипептидная цепь из аминокислот, соединенных между собой пептидными связями. Первичная структура - простейший уровень структурной организации белковой молекулы. Высокую стабильность ей придают ковалентные пептидные связи между α-аминогруппой одной аминокислоты и α-карбоксильной группой другой аминокислоты
Если в образовании пептидной связи участвует иминогруппа пролина или гидроксипролина, то она имеет другой вид .
При образовании пептидных связей в клетках сначала активируется карбоксильная группа одной аминокислоты, а затем она соединяется с аминогруппой другой. Примерно так же проводят лабораторный синтез полипептидов.
Пептидная связь является повторяющимся фрагментом полипептидной цепи. Она имеет ряд особенностей, которые влияют не только на форму первичной структуры, но и на высшие уровни организации полипептидной цепи:
копланарность - все атомы, входящие в пептидную группу, находятся в одной плоскости;
способность существовать в двух резонансных формах (кетоили енольной форме);
транс-положение заместителей по отношению к С-N-связи;
способность к образованию водородных связей, причем каждая из пептидных групп может образовывать две водородные связи с другими группами, в том числе и пептидными.
Исключение составляют пептидные группы с участием аминогруппы пролина или гидроксипролина. Они способны образовывать только одну водородную связь . Это сказывается на формировании вторичной структуры белка. Полипептидная цепь на участке, где находится пролин или гидроксипролин, легко изгибается, так как не удерживается, как обычно, второй водородной связью.
Особенности первичной структуры белка. В остове полипептидной цепи чередуются жесткие структуры (плоские пептидные группы) с относительно подвижными участками (—СНR), которые способны вращаться вокруг связей. Такие особенности строения полипептидной цепи влияют на укладку ее в пространстве.
2.Характеристика пептидной связи
Пептидная связь имеет характеристику частично двойной связи, поэтому она короче, чем остальные связи пептидного остова, и вследствие этого мало подвижна. Электронное строение пептидной связи определяет плоскую жѐсткую структуру пептидной группы. Плоскости пептидных групп расположены под углом друг к другу .
Связь между ?-углеродным атомом и ?-аминогруппой или ?-карбоксильной группой способна к свободным вращениям (хотя ограничена размером и характером радикалов), что позволяет полипептидной цепи принимать различные конфигурации.
Пептидные связи обычно расположены в транс-конфигурации, т.е. ?-углеродные атомы располагаются по разные стороны от пептидной связи. В результате боковые радикалы аминокислот находятся на наиболее удалѐнном расстоянии друг от друга в пространстве.
Пептидные связи очень прочны и самопроизвольно не разрываются при нормальных условиях, существующих в клетках (нейтральная среда, температура тела). В лабораторных условиях гидролиз пептидных связей белков проводят в запаянной ампуле с концентрированной (6 моль/л) соляной кислотой,

при температуре более 105 °С, причѐм полный гидролиз белка до свободных аминокислот проходит примерно за сутки.
В живых организмах пептидные связи в белках разрываются с помощью специальных протеолитических ферментов (от англ, protein - белок, lysis - разрушение), называемых также протеазами, или пептидгидролазами.
Для обнаружения в растворе белков и пептидов, а также для их количественного определения используют биуретовую реакцию (положительный результат для веществ, содержащих в своѐм составе не менее двух пептидных связей).
Химическая природа каждого белка уникальна и тесно связана с его биологической функцией. Способность белка выполнять присущую ему функцию определяется его первичной структурой. Даже небольшие изменения в последовательности аминокислот в белке могут привести к серьезному нарушению в его функционировании, возникновению тяжелого заболевания. Болезни, связанные с нарушениями первичной структуры белка, получили название молекулярных. К настоящему времени открыто несколько тысяч таких болезней. Одной из молекулярных болезней является серповидноклеточная анемия, причина которой кроется в нарушении первичной структуры гемоглобина. У людей с врожденной аномалией структуры гемоглобина в полипептидной цепочке, состоящей из 146 аминокислотных остатков, в шестом положении находится валин, тогда как у здоровых людей на этом месте — глутаминовая кислота. Аномальный гемоглобин хуже транспортирует кислород, а эритроциты крови больных имеют серповидную форму. Заболевание проявляется в замедлении развития, общей слабости организма.
4. Вторичная структура белков. Связи, стабилизирующие вторичную структуру.
Вторичная структура - это пространственное расположение полипептидной цепочки в виде α-спирали или β-складчатости безотносительно к типам боковых радикалов и их конформации. Она стабилизирована водородными связями, которые замыкаются между пептидными, амидными (-N-H) и карбонидными (- C=O)группами, т.е. входят в пептидную единицу, и дисульфидными мостиками между остатками цистеина
Полинг и Кори предложили модель вторичной структуры белка в виде левозакрученной α-спирали, в которой водородные связи замыкаются между каждой первой и четвертой аминокислотой, что позволяет сохранять нативную структуру белка, осуществление им простейших функций, защищать от разрушения. На один виток спирали приходится 3,6 аминокислотных остатка, шаг спирали составляет 0,54 нм. В образовании водородных связей принимают участие все пептидные группы, что обеспечивает максимальную стабильность, снижает гидрофильность и увеличивает гидрофобность белковой молекулы. Альфа-спираль образуется самопроизвольно и является наиболее устойчивой конформацией, отвечающей минимуму свободной энергии .
Полинг и Кори предложили и другую упорядоченную структуру - складчатый β- слой. В отличие от конденсированной α-спирали β- слои почти полностью вытянуты и могут располагаться как параллельно , так и антипараллельно
В стабилизации данных структур также принимают участие дисульфидные мостики и водородные связи.
5. Третичная структура белков. Типы химических связей, участвующих в формировании третичной структуры. Супервторичная структура. Доменная структура и ее роль в функционировании белков. Роль шаперонов (белки теплового шока) в формировании третичной структуры белков in vivo.
У большинства белков полипептидные цепи свернуты особым образом в компактную глобулу. Способ свертывания полипептидных цепей глобулярных белков называется третичной структурой. Третичная структура поддерживается уже обсуждавшимися выше связями трех типов — ионными, водородными и дисульфидными, а также гидрофобными взаимодействиями. В количественном отношении наиболее важны именно гидрофобные взаимодействия; белок при этом свертывается таким образом, чтобы его гидрофобные боковые цепи были скрыты внутри молекулы, а гидрофильные, наоборот, выставлены наружу
Супервторичная структура - это более высокий уровень организации белковой молекулы, представленный ансамблем взаимодействующих между собой вторичных структур
Некоторый специфический порядок чередования вторичных структур наблюдается во многих разных по структуре и функциям белках и носит название супервторичной структуры.
Такие упорядоченные структуры часто обозначают как структурные мотивы, которые имеют специфические названия: «а-спираль—поворот—а-спи-раль», «лейциновая застежка-молния», «цинковые пальцы», «структура Р-бочонка» и др.
По наличию а-спиралей и b-структур глобулярные белки могут быть разделены на 4 категории:
1.В первую категорию включены белки, в которых имеются только а-спирали, например миоглобин и гемоглобин .
2.Во вторую категорию включены белки, в которых имеются а-спирали и b-структуры. ЛДГ.
3.В третью категорию включены белки, имеющие только вторичную b-структуру. Такие структуры обнаружены в иммуноглобулинах, в ферменте супероксиддисмутазе
4.В четвертую категорию включены белки, имеющие в своем составе лишь незначительное количество регулярных вторичных структур. К таким белкам можно отнести небольшие богатые цистином белки или металлопротеины.
В ДНК-связывающих белках имеются общие виды супервторичных структур: «спираль—поворот— спираль», «лейциновая застежка-молния», «цинковые пальцы». ДНК-связывающие белки содержат центр связывания, комплементарный участку ДНК с определенной нуклеотидной последовательностью. Эти белки участвуют в регуляции действия генов.
«а- Спираль—поворот—а-спираль»
Двуспиральная структура ДНК имеет 2 бороздки: большую и малую. Большая бороздка хорошо приспособлена для связывания белков, имеющих небольшие спиральные участки.
В данный структурный мотив входят 2 ос-спирали: одна более короткая, другая более длинная, соединенные поворотом полипептидной цепи.
Более короткая а-спираль располагается поперек бороздки ДНК, а более длинная а-спираль находится в большой бороздке, образуя нековалентные специфические связи радикалов аминокислот с нуклеотидами ДНК.
«Цинковый палец»
«Цинковый палец» — фрагмент белка, содержащий около 20 аминокислотных остатков.
Атом цинка связан с радикалами 4 аминокислот: 2 остатков цистеина и 2 — гистидина.
В некоторых случаях вместо остатков гистидина находятся остатки цистеина.
Этот участок белка образует а-спираль, которая может специфично связываться с регуляторными участками большой бороздки ДНК.
Специфичность связывания индивидуального регуляторного ДНК-связывающего белка зависит от последовательности аминокислотных остатков, расположенных в области «цинкового пальца».
«Лейциновая застежка-молния»
Взаимодействующие белки имеют а-спиральный участок, содержащий по крайней мере 4 остатка лейцина.
Лейциновые остатки расположены через 6 аминокислот один от другого.
Так как каждый виток а-спирали содержит 3,6-аминокислотного остатка, радикалы лейцина находятся на поверхности каждого второго витка.
Лейциновые остатки а-спирали одного белка могут взаимодействовать с лейциновыми остатками другого белка (гидрофобные взаимодействия), соединяя их вместе .
Многие ДНК-связывающие белки взаимодействуют с ДНК в виде олигомерных структур, где субъединицы связываются друг с другом «лейциновыми застежками». Примером таких белков могут служить гистоны.
Гистоны — ядерные белки, в состав которых входит большое количество положительно заряженных аминокислот — аргинина и лизина (до 80%).
Молекулы гистонов объединяются в олигомерные комплексы, содержащие 8 мономеров с помощью «лейциновых застежек», несмотря на сильный положительный заряд этих молекул.
ДОМЕННАЯ СТРУКТУРА И ЕЕ РОЛЬ В ФУНКЦИОНИРОВАНИИ БЕЛКОВ 1. Длинные полипептидные цепи часто складываются в несколько компактных, относительно независимых

областей. Они имеют самостоятельную третичную структуру, напоминающую таковую глобулярных
белков, |
|
и |
|
|
|
называются |
|
|
доменами. |
Благодаря |
доменной |
структуре |
белков |
легче |
формируется |
их |
трехмерная |
структура. |
|
2. Центры |
связывания |
белка с |
лигандом часто располагаются между доменами (например, центр |
||||||
связывания трипсина с его лигандом - пищевым белком). Разные домены в белке могут перемещаться относительно друг друга при взаимодействии с лигандом (например, в молекуле гексокиназы). В некоторых белках домены выполняют самостоятельные функции, связываясь с различными лигандами. Такие белки называются многофункциональными белками.
Правильное сворачивание ( фолдинг ) полипептидных цепей белков в клетках эукариот обеспечивается специфическими белками, называемыми шаперонами (chaperone). Шапероны необходимы для
эффективного формирования третичной структуры полипептидных цепей других белков, но они не входят в
состав конечной белковой структуры.
Новосинтезированные белки после выхода с рибосом для правильного функционирования должны
укладываться в стабильные трехмерные структуры и оставаться такими на протяжении всей
функциональной жизни клетки. Поддержание контроля качества структуры белка и осуществляется
шаперонами, катализирующими укладку полипептидов. Сборка полипротеинов и укладка мультибелковых
комплексов также осуществляется шаперонами. Шапероны связываются с гидрофобными участками
неправильно уложенных белков, помогают им свернуться и достигнуть стабильной нативной структуры и,
тем самым, предотвращают их включение в нерастворимые и нефункциональные агрегаты. В течение своей
функциональной жизни белок может подвергаться различным стрессам и денатурации. Такие частично
денатурированные белки могут стать, во-первых, мишенью протеаз, во-вторых, агрегировать и, в-
третьих, укладываться в нативную структуру с помощью шаперонов. Баланс и эффективность, с которой
происходят эти три процесса, определяются соотношением компонентов, участвующих в этих реакциях
6. Активный центр белков и его специфическое взаимодействие с лигандом как основа биологической функции белков. Конформационная лабильность белков.Комплементарность взаимодействия белков с лигандами. Обратимость связывания.
Активный центр белков - определѐнный участок белковой молекулы, как правило, находящийся в еѐ углублении ("кармане"), сформированный радикалами аминокислот, собранных на определѐнном пространственном участке при формировании третичной структуры и способный комплементарно связываться с лигандом. В линейной последовательности полипептидной цепи радикалы, формирующие активный центр, могут находиться на значительном расстоянии друг от друга.
Высокая специфичность связывания белка с лигандом обеспечивается комплементарностью структуры активного центра белка структуре лиганда
Под комплементарностью понимают пространственное и химическое соответствие взаимодействующих молекул. Лиганд должен обладать способностью входить и пространственно совпадать с конформацией активного центра. Это совпадение может быть неполным, но благодаря конформационной лабильности белка активный центр способен к небольшим изменениям и "подгоняется" под лиганд. Кроме того, между функциональными группами лиганда и радикалами аминокислот, образующих активный центр, должны возникать связи, удерживающие лиганд в активном центре. Связи между лигандом и активным центром белка могут быть как нековалентными (ионными, водородными, гидрофобными), так и ковалентными.
1. Характеристика активного центра
Активный центр белка - относительно изолированный от окружающей белок среды участок, сформированный аминокислотными остатками. В этом участке каждый остаток благодаря своему индивидуальному размеру и функциональным группам формирует "рельеф" активного центра.
Объединение таких аминокислот в единый функциональный комплекс изменяет реакционную способность их радикалов, подобно тому, как меняется звучание музыкального инструмента в ансамбле. Поэтому аминокислотные остатки, входящие в состав активного центра, часто называют "ансамблем" аминокислот.
Уникальные свойства активного центра зависят не только от химических свойств формирующих его аминокислот, но и от их точной взаимной ориентации в пространстве. Поэтому даже незначительные нарушения общей конформации белка в результате точечных изменений его первичной структуры или условий окружающей среды могут привести к изменению химических и функциональных свойств радикалов, формирующих активный центр, нарушать связывание белка с лигандом и его функцию. При денатурации активный центр белков разрушается, и происходит утрата их биологической активности.
Часто активный центр формируется таким образом, что доступ воды к функциональным группам его радикалов ограничен, т.е. создаются условия для связывания лиганда с радикалами аминокислот.
В некоторых случаях лиганд присоединяется только к одному из атомов, обладающему определѐнной реакционной способностью, например присоединение О2 к железу миоглобина или гемоглобина. Однако свойства данного атома избирательно взаимодействовать с О2 определяются свойствами радикалов, окружающих атом железа в составе тема. Гем содержится и в других белках, таких как цитохромы. Однако функция атома железа в цитохромах иная, он служит посредником для передачи электронов от одного вещества другому, при этом железо становится то двух-, то трѐхвалентным.
Основное свойство белков, лежащее в основе их функций, - избирательность присоединения к определѐнным участкам белковой молекулы специфических лигандов.
2.Многообразие лигандов
Лигандами могут быть неорганические (часто ионы металлов) и органические вещества, низкомолекулярные и высокомолекулярные вещества;
существуют лиганды, которые изменяют свою химическую структуру при присоединении к активному центру белка (изменения субстрата в активном центре фермента);
существуют лиганды, присоединяющиеся к белку только в момент функционирования (например,
О2, транспортируемый гемоглобином), и лиганды, постоянно связанные с белком, выполняющие вспомогательную роль при функционировании белков (например, железо, входящее в состав гемоглобина).
В тех случаях, когда аминокислотные остатки, формирующие активный центр, не могут обеспечить функционирование данного белка, к определѐнным участкам активного центра могут присоединяться небелковые молекулы. Так, в активном центре многих ферментов присутствует ион металла (кофактор) или органическая небелковая молекула (кофермент). Небелковую часть, прочно связанную с активным центром белка и необходимую для его функционирования, называют "простатическая группа". Миоглобин, гемоглобин и цитохромы имеют в активном центре простетическую группу - гем, содержащий железо .
Соединение протомеров в олигомерном белке - пример взаимодействия высокомолекулярных лигандов. Каждый протомер, соединѐнный с другими протомерами, служит для них лигандом, так же как они для него.
Иногда присоединение какого-либо лиганда изменяет конформацию белка, в результате чего формируется центр связывания с другими лигандами. Например, белок кальмодулин после связывания с четырьмя ионами Са2+ в специфических участках приобретает способность взаимодействовать с некоторыми ферментами, меняя их активность.
7. Четвертичная структура белков. Особенности строения и функционирования олигомерных белков на примере гемоглобина. Кооперативные изменения конформации протомеров. Возможность регуляции биологической функции олигомерных белков аллостерическими лигандами.
Под четвертичной структурой подразумевают способ укладки в пространстве отдельных полипептидных цепей, обладающих одинаковой (или разной) первичной, вторичной или третичной структурой, и формирование единого в структурном и функциональном отношениях макромолекулярного образования. Многие функциональные белки состоят из нескольких полипептидных цепей, соединенных не