

Розділ 7. Обмін нуклеотидів
7.1. Азотисті основи як складові компоненти нуклеїнових кислот
У кожному живому організмі присутні 2 типи нуклеїнових основ: рибонуклеїнова (РНК) та дезоксирибонуклеїнова (ДНК). Вони характеризуються унікальними структурними та функціональними властивостями і являють собою складні біополімери клітин, які містять велику кількість мономерних одиниць – нуклеотидів, тому нуклеїнові кислоти називають полінуклеотидами.
При повному гідролізі нуклеїнових кислот у гідролізаті виявляють пуринові та піримідинові основи, вуглеводи та фосфорну кислоту (табл.7.1).
Таблиця 7.1. Особливості будови нуклеїнових кислот
|
|
ДНК |
РНК |
|
Азотисті основи |
аденін, гуанін, цитозин, тимін |
аденін, гуанін, цитозин, урацил |
|
Вуглевод |
дезоксирибоза |
рибоза |
|
Структура |
дволанцюгова |
одноланцюгова |
|
Молекулярна маса |
від десятків до сотень млн. да |
від десятків до сотень тисяч Да |
|
Локалізація в клітині |
ядро, мітохондрії, тилакоїди |
ядро, цитоплазма |
|
Функції |
збереження генетичної інформації |
передача генетичної інформації |
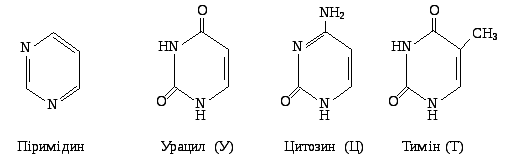
Азотисті основи, які входять до складу нуклеїнових кислот, є похідними гетероциклічних сполук - піримідину та пурину:

Однією з важливих властивостей азотистих основ є їх можливість існувати в двох таутомерних формах (лактимній і лактамній), у залежності від значення рН середовища. Таутомерні форми можна показати на прикладі урацилу:
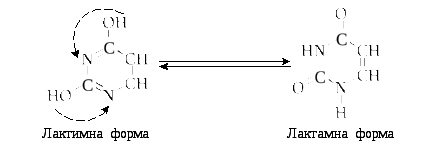
У складі нуклеїнових кислот всі оксипохідні пуринів і піримідинів знаходяться в лактамній формі.
Крім основних азотистих основ, у невеликій кількості зустрічаються рідкісні, так звані, мінорні основи: 5-метил- і 5-гідроксиметилцитозин, 5-карбоксиурацил, 4-тіоурацил, диметиламіноаденін, 1-метилгуанін:
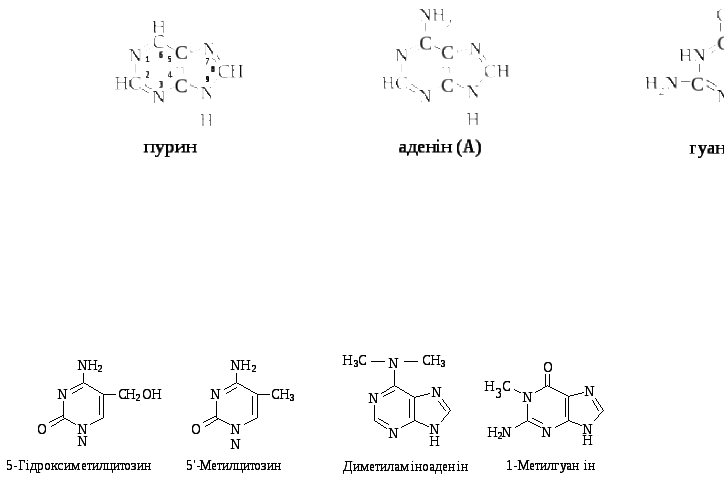
Лише в одній тРНК знаходиться приблизно 50 мінорних азотистих основ, їх роль, очевидно, зводиться до захисту молекули РНК від дії гідролітичних ферментів. Азотисті основи поглинають світло в ультрафіолетовій частині спектра з довжиною хвилі 200 – 300 нм і максимумом біля 260 нм. Ця властивість використовується при кількісному визначенні нуклеїнових кислот.
Деякі синтетичні похідні пуринів і піримідинів використовують як лікарські засоби. За будовою вони подібні на природні метаболіти (У, Т, Ц, А, Г), але не зовсім ідентичні (антиметаболіти). Наприклад, 5-фторурацил виступає як антагоніст урацилу і тиміну, а 6-меркаптопурин - аденіну. Конкуруючи з метаболітами, вони порушують синтез нуклеїнових кислот в організмі, що використовується при лікуванні пухлин.
Нуклеозиди.Азотисті основи, з’єднуючись з пентозами, утворюють сполуки, які називаються нуклеозидами. До складу нуклеїнових кислот входять лише-нуклеозиди.

Пуринові основи через дев’ятий атом азоту, а піримідинові через перший - утворюють N-глікозидний зв’язок з рибозою або дезоксирибозою. При цьому завжди утворюється -глікозидний зв’язок (за рахунок відщеплення молекули води).
У залежності від природи пентози (рибоза або дезоксирибоза) нуклеозиди поділяють на рибонуклеозиди та дезоксирибонуклеозиди.
Н
Рис.7.1.Механізм утворення
моно-, ди- та тринуклеотидів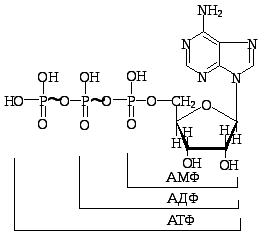
Мононуклеотиди та їх похідні, а також динуклеотиди наявні в клітинах у вільному стані та відіграють важливу роль в обміні речовин. Мононуклеотиди, приєднуючи ще один залишок фосфату, утворюють фосфоангідридний зв'язок і перетворюються на нуклеозиддифосфати (АДФ, ГДФ, УДФ, ТДФ і ЦДФ), останні, приєднуючи ще один залишок фосфату, утворюють нуклеозидтрифосфати (АТФ, ГТФ, УТФ, ТТФ і ЦТФ). Слід зазначити, що лише вільні нуклеозидтрифосфати в клітинах є попередниками ферментативного синтезу ДНК і РНК. У клітинах, однак, містяться й такі нуклеозидтрифосфати, які не беруть участі в синтезі білка, але виконують життєво важливі функції. Наприклад, АТФ бере участь в енергетичному обміні, є однією з макроергічних сполук (рис. 7.1). При відщепленні від АТФ однієї або двох молекул фосфатної кислоти, які зв’язані між собою макроергічними зв’язками (), виділяється 32-42 кДж/моль енергії, тоді як енергія звичайного фосфатного зв’язку 8-12 кДж/моль. В обміні речовин і енергії беруть участь й інші фосфорильовані нуклеотиди: ГТФ, ЦТФ, УТФ тощо.
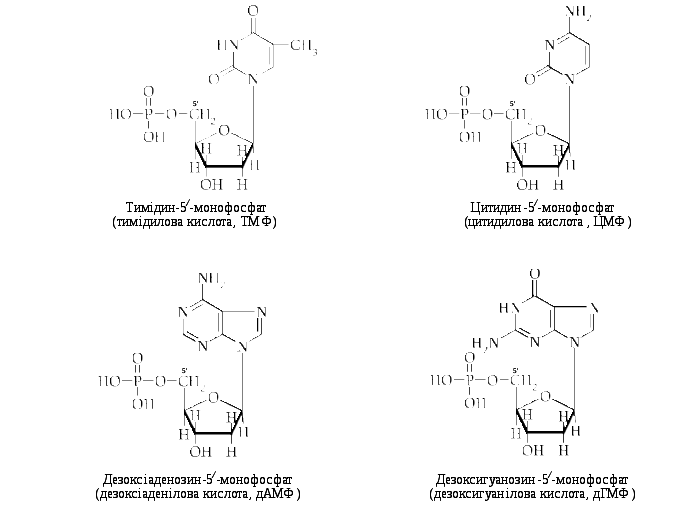
Номенклатура нуклеотидів представлена в таблиці 7.2.
Таблиця 7.2. Номенклатура нуклеотидів
|
Азотиста основа |
Нуклеозид |
Нуклеотид |
Трибуквове позначення |
Однобуквове позначення |
|
Аденін Гуанін Цитозин Урацил Тимін |
Аденозин Гуанозин Цитидин Уридин Тимідин |
Аденозинмонофосфат Гуанозинмонофосфат Цитидинмонофосфат Уридинмонофосфат Тимідинмонофосфат |
АМФ ГМФ ЦМФ УМФ ТМФ |
A G C U T |
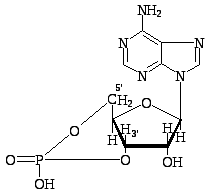
К
Аденозин 3',5'-циклофосфат
(цАМФ)
Синтез нуклеїнових кислот, мономерними одиницями яких є мононуклеотиди, визначатиметься швидкістю синтезу пуринових і піримідинових нуклеотидів; синтез останніх в свою чергу залежить від наявності всіх складових із трьох компонентів. Джерелом рибози і дезоксирибози служать продукти перетворення глюкози в пентозофосфатному циклі. Фосфатна кислота також не є лімітуючим чинником, оскільки вона поступає в достатній кількості з їжею. Отже, біосинтез нуклеїнових кислот починається з синтезу азотистих основ (мононуклеотидів).