|
Тема 5. Экология популяций
5.1. Принцип Олли. 5.2. Динамические характеристики популяций. 5.1. Принцип Олли. Принцип агрегации особей В. Олли - в экологии – закон (1931), согласно которому скопление особей, усиливает конкуренцию между ними за пищевые ресурсы и жизненное пространство, но приводит к повышенной способности группы к выживанию. Следовательно, как «перенаселенность» (повышенная агрегация особей), так и «недонаселенность» (отсутствие агрегации) могут служить лимитирующими факторами. 5.2. Динамические характеристики популяций. Динамические показатели характеризуют процессы, протекающие в популяции за какой-то промежуток времени. Основными динамическими показателями популяций являются: рождаемость, смертность, скорость роста популяций. Рождаемость – это число особей, рождающихся в популяции за единицу времени. Различают абсолютную и относительную рождаемость. Общая (абсолютная) рождаемость – это абсолютное число новых особей: например, в популяции в течение года родилось 156 новых особей. Относительная (удельная) рождаемость – отношение числа новых особей к числу имевшихся особей; относительная рождаемость может рассчитываться или на одну особь, или на 1000 особей. Например, в популяции в начале года было 10 000 особей, а в течение года родилось 156 новых особей; тогда относительная рождаемость равна 156 : 10000 = 0,0156 на одну особь, или 0,0156 · 1000 = 15,6 на тысячу особей. Численность популяции может увеличиваться не только за счет рождаемости, но и за счет иммиграции особей из других популяций. Существуют зависимые и полузависимые популяции, которые поддерживают и увеличивают свою численность именно за счет иммиграции. Рождаемость, или скорость рождаемости, выражают отношением: В = Nn/t, где Nn — число особей, родившихся за некоторый промежуток времени t. Но для сравнения рождаемости в различных популяциях пользуются величиной удельной рождаемости: отношением скорости рождаемости к исходной численности (N): b = Nn/Nt. За бесконечно малый промежуток времени (t —> 0) мы получим мгновенную удельную рождаемость, которую обозначают латинской буквой «b». Эта величина имеет размерность «единица времени -1» и зависит от интенсивности размножения особей: для бактерий — час, для фитопланктона — сутки, для насекомых — неделя или месяц, для крупных млекопитающих — год. Причиной массовых вспышек рождаемости в популяциях являются, как правило, погодные факторы и деятельность человека. Часто популяции при помощи механизма обратных связей способны регулировать свою численность с тем, чтобы она не превысила предел, за которым наступит катастрофа. Так, гусеницы некоторых бабочек массово гибнут, если выходят из яиц до окончания холодов (когда они погибают от переохлаждения) либо через длительное время после распускания листьев (тогда они погибают от голода). Если гусеницы появляются вовремя, то популяции грозит перенаселение; в это время в действие вступает второй лимитирующий фактор – например, хищники или паразиты. В данном случае действие регулирующего фактора зависит от плотности популяции. Смертность – это число особей, погибших в популяции в единицу времени. Различают абсолютную смертность (количество погибших особей за единицу времени) и относительную (удельную) смертность (количество погибших особей за единицу времени в расчете на одну особь или на 1000 особей). В отличие от рождаемости смертность наблюдается постоянно. Характер смертности описывается таблицами и кривыми выживаемости, которые показывают, какая часть новорожденных особей дожила до определенного возраста. Убыль и прибыль организмов в популяции зависит не только от рождаемости и смертности, но и от скорости их миграции и эмиграции – количества особей, прибывших и убывших в популяции в единицу времени. Смертность — величина обратная рождаемости, но измеряется в тех же величинах и вычисляется по аналогичной формуле. Если принять, что Nm — число погибших особей (независимо от причины) за время t, то удельная смертность: Nm/Nt, а при t 0 имеем мгновенную удельную смертность, которую обозначают буквой «d». Величины рождаемости и смертности по определению могут иметь положительное значение, либо равное нулю. Выживаемость есть параметр, обратный смертности. Если смертность на N особей обозначить через М, то выживаемость определяется разностью N - M. Большой интерес представляют кривые выживания особей популяции в зависимости от их возраста.
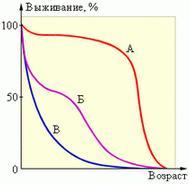
Учёным, исследующим динамику популяций, важно бывает знать не только общую смертность в популяции, но и количество особей, погибающих до достижения половой зрелости. Условием стабильности популяции является тот факт, что до половой зрелости доживают два потомка от каждой пары. Динамику смертности популяции в зависимости от возраста отображают на кривых выживания. Существуют три основных типа этих кривых: когда главным фактором смертности является естественное старение особей (на рисунке эта кривая обозначена буквой А), когда в популяции высока ранняя смертность (Б) и, наконец, когда смертность постоянна в течении всей жизни организмов (В). Кривые выживания конкретных видов могут занимать промежуточные положения между этими крайними формами.
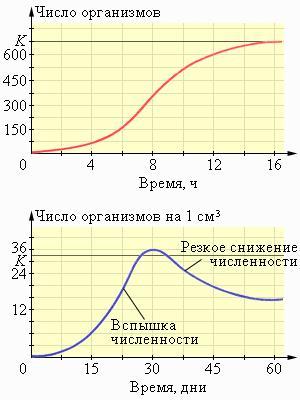
Кривая а на рисунке соответствует почти идеальной ситуации, при которой главная причина гибели особи — старение. Такой график можно получить также для однолетней культуры, например, пшеницы — почти все растения доживают до осени, а потом одновременно плодоносят и отмирают. Кривая б соответствует ситуации, когда смертность особенно высока в ранний период жизни особей. Это наблюдается, например, у горных баранов и людей в слаборазвитых странах с плохим медицинским обслуживанием и дефицитом питания. Кривая в отражает не зависящую от возраста смертность, равную 50% для любого временного интервала. В этом случае гибель особей происходит в основном случайно еше до того, как станут очевидными признаки старения и не связана с внутренней устойчивостью особей к внешним факторам. Сходную кривую можно получить, например, для пресноводной гидры, молодь которой не подвержена особому риску по сравнению с другими возрастными группами. Однако у большей части беспозвоночных и растений кривая выживания, хотя и соответствует в целом типу в, требует поправки на повышенную уязвимость неполовозрелых стадий, т. е. правая часть графика должна идти вниз более круто. По кривым выживания можно определить смертность разных возрастных групп, т. е. определить, какие стадии их жизни наиболее уязвимы. Выявив факторы, вызывающие гибель на этих стадиях, нетрудно понять, как регулируется численность популяции. Если рождаемость в изолированной популяции превышает смертность, то популяция растёт. В начале роста кривая роста популяции – это экспонента. Однако рано или поздно питательные запасы в окружающей среде исчерпываются. В одних популяциях уменьшение количества питательных веществ начинает воздействовать на динамику роста загодя, и кривая роста приобретает S-образную форму. В других популяциях бесконтрольный рост численности заходит слишком далеко, после чего происходит катастрофический «обвал» численности, связанный с истощением ресурсов (J-образная форма). Рост популяций на первой (экспоненциальной) фазе можно приближённо описать дифференциальным уравнением
где N (t) – количество особей в зависимости от времени, b – рождаемость, d – смертность, а r = b – d – врождённая скорость роста численности популяции. Дифференциальным уравнением можно описать и S-образную кривую:
Здесь K – это максимальный размер стабильной популяции, которая может существовать неопределённо долго, если не изменятся внешние условия. Заметим, что дифференциальные уравнения роста – приблизительные уравнения, так как они не учитывают различия между особями, сезонные колебания количества пищи и способности к размножению и т. п. Виды, которые быстро размножаются со скоростью, не зависящей от плотности вида, называют r-стратегами. Размеры r-популяций не стабилизируются и в течение некоторого времени могут превышать поддерживающую ёмкость среды. Как правило, r-стратеги имеют небольшие размеры и малую продолжительность жизни. Среди них много микроорганизмов, мелких насекомых и однолетних растений. Обычно r-стратеги быстрее заселяют новые местообитания, однако через некоторое время их вытесняют более конкурентоспособные K-стратеги. K-стратеги размножаются относительно медленно. Скорость их размножения зависит от плотности популяции. Численность популяций K-стратегов через некоторое время стабилизируется на определенном значении. Среди K-стратегов обычно встречаются крупные и долгоживущие виды: деревья, крупные птицы и звери, человек.
Биотический потенциал и сопротивление среды. Сохранение или рост численности зависит не только от скорости размножения (число новорожденных, отложенных яиц, произведённых семян или спор в единицу времени). Не менее важно и пополнение взрослого состава популяции за счёт потомства. Высокая скорость размножения при низких темпах пополнения не может существенно увеличить её численность. Например - рыбы вымётывают тысячи или миллионы икринок, но лишь ничтожно малая часть выживает и превращается во взрослое животное. Растения рассеивают огромное количество семян. И напротив, размер популяции может расти за счёт увеличения темпов пополнения при малой скорости размножения. Это относится к людям (рождаемость низкая, но детская смертность низкая, поэтому практически все дети доживают до взрослого возраста). Другим важным фактором, ведущим к росту популяции, относится способность животных мигрировать, а семян рассеиваться на новых территориях, приспосабливаться к новым местам обитания и заселять их, наличие защищённых механизмов и устойчивость к неблагоприятным условиям среды и болезням. Биотический потенциал - это совокупность факторов, способствующих увеличению численности вида. У разных видов составляющие биотического потенциала неодинаковы, но они имеют одно общее свойство - стремительное увеличение численности при благоприятных условиях среды. В естественных условиях такое наблюдается редко. Вероятность того, что все условия окажутся благоприятными очень низка. Обычно один или несколько факторов (t, влажность, солёность, хищники, паразиты, нехватка пищи) становятся лимитирующими. Сочетание всех таких «ограничителей» называют сопротивлением среды. Сильнее всего они действуют на молодых особей, а это снижает темпы пополнения. При более суровых условиях гибнет часть взрослых особей. Следовательно: рост, снижение и постоянство популяции зависит от соотношения между биотическим потенциалом и сопротивлением среды. Принцип изменения популяции: это результат нарушения равновесия между биотическим потенциалом и сопротивлением окружающей её среды. Подобное равновесие является динамическим, т.е. непрерывно регулирующимся, т.к. факторы сопротивления среды редко подолгу остаются неизменными. Например: в один год численность популяции снизилась из-за засухи, а в следующий год полностью восстановилась при обильных дождях. Подобные колебания продолжаются неопределённо долго. Равновесие - понятие относительное. Иногда амплитуда отклонений мала, иногда значительна, но пока сократившаяся популяция способна восстановить прежнюю численность, она существует. Равновесие в природных системах зависит от плотности популяции, т.е. числа особей на единицу площади. Если плотность популяции растёт - сопротивление среды увеличивается, в связи, с чем увеличивается смертность и рост численности прекращается. И, наоборот, с уменьшением плотности популяции - сопротивление среды ослабевает и восстанавливается прежняя численность. Воздействие человека на природу часто приводит к вымиранию популяции. Разрушение экосистем, загрязнение окружающей среды одинаково влияют на популяции, как с низкой, так и высокой плотностью. Кроме этого, биотический потенциал зависит от критической численности популяции. Если численность популяции (оленей, птиц или рыб) падает ниже этой величины, гарантирующей воспроизводство, биотический потенциал стремится к нулю и вымирание неизбежно. Существование может быть поставлено под угрозу, даже когда множество представителей вида живы, но живут в домашних условиях, т.е. изолированы друг от друга (попугаи).
|