

Типи яйцеклітин
Залежно від кількості жовтка (lecithos) в цитоплазмі яйцеклітин різних видів тварин їх підрозділяють на наступні типи: - безжовткові (алецитальні), - маложовткові (оліголецитальні), - середньожовткові (мезолецитальні ) і - багатожовткові (полілецитальні).
По характеру поділу жовтка в цитоплазмі яйцеклітин вони діляться на:
• центролецитальні, коли жовток займає центральну частину цитоплазми;
• ізолецитальні ,в яких жовток рівномірно розподілений по цитоплазмі . До цієї групи відносяться оліголецитальні яйцеклітини. Серед них розрізняють первинно ізолецітальние (у ланцетника) і вдруге ізолецітальние (у ссавців ) ;
• телолецитальні, якщо жовток концентрується поблизу вегетативного полюса. До цієї групи входять мезолецитальні - із середнім вмістом жовтка (у амфібій) і полілецитальні яйцеклітини , перевантажені жовтком від якого вільна лише невелика частина анімального полюса (уптахів)
Яйцеклітина у всіх тварин має полярність. Два її протилежні полюси називають анімальним і вегетативним. Полярність яйця часто виявляється в розташуванні цитоплазматичних включень. У багатьох яйцях жовток розташований нерівномірно, його кількість зростає від анімального полюса до вегетативного. Тип дроблення заплідненої яйцеклітини залежить від кількості жовтка і характеру його поділу в цитоплазмі яйця. Розрізняють повне дроблення, коли дробиться вся яйцеклітина, і неповне, коли дробиться тільки її частина. Це обумовлено тим, що жовток перешкоджає утворенню перетяжки при розподілі тіла клітини. Повне дроблення, у свою чергу, буває рівномірним, якщо в результаті поділу клітини, що утворюються, приблизно однакові за величиною, і нерівномірним, якщо вони відрізняються за своїми розмірами.
Ембріональний розвиток ланцетника
Ембріональний розвиток складається з наступних основних етапів: 1) дроблення, в результаті якого утворюється багатоклітинний зародок; 2) гаструляції, в процесі якої виникають перші ембріональні тканини — ектодерма і ентодерма, а зародок стає двошаровим; 3) первинного органогенезу — утворення комплексу осьових органів зародка — нервової трубки, хорди, кишкової трубки.
Яйцеклітина у всіх тварин має полярність. Два її протилежні полюси називають анімальним і вегетативним. Полярність яйця часто виявляється в розташуванні цитоплазматичних включень. У багатьох яйцях жовток розташований нерівномірно, його кількість зростає від анімального полюса до вегетативного. Тип дроблення заплідненої яйцеклітини залежить від кількості жовтка і характеру його поділу в цитоплазмі яйця. Розрізняють повне дроблення, коли дробиться вся яйцеклітина, і неповне, коли дробиться тільки її частина. Це обумовлено тим, що жовток перешкоджає утворенню перетяжки при розподілі тіла клітини. Повне дроблення, у свою чергу, буває рівномірним, якщо в результаті поділу клітини, що утворюються, приблизно однакові за величиною, і нерівномірним, якщо вони відрізняються за своїми розмірами.
В яйцеклітині ланцетника жовтка мало, і він рівномірно розподілений у цитоплазмі, тому дроблення заплідненої яйцеклітини повне і рівномірне. Перша борозна проходить у меридіональній площині по напряму від анімального полюса до вегетативного, розділяючи зиготу на дві клітини рівної величини. У результаті дроблення зиготи клітини, що утворюються, називаються бластомерами (від грец. blastos — зародок і meros — частина). Друга борозна також проходить у меридіональній площині, але перпендикулярно першій. Виходять чотири клітини. Третя борозна дроблення — широтна, вона проходить дещо вище від екватора і відразу розділяє чотири бластомери на вісім клітин. Далі правильно чергують меридіональні та широтні борозни. У міру збільшення числа клітин поділ їх стає асинхронним. Бластомери все далі розходяться від центру зародка, формуючи порожнину. Врешті-решт зародок приймає форму пухирця із стіною, утвореною одним шаром клітин, тісно прилягаючих один до одного.
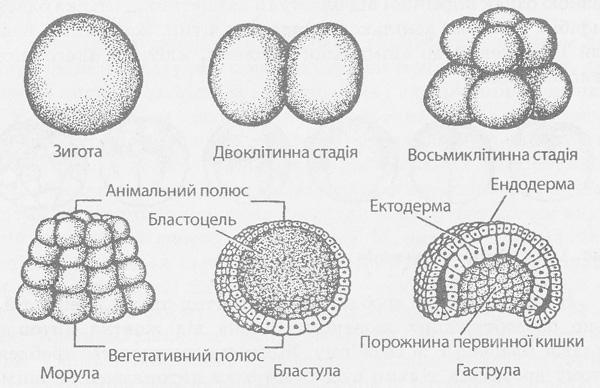
Рис. 146. Дроблення та утворення бластули у ланцетника.
Внутрішня порожнина зародка, що спочатку сполучається із зовнішнім середовищем через проміжки між бластомерами, в результаті їх щільного стулення стає абсолютно ізольованою. Ця порожнина носить назву первинної порожнини тіла, а сам зародок в кінці періоду дроблення — бластули. Загальний об'єм бластомерів, що з'явилися в результаті дроблення, не перевищує об'єму зиготи. Таким чином, мітотичний поділ зиготи і бластомерів не супроводиться зростанням дочірніх клітин, що утворилися, до розмірів материнської, і розміри бластомерів у результаті послідовних розподілів прогресивно зменшуються. Ця особливість мітотичного поділу бластомерів спостерігається при розвитку запліднених яйцеклітин всіх типів. Під час дроблення цитоплазма не переміщається.
Наступний етап ембріонального розвитку — гаструляція, або утворення двошарового зародка. Процес гаструляції здійснюється різними способами і залежить від будови бластули, тобто кінець кінцем від кількості жовтка в яйцеклітині.
У ланцетника двошаровий зародок утворюється шляхом вигинання стінки бластули в порожнину бластоцілі. Виникають два зародкові листки: зовнішній — ектодерма і внутрішній — ентодерма
Диференціювання — це процес появи і наростання морфологічних і функціональних відмінностей між окремими клітинами і частинами зародка
При подальшому диференціюванні клітин, що входять до складу зародкових листків, з ектодерми утворюється нервова система, органи чуття, епітелій шкіри, емаль зубів; з ентодерми — епітелій середньої кишки, травні залози, епітелій зябер і легенів; з мезодерми — м'язова, сполучна, хрящова і кісткова тканина, нирки, статеві залози, кровоносна система.
У різних видів тварин одні й ті ж зародкові листки дають одні й ті ж органи і тканини. Це значить, що зародкові листки гомологічні. Гомологія зародкових листків переважної більшості тварин — один з доказів єдності тваринного світу.
Після завершення гаструляції у зародка утворюється комплекс осьових органів: нервова трубка, хорда, кишкова трубка. У ланцетника осьові органи формуються таким чином. Ектодерма, що знаходиться на спинній стороні зародка, прогинається по середній лінії, перетворюючись на жолоб, а ектодерма, розташована справа і зліва від жолоба, починає наростати на його краї. Жолоб — зачаток нервової системи — занурюється під ектодерму, і краї його стуляються. Утворюється нервова трубка. Вся інша ектодерма є зачатком шкірного епітелію.
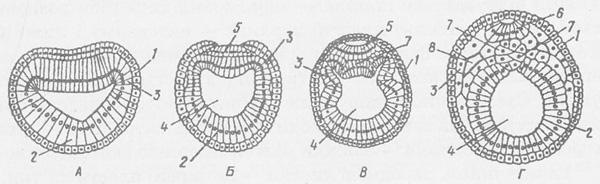
Рис. 148. Гаструляція та утворення комплексу осьових органів у ланцетника (поперечний розріз). А, Б, В — стадії гаструли; Г — формування зачатків осьових органів — нервової трубки, хорди, кишкової трубки: 1 — ектодерма, 2 — ентодерма, 3 — зачаток мезодерми, 4 — порожнина кишки, 5 — нервова пластинка, 6 — нервова трубка, 7 — хорда, 8 — порожнина тіла.
Спинна частина ентодерми, розташована безпосередньо під нервовим зачатком, відособлюється від іншої ентодерми і згортається в щільний тяж — хорду. З частини ентодерми, що залишилася, розвиваються мезодерма та епітелій кишечнику. Подальше диференціювання клітин зародка приводить до виникнення численних похідних зародкових листків — органів і тканин.
Клонування, в біології - це метод одержання декількох ідентичних організмів шляхом безстатевого (у тому числі вегетативного) розмноження. Клонування" - отримання нащадків, що є точною генетичною копією організму.Сукупність таких нащадків-копій, що походять від одного організму, називають клоном.Організми в межах кожного клону характеризуються однаковою фенотипической однорідністю та ідентичним генотипом.
Перші успішні досліди з клонування тварин були проведені в середині 1970-х років англійським ембріологом Дж. Гордоном (J. Gordon) в експериментах на амфібія, коли заміна ядра яйцеклітини на ядро з соматичної клітини дорослої жаби призвела до появи пуголовка.Це показало, що техніка трансплантації ядер із соматичних клітин дорослих організмів у енуклеірованние ооцити дозволяє отримувати генетичні копії організму, що став донором ядер диференційованого клітин.Результат експерименту став підставою для висновку про оборотності ембріональної диференціювання геному принаймні у земноводних [18] У своєму експерименті Кемпбелл і його колеги витягли з ембріона вівці на ранній стадії розвитку (на стадії ембріонального диска) клітину і виростили культуру клітин, тобто домоглися того, що клітина розмножилася в штучному живильному середовищі.Отримані генетично ідентичні клітини (клітинна лінія) зберегли тотіпонентность.Потім вчені взяли яйцеклітину вівці-реципієнта, ретельно видалили з неї весь хромосомний матеріал і домоглися її злиття з тотипотентності кліткою з культури.Отримані синтетичні ембріони вирощували до стадії морули-бластули, а потім імплантували в матку вівці.У результаті вдалося виростити декількох нормальних ягнят, які були генетично ідентичні.
Клонування тварин можливе за допомогою експериментальних маніпуляцій з яйцеклітинами (ооцитами) і ядрами соматичних клітин тварин in vitro та in vivo подібно до того, як в природі з'являються однояйцеві близнюки.Клонування тварин досягається в результаті перенесення ядра з диференційованої клітини в незапліднену яйцеклітину, у якої видалене власне ядро (енуклеірованних яйцеклітина) з подальшою пересадкою реконструйованої яйцеклітини в яйцепровід названої матері.
У 1996 році з'явилися їх публікації по успішному народженню ягнят у результаті трансплантації ядер, отриманих з фібробластів плоду вівці, в енуклеірованние ооцити.[2] В остаточному вигляді проблема клонування тварин була вирішена групою Вільмута в 1997, коли народилася вівця на прізвисько Доллі - перше ссавець, отримане з ядра соматичної клітини дорослої: власне ядро ооцита було замінено на ядро клітини з культури епітеліальних клітин молочної залози дорослої лактуючої вівці .[3] Надалі були проведені успішні експерименти з клонування різних ссавців з використанням ядер, взятих з дорослих соматичних клітин тварин (миша, коза, свиня, корова), а також взятих у мертвих, заморожених [4] на кілька років, тварин.
Методи клонування тварин
Розроблені методи клонування тварин поки ще далеко не досконалі.У процесі експериментів спостерігається висока смертність плодів і новонароджених.Ще не ясні багато теоретичні питання клонування тварин з окремої соматичної клітини.
Проте успіх, досягнутий в клонуванні вівці і мавп, показав теоретичну можливість створення генетичних копій також людини з окремої клітини, взятої з якого-небудь його органу.Багато вчених з ентузіазмом сприйняли ідею клонування людини.
1 Методи трансплантації ядер
У нашій країні Б.В.Конюховим і Є.С.Платоновим в 1985 р. був розроблений метод менш травматичного перенесення ядер методом мікроманіпуляції.Він протікає в два етапи: спочатку тонкою мікропіпеткою проколюють зони пеллюціда і плазматичної мембрани і витягають пронуклеус, а потім інший піпеткою, більшого діаметру (12 мкм) у той же отвір вводять диплоидное ядро донора.У цьому випадку менше травмується цитоплазма зиготи і транспортується ядро донора.
2 Метод генетичного перепрограмірованія клітин шкіри
Розроблено новий метод клонування - менш трудомісткий, ніж спосіб, завдяки якому була створена овечка Доллі.У зв'язку з цим виникли побоювання, що одного разу він буде використаний для обробки ембріонів людини, щоб формувати дітей "на замовлення".
Вчені, завдяки цьому методу отримали мишенят з клітин шкіри дорослих особин, виявили: така технологія набагато ефективніша, ніж спосіб створення Доллі, а побічних ефектів у неї менше - отже, вона краще підходить для використання стосовно людини.