

Лекция 3. Биология
.docxЛекция 3. ГЕНЕТИЧЕСКИЙ КОД. БИОСИНТЕЗ БЕЛКА. ТРАНСЛЯЦИЯ.
Впервые 2 английских ученых Бидл и Татум доказали, что на основе молекулы ДНК происходит синтез полипептидной белковой молекулы и вывели формулу 1 ген = 1 полипептидная молекула. Позднее это оказалось не совсем так, потому что много генов определяют не белок, а, к примеру, тРНК.
Закономерно встал вопрос: как на основе ДНК информация идет на белковые молекулы? Впервые американец русского происхождения Гамов вывел закономерность: 4 нуклеотида определяют 20 аминокислот. Если 1 нуклеотид будет кодировать 1 аминокислоту (код моноплетен), то можно зашифровать только 4 аминокислоты (4^1=4). Если 2 нуклеотида определяют 1 аминокислоту (код диплетен), то можно зашифровать 16 аминокислот (4^2=16). Если код триплетен, то можно зашифровать 64 аминокислоты (4^3=64).
Впервые американский ученый Нирренберг синтезировал искусственную нуклеиновую кислоту. Она состояла только из У, т. н. полиуридиновая кислота. И в искусственных условиях ему удалось на ее основе получить полипептидную молекулу, которая состояла только из фенилаланина (1 аминокислота). Было принято решение, что УУУ определяет в белке аминокислоту фенилаланин. На ней же удалось получить все 64 варианта кодонов и определить, какую аминокислоту эти кодоны определяют. Были ведены общие свойства генетического кода:
1) генетический код универсален, практически одинаков для всех живых организмов;
2) код триплетен (1 аминокислота = 3 нуклеотида);
3) код не перекрывается (коды идут линейно друг за другом, и нуклеотид из одного кода используется другим);
4) нет разделительных знаков между триплетами;
5) код отличается коллинеарностью (совпадением кодонов иРНК и аминокислот в белке);
6) код является вырожденным, т. е. избыточным, поскольку 64 кодона определяют 20 аминокислот, то некоторые аминокислоты (большинство) имеют 2-3 (иногда даже 4) кодона;
7) из 3 нуклеотидов в кодоне главными являются первые 2, третий может варьировать;
8) в среднем, каждая аминокислота кодируется 3 триплетами;
9) частота использования различных кодонов может быть видоспецифичной, т. е. по частоте кодонов виды могут различаться;
10) инициирование кодонами, т. е. кодонами в начале молекулы ДНК являются УАГ и ГУГ. ОНи указывают начало синтеза полипептидной цепи, а перед ними еще стоит регуляторная часть, с которой полипептидная цепь не синтезируется.
Первая аминокислота - метиамин или формилметиамин. В конце - терминирующие нонсенс-кодоны, они не определяют никакую аминокислоту, а определяют точку конца информации (УАА, УГА, УАГ). За последним стоп-кодоном на 3'-конце иРНК располагаются некодирующие белок последовательности - трейлеры. В состав начального лидерного участка входит последовательность АГГ АГГ, она служит сигналом начала трансляции с иРНК. Ее стали называть последовательностью Шайн-Далгарно. Эта последовательность располагается на 5-7 нуклеотидов левее стартового кодона, она обеспечивает присоединение иРНК к рибосоме.
Имеются существенные отличия РНК прокариот и эукариот. У эукариот РНК более стабильна, это связано с процессами модификации незрелой иРНК. Сразу же после синтеза иРНК на 5'-головном конце образуется т. н. колпачок - кэп. Кэп представляет из себя гуанозин, у которого в 7 положении имеется метильная группа - 7-метилгуанозин. Кроме того, метилируется рибоза. Это формирует 2-й или даже 3-й кэп. Кроме того, к 3'-концу хвостовой части ферментами присоединяется до 200 адениловых остатков. Этот адениловый хвост не кодирует белок. После этого РНК покидает ядро, при этом число адениловых остатков может сокращаться. Установлено, что такие модификации (кэп и хвост) существенно стабилизируют молекулу, и уже клеточные нуклеазы не разрушают эту РНК. У прокариот это отсутствует и после 1 цикла синтеза белка РНК разрушается нуклеазами.
Когда ученые впервые определили, что количество нуклеотидов в гене и количество нуклеотидов в РНК не совпадают, было определено, что иРНК во много раз короче, чем ген. Было установлено, что в гене имеются незначимые участки, их назвали "интроны"; и значимые участки, которые несу информацию о белке, их назвали "экзоны". Кроме этого, в гене есть спейсеры – регуляторные участки. Оказалось, что из незрелой РНК происходит вырезание интронов и спейсеров, это явление было названо сплайсингом, а все преобразование незрелой РНК в зрелую, образование кэпов, аденилового хвоста, вырезание интронов и сшивание экзонов называется процессингом. Вырезание идет по правилу Шамбона, согласно которому интроны всегда начинаются ГУ и заканчиваются АГ.
МЕХАНИЗМ ТРАНСЛЯЦИИ.
Сразу после опубликования работы Уотсона и Крика о структуре молекулы ДНК были высказаны разные идеи о матричном синтезе белка, они предполагали взаимодействие РНК и белковой молекулы, но эта идея была отвергнута. И впервые Френсис Крик пришел к выводу, что прямого контакта между РНК и синтезируемой полипептидной молекулой нет. Он выдвинул адаптерную гипотезу, т. е. должны быть адаптерные молекулы, которые с одной стороны должны соединяться с аминокислотой, а с другой - узнавать кодоны на иРНК.
Впервые американский ученый Хогленд обнаружил, что в клетке аминокислоты соединены с низкомолекулярной РНК, при этом было установлено, что это соединение обусловлено ферментом аминоацил-тРНК-синтетазами (АРСазы).
Согласно адаптерной гипотезе, функции адаптера осуществляются в 2 этапа:
1) трансляция I
а) активация аминокислоты;
б)аминоацилирование - соединение тРНК с аминокислотой при посредстве АРСазы
2) трансляция II
а) перенос аминокислоты с помощью тРНК к кодонам иРНК
б) опознание кодона тРНК
тРНК для этого должна обладать следующими сайтами:
1. сайтом, специфичным АРСазе
2. сайтом связывания с кодоном антикодона тРНК
3. сайтом связывания с рибосомой
4. сайтом взаимодействия с различными факторами трансляции II
Таким образом, число тРНК и АРСаз в клетке должно быть не менее 20. Нет прямой связи между тРНК и аминокислотой. Свободная тРНК и аминокислота сцеплены с помощью АРСазы.
тРНК по своему строению напоминает трилистник клевера, при этом у нее есть акцепторный стебель, здесь на 3'-конце имеется кодон А АЦЦ, справа ТYC [ти пси цэ], дигидроуридиновая петля, антикодон внизу.
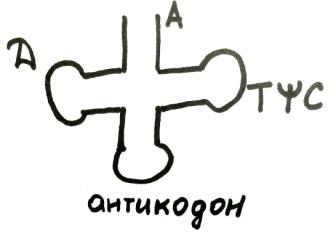
Впервые тРНК аланиновой аминокислоты была расшифрована Робертом Холли, а затем были расшифрованы и другие тРНК. В настоящее время известно более 300.
тРНК имеет 4 двуцепочечных и 5 одноцепочечных участков, в ней имеются т. н. минорные азотистые основания (дигидроуридин, псевдоуридин, метилинозин, метилурацил). Они не могут образовать двуцепочечные участки и содержатся в одноцепочечных.
Антикодирующая петля состоит из 7 нуклеотидов, с помощью которых тРНК взаимодействует с кодоном иРНК. Дигидроуридиновая петля способствует формированию третичной структуы. Двуцепочечные структуры делят 2 витка. Как ДНК и тРНК имеет Г-образную форму. Найдено много тРНК, несущих 1 аминокислоту, но иеющих разные антикодоны. Это изоакцепторные тРНК. Физическая конфигурация АРСаз повторяет конфигурацию родственной тРНК.
Антикодон тРНК узнает кодон иРНК путем спаривания азотистых оснований, например, дрожжевой аланин тРНК может связываться с 3 кодонами (ГЦУ, ГЦЦ и ГЦА).
Впервые Френсис Крик предположил, что основными являются только первые 2 нуклеотида, а относительно третьего возможна свобода.
Кодоны, различающиеся по одному из первых двух оснований должны узнаваться различными тРНК. Например, УУА и ЦУА оба кодируют лейцин, но считываются разными тРНК. Первое основание антикодона определяет, считывает ли данная тРНК один, два или три типа кодонов. Если это Ц или А, узнается только один кодон, если У и Г, то 2 кодона. Если необычный инозин, то 3 кодона.
Т. о., причина вырожденности кода - неоднозначность спаривания третьего кодона.
Весь процесс биосинтеза белка происходит на рибосомах. Рибосома - это сложная структура диаметром 2 нм, коэффициент седиминтации осаждения 50s, для маленькой - 30s, у прокариот 50s состоит из 34 видов белков, двух молекул РНК, одна 23s, а другая 25s.Маленькая частица имеет 21 белок и 16 РНК. У эукариот рибосомы крупнее. Крупная - 60S, имеет з РНК, 28, 7 и 5S, а малая (40S) имеет 18S-РНК. Все эти компоненты самособираемы. Если биосинтез не происходит, то большая и малая субчастицы плавают в цитоплазме. Отдельно при реконструкции 30s-субчастицы были сделаны выводы:
1) 16s-РНК необходима для сборки и функционирования рибосомы
2) она видоспецифична
3) из 80 белков 6 строго необходимы для сборки.
Опыты Говарда-Динциса показали, что считываются от аминного конца к карбонильному, при этом на иРНК через каждые 8 нуклеотидов сидит новая рибосома, и те рибосомы, которые близко находятся к РНК-полимеразе синтезируют крупные полипептидные молекулы, а которые на конце - малые, т. е. рибосомы ближе к 5'-концу несут самые короткие полипептидные цепи, а к 3'-концу - самые длинные.
После биосинтеза субчастицы распадаются. Первый трансляционный кодон всегда находится на расстоянии не менее 25 нуклеотидов от 5'-конца.