

Лабораторное занятие 1.
Устройство микроскопа и правила работы с ним. Строение растительной клетки.
Цель: Познакомиться с устройством микроскопа и правилами работы с ним. Познакомиться со строением растительной клетки.
Какие системы входят в состав микроскопа?
Микроскоп - это оптический прибор, позволяющий получить обратное изображение изучаемого объекта и рассмотреть мелкие детали его строения, размеры которых лежат за пределами разрешающей способности глаза.
Разрешающая способность микроскопа дает раздельное изображение двух близких друг другу линий. Невооруженный человеческий глаз имеет разрешающую способность около 1/10 мм или 100 мкм. Лучший световой микроскоп примерно в 500 раз улучшает возможность человеческого глаза, т. е. его разрешающая способность составляет около 0,2 мкм или 200 нм.
Разрешающая способность и увеличение не одно и тоже. Можно получить большое увеличение, но не улучшить его разрешение.
Различают полезное и неполезное увеличения. Под полезным понимают такое увеличение наблюдаемого объекта, при котором можно выявить новые детали его строения. Неполезное - это увеличение, при котором, увеличивая объект в сотни и более раз, нельзя обнаружить новых деталей строения.
В учебных лабораториях обычно используют световые микроскопы, на которых микропрепараты рассматриваются с использованием естественного или искусственного света. Наиболее распространены световые биологические микроскопы: БИОЛАМ, МИКМЕД, МБР, МБИ и МБС. Они дают увеличение в пределах от 56 до 1350 раз.
Стереомикроскоп (МБС) (рис. 2)обеспечивает подлинно объемное восприятие микрообъекта и увеличивает от 3,5 до 88 раз. В микроскопе выделяют две системы: оптическую и механическую (рис. 1). К оптической системе относят объективы, окуляры и осветительную систему (конденсор с диафрагмой и светофильтром, зеркало или электроосветитель).
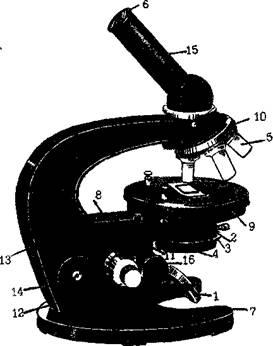 Рис.
1
Рис.
1
1- зеркало, 2- конденсор, 3- ирисовая диафрагма, 4- съемный светофильтр, 5- объектив, 6- окуляр, 7- подставка, 8- коробка с микрометренным механизмом, 9- предметный столик, 10 револьвер, 11 кронштейн конденсора, 12-икрометренный винт, 13-тубусодержатель, 14- винт грубой наводки, 15-тубус, 16 - рукоятка конденсора
Объектив - определяет полезное увеличение объекта. Объектив состоит из нескольких линз. Увеличение объектива обозначено на нем цифрами.
В учебных целях используют обычно объективы х8 и х40.
Окуляр состоит из 2-3 линз. Увеличение окуляров обозначено на них цифрами: х7, х10, х15.
Окуляры не выявляют новых деталей строения и в этом отношении их увеличение бесполезно.
Для определения общего увеличения микроскопа следует умножить увеличение объектива на увеличение окуляра. В случае использования бинокулярной или тринокулярной насадки, в данное уравнение нужно добавить собственное увеличение насадки. Для бинокулярной насадки АУ-12 ЛОМО, увеличение которой составляет 1,5Х. В таких насадках как АУ-26 или МФН-11 увеличение можно менять, собственное сменное увеличение насадки АУ-26 -- 1,1x; 1,6x и 2,5x
Осветительное устройство состоит из зеркала или электроосветителя, конденсора с ирисовой диафрагмой и светофильтром, расположенных под предметным столиком. Они предназначены для освещения объекта пучком света.
Механическая система микроскопа состоит из подставки, коробки с микрометренным механизмом и микрометренным винтом, тубусодержателя, винта грубой наводки, кронштейна конденсора, винта перемещения конденсора, револьвера, предметного столика.
Каковы основные правила работы с микроскопом?
При работе с микроскопом необходимо соблюдать операции в следующем порядке:
1. Работать с микроскопом следует сидя;
2. Микроскоп осмотреть, вытереть от пыли мягкой салфеткой объективы, окуляр, зеркало;
3. Микроскоп установить перед собой, немного слева на 2-3 см от края стола. Во время работы его не сдвигать;
4. Открыть полностью диафрагму, поднять конденсор в крайнее верхнее положение;
5. Работу с микроскопом всегда начинать с малого увеличения;
6. Опустить объектив 8 х в рабочее положение, т.е. на расстояние 1см от предметного стекла;
7. Глядя одним глазом в окуляр и пользуясь зеркалом с вогнутой стороной, направить свет от окна в объектив, а затем максимально и равномерно осветить поле зрения;
8. Положить микропрепарат на предметный столик так, чтобы изучаемый объект находился под объективом. Глядя сбоку, опускать объектив при помощи макровинта до тех пор, пока расстояние между нижней линзой объектива и микропрепаратом не станет 4-5 мм ;
9. Смотреть одним глазом в окуляр и вращать винт грубой наводки на себя, плавно поднимая объектив до положения, при котором хорошо будет видно изображение объекта. Нельзя смотреть в окуляр и опускать объектив. Фронтальная линза может раздавить покровное стекло, и на ней появятся царапины;
10. Передвигая препарат рукой, найти нужное место, расположить его в центре поля зрения микроскопа;
11. Если изображение не появилось, то надо повторить все операции пунктов 6, 7, 8, 9;
12. Для изучения объекта при большом увеличении сначала нужно поставить выбранный участок в центр поля зрения микроскопа при малом увеличении. Затем поменять объектив на 40 х, поворачивая револьвер, так чтобы он занял рабочее положение. При помощи микрометренного винта добиться хорошего изображения объекта. На коробке микрометренного механизма имеются две риски, а на микрометренном винте - точка, которая должна все время находится между рисками. Если она выходит за их пределы, ее необходимо возвратить в нормальное положение. При несоблюдении этого правила, микрометренный винт может перестать действовать;
13. По окончании работы с большим увеличением, установить малое увеличение, поднять объектив, снять с рабочего столика препарат, протереть чистой салфеткой все части микроскопа, накрыть его полиэтиленовым пакетом и поставить в шкаф.
3. Какие основные операции нужно проделать, чтобы привести микроскоп в рабочее состояние?
а) установить объектив малого увеличения на расстоянии 1-1,5 см от предметного столика;
б) поднять конденсор до предела;
в) смотря в окуляр одним глазом, не закрывая другого, равномерно и интенсивно осветить зеркалом поле зрения.
4. Основные части растительной клетки, их характеристика.
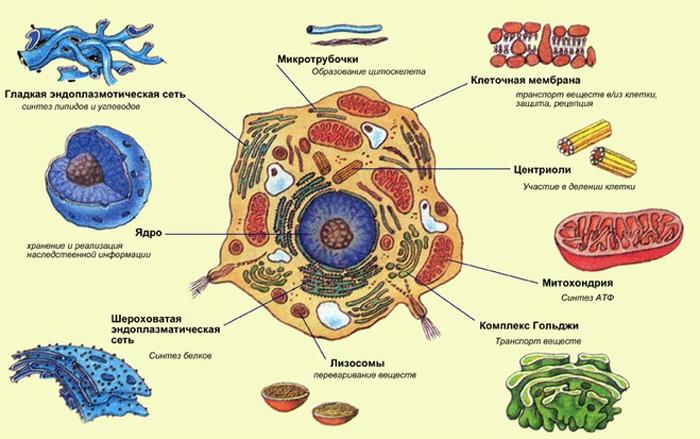
В живой растительной клетке основное вещество находится в постоянном движении. В движение, называемое током цитоплазмы илициклозом,вовлекается органеллы. Циклоз облегчает передвижение веществ в клетке и обмен ими между клеткой и окружающей средой.
Плазматическая мембрана.Представляет собой бислойную фосфолипидную структуру. Для растительных клеток свойственны впячивания плазматической мембраны.
Плазматическая мембрана выполняет следующие функции:
-участвует в обмене веществ между клеткой и окружающей средой;
-координирует синтез и сборку целлюлозных микрофибрилл клеточной стенки;
-передает гормональные и внешние сигналы, контролирующие рост и дифференцировку клеток.
Ядро.Это наиболее заметная структура в цитоплазме эукариотической клетки. Ядро выполняет две важные функции:
-контролирует жизнедеятельность клетки, определяя, какие белки, и в какое время должны синтезироваться;
-хранит генетическую информацию и передает её дочерним клеткам в процессе клеточного деления.
Ядро эукариотической клетки окружено двумя элементарными мембранами, образующиеядерную оболочку.Она пронизана многочисленными порами диаметром от 30 до 100 нм, видимыми только в электронный микроскоп. Поры имеют сложную структуру. Наружная мембрана ядерной оболочки в некоторых местах объединяется с эндоплазматическим ретикулумом. Ядерную оболочку можно рассматривать как специализированную, локально дифференцированную часть эндоплазматического ретикулума (ЭР).
Митохондрии.Как и хлоропласты, митохондрии окружены двумя элементарными мембранами. Внутренняя мембрана образует множество складок и выступов –крист,которые значительно увеличивают внутреннюю поверхность митохондрии.Они значительно меньше, чем пластиды, имеют около 0,5 мкм в диаметре и разнообразны по длине и форме.
В митохондриях осуществляется процесс дыхания, в результате которого органические молекулы расщепляются с высвобождением энергии и передачей её молекулам АТФ, основного резерва энергии всех эукариотических клеток.
Митохондрии, как и пластиды, являются полуавтономными органеллами, содержащими компонентами, необходимые для синтеза собственных белков. Внутренняя мембрана окружает жидкий матрикс, в котором находятся белки, РНК, ДНК, рибосомы, сходные с бактериальными и различные растворенные вещества. ДНК существует в виде кольцевых молекул, располагающихся в одном или нескольких нуклеоидах.
Микротельца.В отличие от пластид и митохондрий, которые отграничены двумя мембранами,микротельцапредставляют собой сферические органеллы, окруженные одной мембраной. Микротельца имеют гранулярное (зернистое) содержимое, иногда в них встречаются и кристаллические белковые включения. Микротельца связаны с одним или двумя участками эндоплазматического ретикулума.
Некоторые микротельца, называемыепроксисомами, играют важную роль в метаболизме гликолевой кислоты, имеющем непосредственное отношение к фотодыханию. В зеленых листьях они связаны с митохондриями и хлоропластами. Другие микротельца, называемые,глиоксисомами,содержат ферменты, необходимые для превращения жиров в углеводы. Это происходит во многих семенах во время прорастания.
Вакуоли –это отграниченные мембраной участки клетки, заполненные жидкостью –клеточным соком.Они окруженытонопластом (вакуолярной мембраной).
Молодая растительная клетка содержит многочисленные мелкие вакуоли, которые по мере старения клетки сливаются в одну большую. В зрелой клетке вакуолью может быть занято до 90% её объема. При этом цитоплазма прижата в виде тонкого периферического слоя к клеточной оболочке. Увеличение размера клетки в основном происходит за счет роста вакуоли. В результате этого возникает тургорное давление и поддерживается упругость ткани. В этом заключается одна из основных функций вакуоли и тонопласта.
Основной компонент сока – вода, остальные варьируют в зависимости от типа растения и его физиологического состояния. Вакуоли содержат соли, сахара, реже белки. Тонопласт играет активную роль в транспорте и накоплении в вакуоли некоторых ионов. Концентрация ионов в клеточном соке может значительно превышать ее концентрацию в окружающей среде. При высоком содержании некоторых веществ в вакуолях образуются кристаллы. Чаще всего встречаются кристаллы оксалата кальция, имеющие различную форму.
Вакуоли – места накопления продуктов обмена веществ (метаболизма). Это могут быть белки, кислоты и даже ядовитые для человека вещества (алкалоиды). Часто откладываются пигменты. Голубой, фиолетовый, пурпурный, темно-красный, пунцовый придают растительным клеткам пигменты из группы антоцианов. В отличие от других пигментов они хорошо растворяются в воде и содержатся в клеточном соке. Они определяют красную и голубую окраску многих овощей (редис, турнепс, капуста), фруктов (виноград, сливы, вишни), цветов (васильки, герани, дельфиниумы, розы, пионы). Иногда эти пигменты маскируют в листьях хлорофилл, например, у декоративного красного клена. Антоцианы окрашивают осенние листья в ярко-красный цвет. Они образуются в холодную солнечную погоду, когда в листьях прекращается синтез хлорофилла. В листьях, когда антоцианы не образуются, после разрушения хлорофилла заметными становятся желто-оранжевые каротиноиды хлоропластов. Наиболее ярко окрашены листья холодной ясной осенью.
Вакуоли участвуют в разрушении макромолекул, в круговороте их компонентов в клетке. Рибосомы, митохондрии, пластиды, попадая в вакуоли, разрушаются. По этой переваривающей активности их можно сравнить слизосомами– органеллами животных клеток.
Вакуоли образуются из эндоплазматической сети (ретикулума)
Рибосомы.Маленькие частицы (17 – 23нм), состоящие примерно из равного количества белка и РНК. В рибосомах аминокислоты соединяются с образованием белков. Их больше в клетках с активным обменом веществ. Рибосомы располагаются в цитоплазме клетки свободно или же прикрепляются к эндоплазматическому ретикулуму (80S). Их обнаруживают и в ядре (80S), митохондриях (70S), пластидах (70S).
Рибосомы могут образовывать комплекс, на которых происходит одновременный синтез одинаковых полипептидов, информация о которых снимается с одной молекулы и РНК. Такой комплекс называетсяполирибосомами (полисомами).Клетки, синтезирующие белки в больших количествах, имеют обширную систему полисом, которые часто прикрепляются к наружной поверхности оболочки ядра.
Эндоплазматический ретикулум.Это сложная трехмерная мембранная система неопределенной протяженности. В разрезе ЭР выглядит как две элементарные мембраны с узким прозрачным пространством между ними. Форма и протяженность ЭР зависят от типа клетки, ее метаболической активности и стадии дифференцировки. В клетках, секретирующих или запасающих белки, ЭР имеет форму плоских мешочков илицистерн,с многочисленными рибосомами, связанными с его внешней поверхностью. Такой ретикулум называетсяшероховатым эндоплазматическим ретикулумом.Гладкий ЭР обычно имеет трубчатую форму. Шероховатый и гладкий эндоплазматические ретикулумы могут присутствовать в одной и той же клетке. Как правило, между ними имеются много численные связи.
Эндоплазматический ретикулум функционирует как коммуникационная система клетки. Он связан с внешней оболочкой ядра. Фактически эти две структуры образуют единую мембранную систему. Когда ядерная оболочка во время деления клетки разрывается, ее обрывки напоминают фрагменты ЭР. Эндоплазматический ретикулум – это система транспортировки веществ: белков, липидов, углеводов, в разные части клетки. эндоплазматические ретикулумы соседних клеток соединяются через цитоплазматические тяжи –плазмодесмы –которые проходят сквозь клеточные оболочки.
Эндоплазматический ретикулум – основное место синтеза клеточных мембран. В некоторых растительных клетках здесь образуются мембраны вакуолей и микротелец, цистерныдиктиосом.
Микротрубочкиобнаружены практически во всех эукариотических клетках. Представляют собой цилиндрические структуры диаметром около 24 нм. Длина их варьирует. Каждая трубочка состоит из субъединиц белка, называемоготубулином.Субъединицы образуют 13 продольных нитей, окружающих центральную полость. Микротрубочки – это динамические структуры, они регулярно разрушаются и образуются на определенных стадиях клеточного цикла. Их сборка происходит в особых местах, которые называются центрами организации микротрубочек. В растительных клетках они имеют слабовыраженную аморфную структуру.
Функции микротрубочек: участвуют в образовании клеточной оболочки; направляют пузырьки диктиосом к формирующейся оболочке, подобно нитям веретена, которые образуются в делящейся клетке; играют определенную роль в формировании клеточной пластинки (первоначальной границы между дочерними клетками). Кроме того, микротрубочки – важный компонент жгутиков и ресничек, в движении которых, играют немаловажную роль.
Микрофиламенты,подобно микротрубочкам, найдены практически во всех эукариотических клетках. Представляют собой длинные нити толщиной 5 – 7 нм, состоящие из сократительного белка актина. Пучки микрофиламентов встречаются во многих клетках высших растений. По-видимому, играют важную роль в токах цитоплазмы. Микрофиламенты вместе с микротрубочками образуют гибкую сеть, называемуюцитоскелетом.
Основное веществодовольно долго считали гомогенным (однородный) богатым белком раствором с малым количеством структур или вообще бесструктурным. Однако в настоящее время, используя высоковольтный электронный микроскоп, было установлено, что основное вещество представляет трехмерную решетку, построенную из тонких (диаметром 3 – 6 нм) тяжей, заполняющих всю клетку. Другие компоненты цитоплазмы, включая микротрубочки и микрофиламенты, подвешены к этоймикротрабекулярной решетке.
Микротрабекулярная структура представляет собой решетку из белковых тяжей, пространство между которыми заполнено водой. Вместе с водой решетка имеет консистенцию геля, гель имеет вид студенистых тел.
К микротрабекулярной решетке прикреплены органеллы. Решетка осуществляет связь между отдельными частями клетки и направляет внутриклеточный транспорт.
Липидные капли– структуры сферической формы, придающие гранулярность цитоплазме растительной клетки под световым микроскопом. На электронных микрофотографиях они выглядят аморфными. Очень похожие, но более мелкие капли встречаются в пластидах.
Липидные капли, принимая за органеллы, называли их сферосомами и считали, что они окружены одно- или двуслойной мембраной. Однако последние данные показывают, что у липидных капель мембран нет, но они могут быть покрыты белком.
Эргастические вещества –это «пассивные продукты» протопласта: запасные вещества или отходы. Они могут появляться и исчезать в разные периоды клеточного цикла. Кроме зерен крахмала, кристаллов, антоциановых пигментов и липидных капель. К ним относятся смолы, камеди, танины и белковые вещества. Эргастические вещества входят в состав клеточной оболочки, основного вещества цитоплазмы и органелл, в том числе вакуолей.
Жгутики и реснички –это тонкие, похожие на волоски структуры, которые отходят от поверхности многих эукариотических клеток. Имеют постоянный диаметр, но длина колеблется от 2 до 150 мкм. Условно более длинные и немногочисленные из них называют жгутиками, а более короткие и многочисленные - ресничками. Четких различий между этими двумя типами структур не существует, поэтому для обозначения обоих используют терминжгутик.
У некоторых водорослей и грибов жгутики являются локомоторными органами, с помощью которых они передвигаются в воде. У растений (например, мхов, печеночников, папоротников, некоторых голосеменных) только половые клетки (гаметы) имеют жгутики.
Каждый жгутик имеет определенную организацию. Наружное кольцо из 9 пар микротрубочек окружает две дополнительные микротрубочки, расположенные в центре жгутика. Содержащие ферменты «ручки» отходят от одной микротрубочки каждой из наружных пар. Это основная схема организации 9 + 2 обнаружена во всех жгутиках эукариотических организмов. Считают, что движение жгутиков основано на скольжении микротрубочек, при этом наружные пары микротрубочек движутся одна вдоль другой без сокращения. Скольжение пар микротрубочек относительно друг друга вызывает локальное изгибание жгутика.
Жгутики «вырастают» из цитоплазматических цилиндрических структур, называемыхбазальными тельцами,образующимися и базальную часть жгутика. Базальные тельца имеют внутреннее строение, напоминающее строение жгутика, за исключением того, что наружные трубочки собраны в тройки, а не в пары, а центральные трубочки отсутствуют.
Плазмодесмы.Это тонкие нити цитоплазмы, которые связывают между собой протопласты соседних клеток. Плазмодесмы либо проходят сквозь клеточную оболочку в любом месте, либо сосредоточены на первичных поровых полях или в мембранах между парами пор. Под электронным микроскопом плазмодесмы выглядят как узкие каналы, выстланные плазматической мембранной. По оси канала из одной клетки в другую тянется цилиндрическая трубочка меньшего размера –десмотрубочка,которая сообщается с эндоплазматическим ретикулумом обеих смежных клеток. Многие плазмодесмы формируются во время клеточного деления, когда трубчатый эндоплазматический ретикулум захватывается развивающейся клеточной пластинкой. Плазмодесмы могут образовываться и в оболочках неделящихся клеток. Эти структуры обеспечивают эффективный перенос некоторых веществ от клетки к клетке.
Деление клеток.У многоклеточных организмов деление клеток наряду с увеличением их размеров является способом роста всего организма. Новые клетки, образовавшиеся во время деления, сходны по структуре и функциям, как с родительской клеткой, так и между собой. Процесс деления у эукариот можно подразделить на две частично перекрывающиеся стадии:митозицитокинез.
Митоз– это образование из одного ядра двух дочерних ядер, морфологически и генетически эквивалентных друг другу.Цитокинез –это деление цитоплазматической части клетки с образованием дочерних клеток.
Клеточный цикл.Живая клетка проходи ряд последовательных событий, составляющих клеточный цикл. Продолжительность самого цикла варьирует в зависимости от типа клетки и внешних факторов, например от температуры или обеспеченности питательными веществами. Обычно цикл делится наинтерфазу и четыре фазымитоза.
Интерфаза.Период между последовательными митотическими делениями.
Интерфазу делят на три периода, обозначаемые как G1, S, G2.
В период G1, который начинается после митоза. В этот период увеличивается количество цитоплазмы, включая различные органеллы. Кроме того, согласно современной гипотезе, в период G1 синтезируются вещества, которые либо стимулируют, либо ингибируют период S и остальную часть цикла, определяя, таким образом, процесс деления.
В период S следует за периодом G1, в это время происходит удвоение генетического материала (ДНК).
В период G2, который следует за S, формируются структуры, непосредственно участвующие в митозе, например, компоненты веретена.
Некоторые клетки проходит неограниченный ряд клеточных циклов. Это одноклеточные организмы и некоторые клетки зон активного роста (меристем). Некоторые специализированные клетки после созревания теряет способность к размножению. Третья группа клеток, например образующих раневую ткань (каллус), сохраняет способность делиться только в специальных условиях.
Митоз,или деление ядра. Это непрерывный процесс, подразделяемый на четыре фазы:профазу, метафазу, анафазу, телофазу. В результате митоза генетический материал, удвоившийся в интерфазе, делится поровну между двумя дочерними ядрами.
Одним из самых ранних признаков перехода клетки к делению служит появление узкого, кольцеобразного пояска из микротрубочек непосредственно под плазматической мембраной. Это относительно плотный поясок окружает ядро в экваториальной плоскости будущего митотического веретена. Так как он проявляется перед профазой, его называютпрепрофазным пояском.Он исчезает после митотического веретена, задолго до появления в поздней телофазе клеточной пластинки, которая растет от центра к периферии и сливается с оболочкой материнской клетки в области, ранее занятой препрофазным пояском.
Профаза.В начале профазы хромосомы напоминают длинные нити, разбросанные внутри ядра. Затем, по мере того как нити укорачиваются и утолщаются, можно увидеть, что каждая хромосома состоит не из одной, а из двух переплетенных нитей, называемыххроматидами.В поздней профазе две укороченные спаренные хроматиды каждой хромосомы лежат рядом параллельно, соединённые узким участком, называемымцентромерой.Она имеет определённое положение на каждой хромосоме и делит хромосому на два плеча различной длины.
Микротрубочки располагаются параллельно поверхности ядра вдоль оси веретена. Это само раннее проявление сборки митотического веретена.
К концу профазы ядрышко постепенно теряет чёткие очертания и наконец исчезает. Вскоре после этого распадается и ядерная оболочка.
Метафаза.В началеметафазы веретено,которое представляет трёхмерную структуру, наиболее широкую в средине и суживающуюся к полюсам, занимает место, прежде занятое ядром. Нити веретена – это пучки микротрубочек. Во время метафазы хромосомы, состоящие из двух хроматид каждая, располагаются так, что их центромеры лежат в экваториальной плоскости веретена. Своей центромерой каждая хромосома прикрепляется к нитям веретена. Однако, некоторые нити проходят от одного полюса к другому, не прикрепляясь к хромосомам.
Когда все хромосомы расположатся в экваториальной плоскости, метафаза завершится. Хромосомы готовы к делению.
Анафаза.Хроматиды каждой хромосомы расходятся. Теперь этодочерниехромосомы. Прежде всего, делится центромера, и две дочерние хромосомы увлекаются к противоположным полюсам. При этом центромеры движутся впереди, а плечи хромосом тянутся сзади. Нити веретена, прикрепленные к хромосомам, укорачиваются, способствуя расхождению хроматид и движению дочерних хромосом в противоположные стороны.
Телофаза.В телофазе завершается обособление двух идентичных групп хромосом, при этом вокруг каждой из них формируется ядерная мембрана. В этом активное участие принимает шероховатый ретикулум. Аппарат веретена исчезает. В ходе телофазы хромосомы теряют чёткость очертаний, вытягиваются, превращаясь снова в тонкие нити. Ядрышки восстанавливаются. Когда хромосомы становятся невидимыми, митоз завершается. Два дочерние ядра вступают в интерфазу. Они генетически эквивалентны друг другу и материнскому ядру. Это очень важно, так как генетическая программа, а вместе с ней и все признаки должны быть переданы дочерним организмам.
Продолжительность митоза варьирует у различных организмов и она зависит от типа ткани. Однако профаза самая длинная, а анафаза самая короткая. В клетках кончика корня продолжительность профазы составляет 1 – 2 ч; метафазы – 5 – 15 мин; анафазы – 2 – 10 мин; телофазы – 10 – 30 мин. Продолжительность интерфазы составляет от 12 до 30 ч.
Во многих эукариотических клетках центры организации микротрубочек, ответственные за формирование митотического веретена, связаны сцентриолями.