

30. Этапы энергетического обмена в клетке. Анаэробный этап. Брожение.
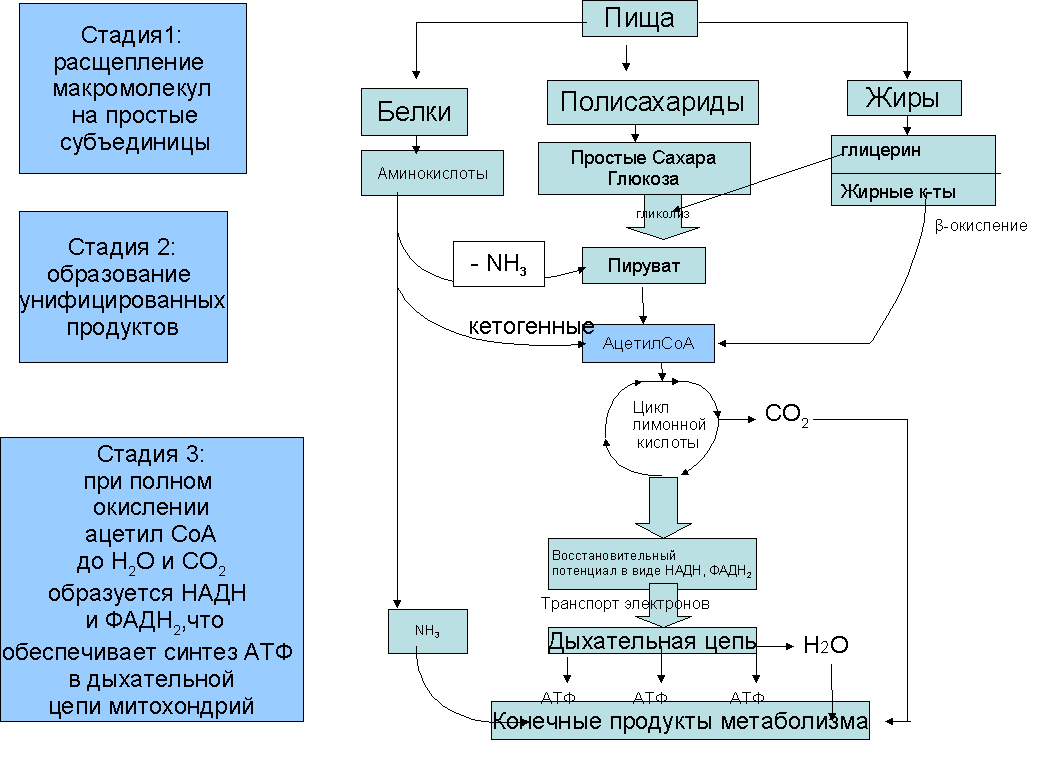
Первый этап— подготовительный. Пища поступает в организм животных и человека в виде сложных высокомолекулярных соединений. Прежде чем поступить в клетки и ткани, эти вещества должны разрушиться до низкомолекулярных, более доступных для клеточного усвоения веществ. На первом этапе происходит гидролитическое расщепление органических веществ, идущее при участии воды. Оно протекает под действием ферментов в пищеварительном тракте многоклеточных животных, в пищеварительных вакуолях одноклеточных, а на клеточном уровне — в лизосомах.
Второй этап осуществляется на клеточном уровне при отсутствии кислорода. Он протекает в цитоплазме клетки. Рассмотрим расщепление глюкозы, как одного из ключевых веществ обмена в клетке. Все остальные органические вещества (жирные кислоты, глицерин, аминокислоты) на разных этапах втягиваются в процессы ее превращения. Бескислородное расщепление глюкозы называется гликолизом.
Третий этап — биологическое окисление, или дыхание. Этот этап протекает только в присутствии кислорода и иначе называется кислородным. Он протекает в митохондриях.
Начальные этапы окисления углеводов происходят в цитозоле и не требуют участия кислорода, поэтому эта стадия процесса называется анаэробным окислением, или гликолизом. Главным субстратом окисления при анаэробном получении энергии служат гексозы и в первую очередь глюкоза. Процесс гликолиза включает 9 последовательных ферментативных реакций, в результате которых молекула глюкозы превращается в две молекулы пирувата. В ходе некоторых из этих реакций происходит восстановление НАД+ до НАДН и перенос неорганического фосфата на АДФ с образованием высокоэнергетической связи АТФ. В глюкозе количество потенциальной энергии, заключенной в связях между атомами С, Н и О, составляет около 680 ккал на 1 моль. Эта энергия освобождается при полном окислении глюкозы: С6Н12О6+6О2=6Н2О+6СО2+680ккал.
В процессе гликолиза происходит неполное окисление субстрата. В результате него глюкоза распадается до триоз, при этом тратятся 2 молекулы АТФ и синтезируются 4 молекулы АТФ, так что в конечном результате клетка получает всего 2 молекулы АТФ. В энергетическом отношении этот процесс малоэффективен, поэтому из 680 ккал, заключающихся в связях 1 моля глюкозы, освобождается менее 10% энергии. Несмотря на низкий энергетический выход, анаэробное окисление, гликолиз, широко используется в живой природе. Эритроциты млекопитающих, например, получают всю необходимую им энергию за счет гликолиза, так как у них нет митохондрий. Конечные продукты гликолиза - триозы, в первую очередь пировиноградная кислота, все еще несут большое количество химической энергии и вовлекаются в дальнейшее окисление, происходящее в самих митохондриях.
31. Этапы энергетического обмена в клетке. Аэробный этап.
Про этапы см. 30 вопрос.
Окислительные процессы в митохондриях связаны с окислительным циклом трикарбоновых кислот и с дыхательной цепью переноса электронов. Они начинаются после образования в матриксе достаточного количества ацетил-КоА из пирувата и жирных кислот. Пируват, образовавшийся в результате гликолиза, перемещается из цитозоля в матрикс митохондрии, где теряет молекулу СО и, окисляясь до ацетата, соединяется с коферментом А. Жирные кислоты в митохондриальном матриксе окисляются, молекула жирной кислоты последовательно укорачивается на два углеродных атома, и образуется ацетил-КоА. Ферменты, катализирующие эти реакции, как и большинство ферментов цикла трикарбоновых кислот, находятся в матриксе митохондрий.
Главная функция цикла лимонной кислоты-окисление ацетата, который включается в цикл в форме молекул ацетил-КоА. Цикл начинается взаимодействия ацетил-КоА с оксалоацетатом, что приводит к образованию цитрата. Далее в ходе семи последовательных ферментативных реакций два атома углерода удаляются в виде углекислого газа, и в конце концов регенерируется оксалоацетат. Каждый оборот цикла дает две молекулы углекислого газа, который диффундирует из митохондрий и покидает клетку. Энергия, высвобождающаяся при окислении химических связей лимонной кислоты, потребляется в цикле трикарбоновых кислот несколькими способами. Во-первых, в одной из реакций цикла синтезируется одна молекула АТФ. Во-вторых, в ходе реакций цикла лимонной кислотыпроисходит извлечение высокоэнергетических электронов, которые используются для восстановления молекул-переносчиков водорода НАД (никотинамидадениндинуклеотида) и ФАД(флавинадениндинуклеотида). В каждом обороте цикла три молекулы НАД превращаются в НАДН и одна молекула ФАД-в ФАДН. Образование НАДН происходит не только в ходе реакций цикла лимонной кислоты, но и на более ранних этапах окисления веществ: в процессе гликолиза и при образовании ацетил-КоА из пирувата. Энергия, переносимая молекулами НАДН и ФАДН далее используется в реакциях окислительного фосфорилирования для синтеза АТФ.
Окислительное фосфорилирование связано с переносом электронов от НАДН и ФАДН по дыхательной цепи к молекулярному кислороду и использованием энергии для синтеза АТФ. Эти процессы протекают на внутренней мембране митохондрий, в которой находятся белки-переносчики дыхательной цепи и комплексы АТФ-синтетазы.
Дыхательная цепь внутренней мембраны митохондрий содержит три главных ферментных комплекса, участвующих в переносеэлектронов с НАДН на кислород. Первый, НАДН-дегидрогеназный, комплекспринимает электроны от HAДH и переносит их во второй комплекс-комплекс цитохромов b-с, который переносит их на цитохромоксидазный комплекс, передающий электроны на кислород, в результате чего образуется вода. На этом процесс окисления заканчивается (конечными продуктами окисления исходного субстрата являются углекислый газ и вода). Перенос электронов по дыхательнойцепи происходит с участием убихинона и цитохрома с-мобильных переносчиков, передвигающихся в мембране от одного комплекса к другому и обратно. Переносчики электронов расположены и дыхательной цепи таким образом, что каждый последующий комплекс обладает большим сродством к электронам, чем предыдущий. Электроны перемещаются от одного комплекса к другому, пока не перейдут на кислород, имеющий наибольшее сродство к электрону. Перенос электронов сопровождается их переходом на более низкие энергетические уровни и выделением энергии. Элементы дыхательной цепи последовательно окисляются и восстанавливаются, в результате чего энергия высвобождается небольшими порциями и используется для перекачивания протонов из матрикса митохондрии через внутреннюю митохондриальную мембрану в межмембранное пространство и далее за пределы митохондрии. Перенос протонов, осуществляемый компонентами дыхательной цепи, приводит к возникновению электрохимического протонного градиента, который складывается из мембранного потенциала и градиента pH. Энергия электрохимического протонного градиента используется для синтеза АТФ.
Преобразование энергии окислительных реакций в энергию АТФ происходит на внутренней митохондриальной мембране в результате хемиосмоса. При достижении определенной разности потенциалов (220 мВ) белковый комплекс АТФ-синтетазы начинает транспортировать протоны обратно в матрикс и при этом превращает одну форму энергии в другую: образует АТФ из АДФ и неорганического фосфата. АТФ-синтетаза состоит из двух основных частей: мембранного компонента F0, осуществляющего транспорт протонов через мембрану, и сопрягающего фактора F1, осуществляющего синтез АТФ.
Окислительные процессы в митохондриях и фосфорилирование АДФ можно разобщить, сняв разность потенциалов на митохондриальной мембране, сделав в ней диффузионные каналы, а также механическим нарушением, либо с помощью различных химических соединений. Перенос электронов и окисление субстрата будут продолжаться, но синтеза АТФ не будет. Разобщение метаболических процессов окисления и фосфорилирования приводит к образованию значительного количества тепла вместо накопления энергии в форме макроэргических соединений.
Суммарное уравнение: С6Н12О6 + 6О2 → 6СО2 + 6Н2О + 38АТФ + Q
32. Гипотеза эволюционного происхождения митохондрий, хлоропластов, ядра, ЭПР. Возникновение клеточной организации в ходе эволюции. (нет) Теорию эндосимбиотического происхождения хлоропластов впервые предложил в 1883 годуАндреас Шимпер, показавший их саморепликацию внутри клетки. А. С. Фаминцин и О. В. Баранецкий-о двойственной природе лишайников - комплекса гриба и водоросли. К. С. Мережковский в 1905 году предложил само название «симбиогенез», впервые детально сформулировал теорию и даже создал на её основе новую систему органического мира. Фаминцин в 1907 году, опираясь на работы Шимпера, также пришёл к выводу, что хлоропласты являются симбионтами, как и водоросли в составе лишайников. В 1920-е Б. М. Козо-Полянский- симбионтами являются и митохондрии.
Митохндрии — это потомки аэробных бактерий (прокариот),поселившихся некогда в предковой эукариотической клетке и «научившимися» жить в ней в качестве симбионтов.Пластиды, подобно митохондриям, имеют свои собственные прокариотические ДНК и рибосомы. По-видимому, хлоропласты произошли от фотосинтезирующих бактерий, поселившихся в своё время в гетеротрофных клетках протистов, превратив их в автотрофные водоросли.
Доказательства Митохондрии и пластиды:• имеют две полностью замкнутые мембраны. При этом внешняя сходна с мембранами вакуолей, внутренняя — бактерий.• размножаются бинарным делением (причём делятся иногда независимо от деления клетки)• генетический материал — кольцевая ДНК, не связанная с гистонами (ДНКмитохондрий и пластид ближе к ДНК бактерий, чем к ядерной ДНК эукариот)• имеют свой аппарат синтеза белка— рибосомы и др.
Смешение у эукариот многих свойств, характерных для архей и бактерий, позволило предположить симбиотическое происхождение ядра от метаногенной архебактерии, внедрившейся в клетку миксобактерии. В последнее десятилетие сформировалась также гипотеза вирусного эукариогенеза (англ.viral eukaryogenesis). В её основании лежит ряд сходств устройства генетического аппарата эукариот и вирусов: линейное строение ДНК, её тесное взаимодействие с белками и др. Было показано сходство ДНК-полимеразы эукариот и поксивирусов, что сделало именно их предков основными кандидатами на роль ядра.