

гиста ответы
.pdfВторичные фолликулы. Дальнейший рост фолликула обусловлен разрастанием однослойного фолликулярного эпителия и превращением его в многослойный эпителий. Эпителий секретирует фолликулярную жидкость (liquor folliculi), которая накапливается в формирующейся полости фолликула и содержит стероидные гормоны (эстрогены). При этом овоцит с окружающими его вторичной оболочкой и фолликулярными клетками в виде яйценосного бугорка (cumulus oophorus) смещается к одному полюсу фолликула. В дальнейшем в теку врастают многочисленные кровеносные капилляры и она дифференцируется на два слоя — внутренний и наружный. Во внутренней теке (theca interna) вокруг разветвляющихся капилляров располагаются многочисленные интерстициальные клетки, соответствующие интерстициальным клеткам семенника (гландулоцитам). Наружная тека (theca folliculi externa) образована плотной соединительной тканью.
Такие фолликулы, в которых формируется фолликулярная полость, а тека состоит из двух слоев, - называются уже вторичными фолликулами (folliculus secundorius). Овоцит в этом фолликуле уже не увеличивается в объеме, хотя сами фолликулы за счет накопления в их полости фолликулярной жидкости резко увеличиваются. При этом овоцит с окружающим его слоем фолликулярных клеток оттесняется к верхнему полюсу растущего фолликула. Этот слой фолликулярных клеток получил название «лучистый венец», или corona radiata.
Зрелый фолликул, достигший своего максимального развития и включающий одну полость, заполненную фолликулярной жидкостью, называется третичным, или пузырчатым фолликулом (folliculus ovaricus tertiarius), или же Граафовым пузырьком. Клетки лучистого венца, непосредственно окружающие растущий овоцит, имеют длинные ветвистые отростки, проникающие через блестящую зону и достигающие поверхности овоцита. По этим отросткам к овоциту от фолликулярных клеток поступают питательные вещества, из которых в цитоплазме синтезируются липопротеиды желтка, а также другие вещества.
Пузырчатый (третичный) фолликул достигает такого размера, что выпячивает поверхность яичника, причем яйценосный бугорок с овоцитом оказывается в выступающей части пузырька. Дальнейшее увеличение объема пузырька, переполненного фолликулярной жидкостью, приводит к растягиванию и истончению как его наружной оболочки, так и белочной оболочки яичника в месте прилегания этого пузырька с последующим разрывом и овуляцией.
Между фолликулами встречаются атретические тела (corpus atreticum). Они формируются из фолликулов, прекративших свое развитие на разных стадиях.
Под корковым веществом в яичнике располагается мозговое вещество (medulla ovarii), состоящее из соединительной ткани, в которой проходят магистральные кровеносные сосуды и нервы, а также эпителиальные тяжи — остатки канальцев первичной почки.
Генеративная функция яичников Овогенез
Овогенез отличается от сперматогенеза рядом особенностей и проходит в три стадии: размножения; роста; созревания
40
1.Клеточная мембрана. Строение и функции. Типы межклеточных соединений.
Структурной основой мембран является двойной слой липидов, в который включены молекулы белка. К липидам относится большая группа органических веществ, обладающих плохой растворимостью в воде (гидрофобность) и хорошей растворимостью в органических растворителях (липофильность). Состав липидов, входящих в мембраны клеток, очень разнообразен. Характерными представителями липидов, встречающихся в клеточных мембранах, являются фосфолипиды, сфингомиелины и холестерин (в растительных клетках не обнаружен). Характерной особенностью липидов мембран является разделение их молекулы на две функционально различные части: неполярные хвосты, состоящие из жирных кислот, и заряженные полярные головки.
Обязательным компонентом клеточных мембран являются белки (протеины). В среднем они составляют 50% массы мембраны (в мембранах митохондрий на долю белков приходится около 75%, а в плазматической мембране клеток миелиновой оболочки – около 25%).
Выявлено два типа мембранных белков. Белки первого типа, называемые периферическими белками, связаны с мембраной в основном ионными взаимодействиями. Мембранные белки второго типа называют интегральными белками. Эти протеины или погружены в толщу липидного бислоя, или пронизывают
мембрану насквозь (трансмембранные белки). Интегральные белки перемещаются в плоскости мембраны. По биологической роли мембранные белки можно разделить на три группы: ферменты, рецепторные белки и структурные белки.
Особое место среди клеточных мембран занимает плазматическая мембрана или плазмалемма. Плазматической мембраной называется барьер, который окружает цитоплазму, определяя границы клетки. Она имеет толщину около 10 нм и представляет собой самую толстую из клеточных мембран. Большая ее толщина обусловлена тем, что на ее внутренней стороне локализован слой периферических белков. На наружной стороне клеток животных располагается слой углеводных компонентов (гликокаликс), а у растений – клеточная стенка.
Функции мембран:
Ограничение клетки от внешней среды, поддержание формы клетки;
Обеспечение транспорта различных веществ как внутрь клетки, так и из нее. Различают активный и пассивный типы транспорта. К пассивным механизмам относят диффузию, облегченную диффузию и осмос, к активным – работу белковых ионных насосов, эндоцитоз и экзоцитоз;
Восприятие сигналов и передача их внутрь клетки. На поверхности плазмалеммы располагаются различные рецепторные структуры (для гормонов, медиаторов и др.), специфически взаимодействующие с внеклеточными факторами и с соседними клетками;
Отдельные участки плазматической мембраны в специализированных клетках животных принимают участие в построении специальных отростков клетки, таких, как микроворсинки, реснички, рецепторные выросты и др.;
Плазматическая мембрана играет важную роль при делении клетки;
Участие в биохимических процессах, поскольку большинство ферментов связано с мембранами;
Формирование мембранных структур клетки;
Плазматическая мембрана принимает участие в образовании межклеточных взаимодействий у многоклеточных организмов, что способствует формированию тканей.
ЗАМЫКАЮЩИЕ Простой контакт — соединение клеток за счет пальцевидных впячиваний и выпячиваний цитомембран
соседних клеток. Специфических структур, формирующих контакт, нет.
Плотный замыкающий контакт — соприкасаются билипидные слои мембран соседних клеток. В области зоны плотных контактов между клетками не проходят практически никакие вещества.
АДГЕЗИОННЫЕ Межклеточные адгезионные соединения:
Точечные — контакт образуется на небольшом по площади участке цитомембран соседних клеток. Адгезионные пояски — контакт окружает по периметру всю клетку в виде пояса, располагается в верхних отделах боковых поверхностей эпителиальных клеток.
В области контакта в цитомембрану встроены специальные трансмембранные белки — кадгерины, которые соединяются с кадгеринами другой клетки.
Для соединения кадгеринов нужны ионы кальция.
Со стороны цитоплазмы к кадгеринам присоединяются белки ,бета-катенин, альфа-катенин, гаммакатенин, PP-120, EB-1, и к ним присоединяются актиновые микрофиламенты.
Адгезионные соединения между клеткой и внеклеточным матриксом:
Контакт образуется на небольшом по площади участке.
В месте контакта в цитомембрану встроены трансмембранные белки альфа- и бета-интегрины, которые соединяются с элементами межклеточного матрикса.
|
Со стороны цитоплазмы к интегринам присоединяются несколько промежуточных белков (тензин, |
|
талин, альфа-актинин, винкулин, паксилин, фокальная адгезионная киназа), к которым присоединяются |
|
актиновые микрофиламенты. |
Десмосомы:
Контакт образуется на небольшом по площади участке.
В месте контакта в цитомембрану встроены трансмембранные белки десмоглеин и десмоколин, которые соединяются с такими же белками другой клетки.
Для соединения десмоколинов и десмоглеинов нужны ионы кальция.
Со стороны цитоплазмы к десмоколину и десмоглеину присоединяются промежуточные белки —
десмоплакин и плактоглобин, к которыем присоединяются промежуточные филаменты.
Все типы адгезионных контактов, кроме механического скрепления клеток между собой и с внеклеточным матриксом, передают информацию о состоянии окружающей среды внутрь клетки (в цитоплазму и ядро). При этом ход различных биологических процессов в клетке может меняться.
ПРОВОДЯЩИЕ Нексусы (щелевидные контакты):
Контакт образуется на небольшом по площади участке.
В месте контакта в цитомембрану встроены трансмембранные белки коннексины, которые соединяются между собой и образуют водный канал в толще мембраны — конексон.
Коннексоны контактирующих клеток соединяются (или сопоставляются), в результате чего между соседними клетками образуется канал, с помощью которого из одной клетки в другую (в обоих направлениях) свободно проходит вода, малые молекулы и ионы, а также электрический ток.
Синапсы — см. раздел по нервной ткани. Обеспечивают передачу потенциала действия (нервного импульса) с нервной клетки на другую нервную или иную клетку.
2 .Костная ткань. Способы остеогенеза. Развитие кости на месте хряща.
Костные ткани (textus ossei) — это специализированный тип соединительной ткани с высокой минерализацией межклеточного органического вещества, содержащего около 70 % неорганических соединений, главным образом фосфатов кальция.
Функции: опорная; защитная; депо Ca, Р.
Межклеточное вещество костной ткани состоит:
1.Неорганические соединения (фосфорнокислые и углекислые соли кальция, кристаллы гидроксиапатита) - составляют 70% межклеточного вещества.
2.Вода – 25%.
3.Органическая часть межклеточного вещества представлена коллагеновыми (синоним - оссеиновыми) волокнами и аморфной склеивающей массой (оссеомукоид) - составляет 25%.
Непрямой остеогенез – образование кости на месте гиалинового хряща. Таким образом образуются все трубчатые кости. На месте будущей кости из гиалинового хряща формируется зачаток трубчатой кости,
снаружи он покрыт надкостницей. Этот процесс протекает на втором месяце эмбриогенеза. Далее в области диафиза между надкостницей и веществом хряща образуется из грубоволокнистой костной ткани перихондральная кость или перихондральная костная манжетка, которая полностью окружает вещество хряща в зоне диафиза и тем самым нарушает поступление питательных веществ из надхрящницы в хрящ. Это вызывает частичное разрушение гиалинового хряща в диафизе, а остатки хряща обызветствляются. Надхрящница превращается в надкостницу, и из надкостницы кровеносные сосуды пронизывают костную манжетку. При этом грубоволокнистая ткань костной манжетки разрушается и замещается пластинчатой костной тканью. Кровеносные сосуды глубоко врастают в диафиз, вместе с ними проникают остеобласты, остекласты и мезенхимные клетки. Остеокласты постепенно разрушают обызвествленый хрящ,
аостеобласты вокруг участков обызвествленного хряща образуют пластинчатую костную ткань, которая формирует эндохондральную кость. Перихондральная и эндохондральная костные ткани разрастаются, соединяются, остеокласты начинают разрушать костную ткань в средней части диафиза, и постепенно формируется костномозговой канал (полость). Из мезенхимы закладывается красный костный мозг.
Позднее осуществляется окостенение эпифиза, между эпифизами и диафизом сохраняется метаэпифизарный хрящ (зона роста кости). За счет этой пластинки кость растет в длинну. В ней выделяют пузырчатый слой на границе с диафизом, содержащий разрушающиеся клетки. Затем идет столбчатый слой, в котором молодые хондроциты образуют ряды. Молодые хондроциты пролиферируют, образуют межклеточное вещество. Также выделяют пограничный слой, имеющий строение типичного гиалинового хряща. Эти пластинки окостеневают последними.
Костная ткань в общем и кости в частности хорошо регенерируют за счет метаэпифизарных стволовых клеток надкостницы. В начале с помощью фибробластов надкостницы образуется рыхлая соединительная ткань. Далее активируются остеобласты, вырабатывающие грубоволокнистую костную ткань. В течение первых двух недель она заполняет зону повреждения и формирует костные мозоли. Со 2 недели в костные мозоли внедряются кровеносные сосуды и грубоволокнистая костная ткань замещается пластинчатой костной тканью.
3 Печень. Источники развития, особенности кровоснабжения. Строение классической печеночной дольки.
Это самая крупная железа в организме.
Функции печени. Самая главная - дезинтоксикация продуктов обмена. Здесь разрушаются токсические соединения. Из аммиака образуется мочевина. Здесь разрушаются лекарственные препараты. Поэтому если исключить только эту функцию, то человек умрѐт через 2-3 суток. В печени вырабатывается фибриноген, белки крови, альбумины, протромбин, некоторые фракции глобулинов. Здесь депонируются витамины А, Д, Е, К. Печень участвует в реализации белкового обмена, т.к. здесь заканчивается последняя внутриклеточная часть пищеварения. Печень имеет отношение к обмену углеводов. Здесь также происходит синтез гликогена и образование глюкозы под действием инсулина и глюкагона.
Печень имеет большое значение как мощный антибактериальный барьер при помощи макрофагов печени – купферовские клетки. Печень вырабатывает холестерин, который входит в состав плазмолеммы. Печень вырабатывает желчные кислоты, которые необходимы для эмульгирования жиров в кишке. Это единственная экзокринная функция. Ибо все остальные продукты поступают непосредственно в кровь.
РАЗВИТИЕ
Печень закладывается, как и поджелудочная железа, в конце 3 недели эмбриогенеза из эпителия среднего отдела кишки. Из мезенхимы образуется капсула и соединительная ткань, которая разделяет железу на доли, сегменты и дольки. В этих соединительнотканных прослойках проходят сосуды. Поэтому соединительная ткань в печени развита очень слабо. Соединительная ткань в печени между дольками у человека развита только в патологических условиях, т.е. когда разрушаются клетки паренхимы и замещаются соединительной тканью, т.е. развивается цирроз печени. Печень как железа представлена дольками. Между ними прослойки соединительной ткани выражены слабо.
Между дольками проходят 3 типа сосудов:
1.Междольковая артерия – результат ветвления печѐночной артерии.
2.Междольковая вена, которая образуется как разветвления воротной вены.
3.Междольковый желчный проток, который выходит из дольки, но располагается в обратном направлении.
Эти 3 сосуда носят название печѐночная триада.
Вена крупных размеров, т.к. она приносит всю кровь из ЖКТ, богатую продуктами. Эта кровь содержит кислород.
Артерия отходит от аорты. Она небольших размеров и несѐт в основном кислород в печень. Эти сосуды подходят к дольке, разветвляются на вокругдольковые. Артерии и вены распадаются на артериальные и венозные капилляры, которые соединяют на периферии дольки, образуя синусоидные капилляры. Они идут от периферии дольки к центру. В центре они сливаются и образуют один сосуд – центральную вену. Из неѐ начинается отток венозной крови в поддольковые вены, печѐночные вены, которые идут в венозную систему. Поддольковые вены легко увидеть. Таким образом, печѐночная долька имеет вид 6-гранной усечѐнной призмы, по периферии, к которой располагается 5-6 триад. А в центре всегда – центральная вена. И к этой центральной вене радиально идут синусоидные капилляры.
Дольки свободно граничат друг с другом без выраженных прослоек. Поэтому структура печени имеет определѐнную губчатость. Эпителий образует в дольке секреторные отделы. Они идут радиально от периферии к центру, образуя печѐночные балки или пластинки. Таким образом, печѐночные балки располагаются между капиллярами. Они могут анастамозировать друг с другом. Строгая радиальность у человека утрачивается. Каждая балка представлена 2-мя рядами клеток гепатоцитов. Внутри печѐночной балки расположен щелевидный капилляр – это желчный капилляр, который начинается слепо в центральной части дольки и открывается на периферии в междольковый желчный проток. Снаружи этой пластинки – кровеносные капилляры. Поэтому каждый гепатоцит одной частью открывается в желчный капилляр – биллиарная часть, а другой – с кровеносными капиллярами – васкулярная часть. Поэтому желчь поступает только в желчный.
41билет
1.Жизненный цикл клетки. Его этапы, особенности у различных видов клеток.
Клеточный цикл — это период жизни клетки от одного деления до другого или от деления до смерти. Клеточный цикл состоит из интерфазы (период вне деления) и самого клеточного деления.
В интерфазе выделяют три стадии: G1 – пресинтетическая, S – синтетическая и G2 – постсинтетическая.
После деления клетка вступает в фазу G1. Клетка растет, и в ней синтезируются РНК, белки и различные вещества, необходимые для удвоения хромосом, увеличивается количество рибосом и митохондрий. Набор генетического материала можно представить как 2n2c (диплоидный), где n – количество хромосом (центромер), c – количество молекул ДНК (хроматид). Часть клеток из этой фазы переходит в фазу G0, это могут быть специализированные клетки, которые больше не делятся, а выполняют определенные функции (например, эритроциты, миоциты, нейтрофилы и др) или стволовые клетки, которые в этой стадии могут находится от нескольких часов до нескольких десятков лет. Но большинство клеток, накопив необходимые вещества и восстановив свой размер, а иногда и без изменения размеров после предыдущего деления, начинают подготовку к следующему делению.
Фаза S – фаза репликации количества ДНК (2n4c). При подготовке к делению происходит удвоение молекул ДНК, на каждой хромосоме синтезируется ее копия. Пока эти хромосомы после удвоения не расходятся, каждая хромосома в этой паре называется хроматидой. После репликации ДНК конденсируется, хромосомы приобретают более компактную укладку, и в таком состоянии их можно увидеть в световом микроскопе. Между делениями эти хромосомы не столь конденсированы и в большей степени расплетены. Понятно, что в конденсированном состоянии им трудно функционировать. Раньше считалось, что между делениями клетки хромосомная ДНК (хроматин) находится в полностью расплетенном состоянии, но сейчас выясняется, что структура хромосом достаточно сложная и степень деконденсации хроматина между делениями не очень велика (рис.7.1.2).
После того, как хромосомы удвоились, клетка переходит в фазу G2 – постсинтетический период или фазу подготовки в митозу. В этот период происходит активное накопление энергии и ферментов, необходимых для последующего деления (2n4c). После G2-фазы наступает непрямое деление клетки или митоз. Собственно митотическое деление занимает лишь незначительную часть клеточного цикла, после деления клетки цикл повторяется заново.
Митоз (от греч. mitos - нить) - деление ядра, следующее за репликацией хромосом, в результате чего дочерние ядра содержат то же число хромосом, что и родительские. Этот вид деления клеток был впервые описан немецким гистологом В. Флемингом в 1882 г. При митотическом делении ядро клетки претерпевает ряд строго упорядоченных последовательных изменений с образованием специфических нитчатых структур. В митозе выделяют несколько фаз: профазу, метафазу, анафазу и телофазу.
Профаза . В начале профазы многочисленные цитоплазматические микротрубочки, входящие в состав цитоскелета, распадаются; при этом образуется большой пул свободных молекул тубулина. Эти молекулы вновь используются для построения главного компонента митотического аппарата - митотического веретена. Каждая пара центриолей становится частью митотического центра, от которого лучами расходятся микротрубочки (фигура "звезда"). Вначале обе звезды лежат рядом около ядерной мембраны. В поздней профазе пучки полюсных микротрубочек, взаимодействующие друг с другом (и видимые в световой микроскоп как полюсные нити), удлиняются и как будто расталкивают два митотических центра друг от друга вдоль наружной поверхности ядра. Таким способом образуется биполярное митотическое веретено.
В этот период можно наблюдать двойную природу хромосом, т.к. каждая хромосома выглядит продольно удвоенной, состоит из двух хроматид (4с). Эти половинки хромосом, называемые сестринскими хроматидами, удерживаются вместе одним общим участком – центромерой (2n). Начинается расхождение центриолей к полюсам и образование веретена деления (2n4c).
Метафаза . Эта стадия митоза часто продолжается длительное время. Все хромосомы располагаются таким образом, что их центромеры лежат в одной плоскости (метафазная пластинка). На этой стадии хромосомы имеют самую малую длину, поскольку в это время они наиболее сильно спирализованы и конденсированы. Эта стадия наиболее пригодна для подсчета числа хромосом в клетке, изучения и описания их строения, определения размеров и т.д.
За ориентацию хромосом перпендикулярно оси митотического веретена и расположение их на равном расстоянии от обоих полюсов веретена, скорее всего, ответственны кинетохорные нити. Вероятно, такое расположение хромосом в метафазной пластинке обусловлено способом создания тянущей силы в митотическом веретене: этот способ таков, что сила, действующая на кинетохорные нити тем слабее, чем ближе к полюсу находятся кинетохоры. Каждая хромосома удерживается в метафазной пластинке парой кинетохоров и двумя пучками связанных с ними нитей, идущих к противоположным полюсам веретена. Метафаза резко оканчивается разделением двух кинетохоров каждой хромосомы (2n4c).
Анафаза . Продолжается обычно всего несколько минут (рис. 7.1.7). Анафаза начинается внезапным расщеплением каждой хромосомы, которое обусловлено разделением сестринских хроматид в точке их соединения в центромере. С этого момента сестринские хроматиды можно называть дочерними хромосомами, т.к. каждая из них имеет свою центромеру - n.
Это расщепление, разделяющее кинетохоры, не зависит от других событий митоза и происходит даже в хромосомах, не прикрепленных к митотическому веретену; оно позволяет полярным силам веретена, действующим на метафазную пластинку, начать перемещение каждой хроматиды к соответствующим полюсам веретена со скоростью порядка 1 мкм/мин. Во время этого анафазного движения кинетохорные нити укорачиваются по мере того, как хромосомы приближаются к полюсам. Примерно в это же время удлиняются нити митотического веретена и два полюса веретена расходятся еще дальше. Этим обеспечивается согласованное и точное распределение хромосомного материала в дочерние клетки (4n4c).
Телофаза . В заключительной стадии митоза телофазе разделенные дочерние хроматиды подходят к полюсам, кинетохорные нити исчезают. После удлинения полюсных нитей вокруг каждой группы дочерних хроматид образуется новая ядерная оболочка. Конденсированный хроматин начинает разрыхляться, появляются ядрышки, и митоз заканчивается (2n2c).
Цитокинез . Процесс деления цитоплазмы - цитокинез, проходит под действием сократимого кольца и начинается обычно в поздней анафазе или телофазе. Мембрана в средней части клетки (между двумя дочерними ядрами) начинает втягиваться внутрь в плоскости метафазной пластинки под прямым углом к длинной оси митотического
2.Артерии. Классификация, развитие, строение. Взаимосвязь структуры артерии и гемодинамических условий.
А. Кровеносные сосуды:
1. Артериальное звено: а)артерии эластического типа; б) артерии мышечного типа; в) артерии смешанного типа.
Кровеносные сосуды развиваются из мезенхимы. (Первые кровеносные сосуды появляются в мезенхиме стенки желточного мешка на 2-3-й неделе эмбриогенеза). В эмбриогенезе наиболее ранний период характеризуется появлением многочисленных клеточных скоплений мезенхимы в стенке желточного метка - кровяных островков. Внутри островка образуются кровяные клетки и формируется полость, а расположенные по периферии клетки становятся плоскими, соединяются между собой при помощи клеточных контактов и формируют эндотелиальную выстилку образующейся трубочки. Такие первичные кровеносные трубочки по мере образования соединяются между собой и формируют капиллярную сеть. Окружающие клетки мезенхимы превращаются в перициты, гладкие мышечные клетки и адвентициальные клетки. В теле зародыша кровеносные капилляры закладываются из клеток мезенхимы вокруг щелевидных пространств, заполненных тканевой жидкостью. Когда по сосудам усиливается кровоток, эти клетки становятся эндотелиальными, а из окружающей мезенхимы формируются элементы средней и наружной оболочки. В конце 3-й недели внутриутробного развития сосуды тела зародыша начинают сообщаться с сосудами внезародышевых органов.
Сосудистая система обладает очень большой пластичностью. Прежде всего отмечается значительная изменчивость густоты сосудистой сети, так как в зависимости от потребностей органа в питательных веществах и кислороде в широких пределах колеблется количество приносимой ему крови. Изменение скорости кровотока и кровяного давления ведет к образованию новых сосудов и перестройке имеющихся сосудов. Происходит превращение мелкого сосуда в более крупный с характерными особенностями строения его стенки. Наибольшие изменения возникают в сосудистой системе при развитии окольного, или коллатерального, кровообращения.
Артерии бывают трех типов: эластического, мышечного и смешанного (или мышечно-
эластического). Классификация основывается на соотношении количества мышечных клеток и эластических волокон в средней оболочке артерий.
Артерии эластического типа
Артерии эластического типа характеризуются выраженным развитием в их средней оболочке эластических структур. К этим артериям относятся аорта илегочная артерия, в которых кровь протекает под высоким давлением и с большой скоростью. В эти сосуды кровь поступает непосредственно из сердца. Артерии крупного калибра выполняют главным образом транспортную функцию. Наличие большого количества эластических элементов (волокон, мембран) позволяет этим сосудам растягиваться при систоле сердца и возвращаться в исходное положение во время диастолы. В качестве примера сосуда эластического типа рассматривается аорта - самая крупная артерия организма.
Внутренняя оболочка аорты включает эндотелий, подэндотелиальный слой и сплетение эластических волокон (в качестве внутренней эластической мембраны). С возрастом толщина интимы увеличивается.
Эндотелий аорты человека состоит из плоских эндотелиоцитов, расположенных на базальной мембране.
Подэндотелиальный слой состоит из рыхлой тонкофибриллярной соединительной ткани, богатой клетками звездчатой формы. Эти клетки, как консоли, поддерживают эндотелий. В подэндотелиальном слое встречаются отдельные продольно направленные гладкие миоциты.
Густое сплетение эластических волокон соответствует внутренней эластической мембране.
Внутренняя оболочка аорты в месте отхождения от сердца образует три карманоподобные створки - т.н. "полулунные клапаны" - единственные клапаны в артериях. Эти образования чаще называют в единственном числе - аортальный клапан.
Средняя оболочка аорты образует основную часть ее стенки, состоит из нескольких десятков эластических окончатых мембран, которые имеют вид цилиндров, вставленных друг в друга. Они связаны между собой эластическими волокнами и образуют единый эластический каркас вместе с эластическими элементами других оболочек.
Между мембранами средней оболочки аорты залегают гладкие мышечные клетки, косо расположенные по отношению к мембранам, а также фибробласты.
Окончатые эластические мембраны, эластические и коллагеновые волокна и гладкие миоциты погружены в аморфное вещество, богатое гликозаминогликанами (ГАГ). Такое строение средней оболочки делает аорту высокоэластичной и смягчает толчки крови, выбрасываемой в сосуд во время сокращения сердца, а также обеспечивает поддержание тонуса сосудистой стенки во время диастолы.
Наружная оболочка аорты относительно тонкая, не содержит наружной эластической мембраны. Построена из рыхлой волокнистой соединительной ткани с большим количеством толстых эластических и коллагеновых волокон, имеющих главным образом продольное направление. Наружная оболочка предохраняет сосуд от перерастяжения и разрывов.
Артерии мышечного типа
К артериям мышечного типа относятся преимущественно сосуды среднего и мелкого калибра, т.е. большинство артерий организма. В стенках этих артерий имеется относительно большое количество гладких мышечных клеток, что обеспечивает дополнительную нагнетающую силу их и регулирует приток крови к органам.
В состав внутренней оболочки входят эндотелий с базальной мембраной, подэндотелиальный слой и внутренняя эластическая мембрана.
Эндотелиальные клетки, расположенные на базальной мембране, вытянуты вдоль продольной оси сосуда.
Подэндотелиальный слой состоит из тонких эластических и коллагеновых волокон, преимущественно продольно направленных, а также малоспециализированных соединительнотканных клеток.
Кнаружи от подэндотелиального слоя расположена тесно связанная с ним внутренняя эластическая мембрана. В мелких артериях она очень тонкая, а в крупных артериях мышечного типа эластическая мембрана четко выражена.
Средняя оболочка артерий - наиболее толстая, содержит гладкие мышечные клетки, расположенные по пологой спирали (т.е. косоциркулярно). Между гладкими миоцитами находятся соединительнотканные клетки и волокна. Коллагеновые волокна образуют опорный каркас для гладких миоцитов.
Эластические волокна стенки артерии на границе с наружной и внутренней оболочками сливаются с эластическими мембранами. Таким образом, создается единый эластический каркас, который, с одной стороны, придает сосуду эластичность при растяжении, а с другой - упругость при сдавлении. Эластический каркас препятствует спадению артерий, что обусловливает их постоянное зияние и непрерывность в них тока крови.
Наружная оболочка включает в себя наружную эластическую мембрану и прослойку рыхлой волокнистой соединительной ткани.
Наружная эластическая мембрана состоит из продольных, густо переплетающихся эластических волокон, которые иногда приобретают вид эластической пластинки. Обычно наружная эластическая мембрана бывает тоньше внутренней эластической мембраны и не у всех артерий достаточно хорошо выражена.
По мере уменьшения диаметра артерий и их приближения к артериолам все оболочки артерий истончаются. Во внутренней оболочке резко уменьшается толщина подэндотелиального слоя и внутренней эластической мембраны. Количество мышечных клеток и эластических волокон в средней оболочке также постепенно убывает. В наружной оболочке уменьшается количество эластических волокон, исчезает наружная эластическая мембрана.
Артерии мышечно-эластического типа
По строению и функциональным особенностям артерии смешанного типа занимают промежуточное положение между сосудами мышечного и эластического типов и обладают признаками и тех и других.
3.Молочная железа. Источники развития, особенности структуры лактирующей и нелактирующей железы. Регуляция лактации.
Молочные железы (gll. mammae) по своему происхождению представляют видоизмененные кожные потовые железы.
Развитие Молочные железы закладываются у зародыша на 6—7-й неделе в виде двух уплотнений эпидермиса (т.н. «молочные линии»), тянущихся вдоль туловища. Из этих утолщений формируются так называемые «молочные точки», из которых в подлежащую мезенхиму врастают плотные эпителиальные тяжи. Затем они разветвляются на своих дистальных концах, формируют зачатки молочных желез. Несмотря на неполное развитие желез, у новорожденных (и мальчиков, и девочек) уже обнаруживается секреторная деятельность, которая продолжается обычно в течение недели и затем прекращается. С наступлением половой зрелости возникают резкие половые различия в темпах развития молочных желез. У мальчиков образование новых ходов замедляется и затем прекращается. У девочек развитие железистых трубок значительно ускоряется и к началу менструаций на молочных ходах появляются первые концевые отделы. Однако молочная железа достигает окончательного развития только при беременности в период лактации.
Строение У половозрелой женщины каждая молочная железа состоит из 15—20 отдельных железок, разделенных прослойками рыхлой соединительной и жировой ткани. Эти железы по своему строению являются сложными альвеолярными, и их выводные протоки открываются на вершине соска. Выводные протоки переходят в расширенные молочные синусы (sinus lactiferi), служащие резервуарами, в которых накапливается молоко, продуцируемое в альвеолах. молочные синусы впадают многочисленные ветвящиеся
и анастомозирующие молочные протоки (ductus lactiferi), заканчивающиеся до наступления периода лактации тонкими слепыми трубочками — альвеолярными молочными ходами (ductuli alveolares lactiferi). Они во время беременности и лактации дают начало многочисленным альвеолам. Полного развития молочная железа достигает во время беременности. Во второй половине беременности железистые клетки начинают вырабатывать секрет и незадолго до родов наступает секреция молозива (colostrum). Интенсивная секреция полноценного молока устанавливается в течение первых дней после рождения ребенка. Выработка молока происходит в альвеолах, имеющих вид округлых или слегка вытянутых пузырьков. Железистые клетки альвеол – лактоциты – соединяются с помощью замыкающих пластинок и десмосом, располагаются в один слой на базальной мембране. В лактоцитах хорошо развита гранулярная и агранулярная эндоплазматическая сеть, образованная канальцами и цистернами. В аппарате Гольджи, хорошо развитом в лактоцитах, завершаются образование и конденсация казеина, а также синтез лактозы, чему способствует наличие специального фермента — лактосинтетазы. Изменения молочных желез половозрелых женщин в течение овариально-менструального цикла. Разрастание концевых отделов отмечается за несколько дней перед овуляцией и продолжается до 20-го дня: с 22—23-го дня пролиферативные процессы прекращаются и альвеолы претерпевают обратное развитие до первых дней менструальной фазы. С 9— 10-го дня вновь начинается разрастание ацинусов, но в их клетках нет признаков секреции.
Регуляция функции молочных желез В онтогенезе зачатки молочных желез начинают интенсивно развиваться после наступления полового созревания, когда в результате значительного увеличения образования эстрогенов устанавливаются менструальные циклы и форсируется развитие вторичных признаков женского пола. Но полного развития и окончательной дифференцировки молочные железы достигают только во время беременности. С момента имплантации зародыша в эндометрий матки в дольках молочной железы разрастаются альвеолярные ходы, на концах которых формируются альвеолы. Во второй половине беременности в альвеолах начинается секреция молозива. Интенсивная секреция полноценного молока устанавливается в первые дни после рождения ребенка. Регуляция деятельности функционирующей молочной железы осуществляется двумя основными гормонами – пролактином и окситоцином. Гипофизарный пролактин (или лактотропный гормон), стимулирует железистые клетки альвеол (лактоциты) к биосинтезу молока, накопляющегося сначала в млечных ходах. Гипоталамический окситоцин стимулирует выведение молока из млечных протоков во время лактации.
Билет 42
1.Строение яйцеклетки и сперматозоида. Оплодотворение. Строение сперматозоида.
1.Головка спермия (caput spermii) имеет спереди овальное, а сбоку грушевидное очертание, то есть на свободном конце она несколько сплющена. Передняя часть головки покрыта тонким, прозрачным слоем плазмы, которая является довольно устойчивой и умеренно заостренной — так называемый perforatorium. У некоторых животных это приспособление, возможно, играет роль образования, облегчающего проникновение головки спермия в яйцеклетку во время оплодотворения. Почти все вещество головки — за исключением очень тонкого и почти невидимого цитоплазматического слоя на поверхности — состоит из ядерного вещества, очень сгущенного в этой части и интенсивно красящегося ядерными красками. Иногда в ядерном веществе головки обнаруживается шаровидная вакуоль.
2.Шейка спермия (collum spermii) представляет собой короткую, более узкую часть, образованную мягким, гомогенным плазматическим веществом (massa intermedia). Сразу же за головкой в ней располагается проксимальная (передняя) центриоль. Мягкое вещество шейки обусловливает сравнительную подвижность головки по отношению к хвосту и ее наклон под определенным, почти прямым углом. Длина шейки составляет приблизительно 0,5 мк.
3.Соединяющая (средняя) часть спермия (pars conjunctionis spermii) отграничивается от шейки проксимальной частью дистальной (задней) центриоли в виде пластинки, которая, по мнению некоторых авторов, состоит из целого ряда мелких зернышек (noduli posteriores). Соединяющая часть оканчивается кольцеподобным образованием, исходящим из дистальной части дистальной центриоли (annulus). Посередине соединяющей части проходит осевая нить хвоста, образованная тонкими фибриллами и исходящая, по мнению одних авторов, из передней центриоли, по мнению же других — из задних зернышек. Осевая нить выходит из соединяющей части через кольцеподобное образование и проходит далее по хвосту спермия. В области соединяющей части она непосредственно покрыта цитоплазматическим слоем (involucrum), вокруг которого несколько раз обвивается спиралевидное волоконце, образованное митохондриями цитоплазматического слоя (спираль). Длина соединяющей части составляет приблизительно 3,6 мк; она считается двигательным центром хвоста.
4.Хвост, или жгутик (cauda spermii) представляет собой собственно продолжение осевой нити соединяющей, средней части. Осевая нить на отрезке длиной в среднем в 20—30 микрон покрыта тонким слоем цитоплазмы (pars principialis), а ее конец, длиной приблизительно в 5 мк, обнажен и, постепенно утончаясь, заканчивается заострением (pars terminalis). Общая длина спермия колеблется от 50 до 60 мк.
Яйцеклетки позвоночных овальной формы, неподвижны, состоят из ядра и цитоплазмы. Шаровидной формы и большого размера ядро построено, как и у соматической клетки, из хроматина,
ядрышка, кариолимфы и кариолеммы. Ядро содержит гаплоидный (одинарный) набор хромосом. Яйцеклетки млекопитающих относят к гомогаметным, так как в их ядре имеется только Х-хромосома. Ядрышко крупное. Его размер и строение свидетельствуют об интенсивном синтезе рибосом и РНК.Для цитоплазмы характерна относительно высокая степень базофилии, обладающей сродством с РНК. В ней содержатся свободные рибосомы, эндоплазматическая сеть, комплекс Гольджи, митохондрии, мультивезикулярные тельца, желток. В образовании последнего активное участие принимают эндоплазматическая сеть и комплекс Гольджи. Желток, являясь цитоплазматическим включением, имеет вид гранул, 00шаров и пластин; состоит из протеинов, углеводов, фосфолипидов.
Овоциты обладают полярностью. Это объясняется неравномерным расположением желтка и других цитоплазматических структур, в связи с чем в овоците различают два полюса: апикальный и базальный.
Периферический слой цитоплазмы яйцеклетки называют кортикальным слоем (cortex - кора). Он лишен полностью желтка, содержит множество митохондрий. Функции у этого слоя - перенос питательных веществ и участие в развитии зародыша на самых ранних стадиях эмбриогенеза.
Яйцеклетки покрыты оболочками. Различают первичную, вторичную и третичную оболочки. Первичная оболочка - это плазмолемма, поэтому она присутствует всегда. Вторичная оболочка является производной фолликулярных клеток яичника. Она состоит из фолликулярных клеток и выполняет трофическую, защитную функции, препятствует полиспермии (оплодотворение яйцеклетки многими спермиями) и особенно хорошо развита у млекопитающих. Третичная оболочка овоцита формируется из материала, секретируемого клетками яйцевода. Она играет защитную, а также трофическую функции и развита у пресмыкающихся и птиц. Третичными оболочками яйцеклетки птиц являются белок, подскорлуповая и скорлуповая оболочки яйца.
Таким образом, яйцеклетки в связи с выполняемыми ими функциями обладают рядом морфологических особенностей: наличием запаса питательного материала (желтка), кортикального слоя, специальных оболочек, полярностью, генетической однородностью (Х-хромосомой).
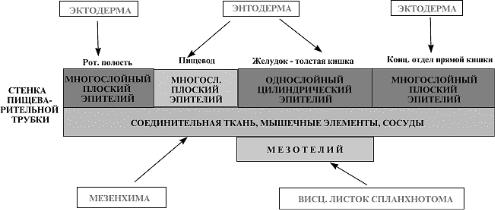
2.Пищеварительная трубка. Общий план строения стенки, источники развития. Миндалины: строение
и функции.
Общий план строения пищеварительной трубки Пищеварительная трубка в любом ее отделе
состоит из четырех оболочек:
внутренней - слизистой оболочки (tunica mucosa),
подслизистой основы (tela submucosa),
мышечной оболочки (tunica muscularis) и
наружной оболочки, которая представлена либо серозной оболочкой (tunica serosa), либо адвентициальной оболочкой (tunica adventitia).
Следует отметить, что подслизистая основа часто рассматривается в составе слизистой (и тогда мы говорим о трех оболочках в составе стенки ЖКТ). Серозная оболочка иногда рассматривается как вид адвентициальной оболочки.
|
эпителий |
слизистой |
оболочки |
- из |
эктодермы |
(в |
наружных |
отделах |
трубки) |
или |
|
из энтодермы (во внутренних отделах); |
|
|
|
|
|
|
|
||
эпителий (мезотелий) серозной оболочкииз висцерального листка спланхнотома;
остальные элементы стенки (кроме нервных сплетений)- из мезенхимы.
. Слизистая оболочка Свое название она получила в связи с тем, что поверхность ее постоянно увлажняется выделяемой железами слизью. Эта оболочка состоит, как правило, из трех пластинок:
-эпителиальной пластинки (эпителия),
-собственной пластинки слизистой (lamina propria mucosae) и
-мышечной пластинки слизистой (lamina muscularis mucosae).
Эпителий (1) в переднем и заднем отделах пищеварительной трубки — многослойный плоский, а в среднем ее отделе — однослойный призматический. По отношению к эпителию пищеварительные железы расположены либо эндоэпителиально (например, бокаловидные клетки в кишечнике), либо
экзоэпителиально: в собственной пластинке слизистой оболочки (пищевод, желудок) и в подслизистой
основе (пищевод, двенадцатиперстная кишка) или же за пределами пищеварительного канала (печень, поджелудочная железа).
Собственная пластинка слизистой оболочки (2) лежит под эпителием, отделена от него базальной мембраной и представлена рыхлой волокнистой соединительной тканью. Здесь находятся кровеносные и лимфатические сосуды, нервные элементы, скопления лимфоидной ткани. В некоторых отделах (пищевод, желудок) могут располагаться простые железы.
Мышечная пластинка слизистой оболочки (3) расположена на границе с подслизистой основой и состоит из 1-3 слоев, образованных гладкими мышечными клетками. В некоторых отделах (язык, десны) гладкие мышечные клетки отсутствуют. Рельеф слизистой оболочки на протяжении всего пищеварительного канала неоднороден. Поверхность ее может быть гладкой (губы, щеки), образовывать углубления (ямочки в желудке, крипты в кишечнике), складки (во всех отделах), ворсинки (в тонкой кишке). Рельеф слизистой зависит от мышечной пластинки слизистой, а также от степени выраженности подслизистой основы.
II. Подслизистая основа Состоит из рыхлой волокнистой соединительной ткани. Наличие подслизистой основы обеспечивает подвижность слизистой оболочки, образование складок. В подслизистой основе имеются сплетения кровеносных и лимфатических сосудов, скопления лимфоидной ткани и подслизистое нервное сплетение Мейснера (plexus nervorum submucosus). В двух отделах ЖКТ - пищеводе и двенадцатиперстной кишке - в подслизистой основе расположены железы.
III. Мышечная оболочка Состоит, как правило, из двух слоев – наружного продольного и внутреннего циркулярного. В переднем и заднем отделах пищеварительного канала мышечная ткань преимущественно поперечнополосатая, а в среднем (большем) отделе – гладкая. Мышечные слои разделены соединительной тканью, в которой находятся кровеносные и лимфатические сосуды и межмышечное нервное сплетение Ауэрбаха (plexus nervorum intermuscularis s. myenteric). Сокращения мышечной оболочки способствуют перемешиванию и продвижению пищи в процессе пищеварения.
IV. Наружная оболочка Большая часть пищеварительной трубки покрыта серозной оболочкой –
висцеральным листком брюшины. Брюшина состоит из соединительнотканной основы (т.е. собственно адвентициальной оболочки), в которой расположены сосуды и нервные элементы, и покрыта однослойным плоским эпителием – мезотелием. Повреждение мезотелия приводит к формированию спаек – т.е. срастанию подлежащей соединительной ткани соседних органов и нарушению их подвижности.
Лимфоэпителиальное глоточное кольцо Пирогова-Вальдейера. Миндалины.
На границе ротовой полости и глотки в слизистой оболочке располагаются большие скопления лимфоидной ткани. В совокупности они образуют лимфоэпителиальное глоточное кольцо, окружающее вход в дыхательные и пищеварительные пути. Наиболее крупные скопления этого кольца носят название миндалин. По месту их расположения различают небные миндалины, глоточную миндалину, язычную миндалину. Кроме перечисленных миндалин, в слизистой оболочке переднего отдела пищеварительной трубки существует ряд скоплений лимфоидной ткани, из которых наиболее крупными являются скопления в области слуховых труб – трубные миндалины и в желудочке гортани – гортанные миндалины. Миндалины выполняют в организме важную защитную функцию, обезвреживая микробы, постоянно попадающие из внешней среды в организм через носовые и ротовое отверстия. Наряду с другими органами, содержащими лимфоидную ткань, они обеспечивают образование лимфоцитов, участвующих в реакциях гуморального и клеточного иммунитета.
Небные миндалины закладываются на 9-й неделе эмбриогенеза в виде углубления псевдомногослойного реснитчатого эпителия латеральной стенки глотки, под которым лежат компактно расположенные мезенхимные клетки и многочисленные кровеносные сосуды. На 11—12-й неделе формируется тонзиллярный синус, эпителий которого перестраивается в многослойный плоский, а из мезенхимы дифференцируется ретикулярная ткань; появляются сосуды, в том числе посткапиллярные венулы с высокими эндотелиоцитами. Происходит заселение органа лимфоцитами. На 14-й неделе среди лимфоцитов определяются главным образом Т-лимфоциты (21 %) и немного В-лимфоцитов (1 %). На 17— 18-й неделе появляются первые лимфатические узелки. К 19- й неделе содержание Т-лимфоцитов возрастает до 60 %, а В-лимфоцитов — до 3 %. Рост эпителия сопровождается формированием в эпителиальных тяжах пробок из ороговевающих клеток.
Строение.
Небные миндалины во взрослом организме представлены двумя телами овальной формы, расположенными по обеим сторонам глотки между небными дужками. Каждая миндалина состоит из нескольких складок слизистой оболочки, в собственной пластинке которой расположены многочисленные лимфатические узелки (noduli lymphathici). От поверхности миндалины в глубь органа отходят 10–20 крипт (criptae tonsillares), которые разветвляются и образуют вторичные крипты. Слизистая оболочка покрыта многослойным плоским неороговевающим эпителием. Во многих местах, особенно в криптах, эпителий часто бывает инфильтрирован (заселен) лимфоцитами и гранулоцитами. Лейкоциты, проникающие в толщу эпителия, обычно в большем или меньшем количестве выходят на его поверхность и мигрируют навстречу бактериям, попадающим в полость рта вместе с пищей и воздухом. Микробы в миндалине активно фагоцитируются лейкоцитами и макрофагами, при этом часть лейкоцитов погибает. Под влиянием микробов