

06 Загальна ембріологія. Ембріональний розвиток людини. Статеві клітини. Запліднення
.pdfЗАГАЛЬНА ЕМБРІОЛОГІЯ. ЕМБРІОНАЛЬНИЙ РОЗВИТОК ЛЮДИНИ. СТАТЕВІ КЛІТИНИ. ЗАПЛІДНЕННЯ.
Користуючись лекціями (на web-сторінці кафедри розміщені презентації та текст лекцій), підручниками, додатковою літературою та іншими джерелами, студенти повинні підготувати такі теоретичні питання:
1.Поняття про філо- і онтогенез. Періоди ембріогенезу людини.
2.Прогенез. Особливості овогенезу.
3.Мікрота субмікроскопічна характеристика чоловічих статевих клітин.
4.Типи яйцеклітин. Тонка будова та функція яйцеклітини людини.
5.Запліднення, його етапи та біологічна роль.
6.Поняття про екстракорпоральне запліднення та його значення.
7.Синкаріон. Зигота.
Ем б рі о л о гі я — наука про розвиток зародка. Під зародковим або ембріональним розуміють той ранній період індивідуального розвитку організму, який триває від моменту запліднення до народження на світ або до вилуплення з яйцевих оболонок (А.Г.Кнорре, 1982).
І н д и в і д у а л ь н и й |
р о з в и т о к |
або онтогенез |
— це розвиток |
||
організму, який триває з моменту зачаття до його природної смерті. |
|||||
Онто генез тісно пов'язаний з фі л о ге не зо м — історією розвитку |
|||||
предків даної істоти (історичний розвиток виду). |
|
|
|||
|
|
||||
Предметом вивчення ембріології є також п р о г е н е з |
(передзародковий |
||||
період) — період формування статевих |
клітин |
(спермато- і овогенез) і |
|||
п о с т е м б р і о н а л ь н и й |
(післязародковий) |
період |
індивідуального |
||
розвитку. |
|
|
|
|
|
|
|
|
|
|
|
Ембріологія вивчає як загальні закономірності індивідуального розвитку, які виявляються в усіх багатоклітинних тваринних організмів, так і окремі особливості індивідуального розвитку, характерні для представників окремих типів, класів та видів тварин.
В зв'язку з цим розрізняють загальну та спеціальну ембріологію.
З а г а л ь н а ембріологія вивчає найбільш загальні і найбільш

широкі |
закономірності |
онтогенезу |
певного |
виду |
тварин. |
С п е ц і а л ь н а ембріологія вивчає |
особливості |
індивідуального |
|||
розвитку окремих груп тварин.
Значення ембріології перш за все полягає в тому, щоб сформувати у студентів загальні біологічні уявлення про онтогенез та створити теоретичну основу для вивчення педіатрії, акушерства і гінекології та тератології.
МЕТОДИ ДОСЛІДЖЕННЯ
1.Найбільш простим є метод спостереження і опис розвитку зародка у тих чи інших тварин або людини. На основі цього виникла
оп и с о в а е м б р і о л о г і я .
2.Значно пізніше в дослідженнях почали застосовувати метод
порівняння, який створив п о р і в н я л ь н у е м б р і о л о г і ю . За допомогою цього методу К.М.Бер (1828 р.) відкрив закон подібності зародків.
3. Поступове нагромадження детальних і точних відомостей про ті структурні перетворення, які відбуваються з зародком у процесі його розвитку, підготували грунт для е в о л ю ц і й н о ї е м б р і о л о г і ї , фундаторами якої є О.Ковалевський та I.Мечников. Розвиток еволюційної ембріології привів до відкриття біогенетичного закону Е.Геккелем, Ф.Мюлером.
4. Е к с п е р и м е н т а л ь н а е м б р і о л о г і я виникла внаслідок розвитку та удосконалення експериментальної техніки. Засновниками цього методу є В.Ру, Г.Дріш і Г.Шпеман. Експериментальна ембріологія намагається встановити контролюючі та регулюючі механізми розвитку.
Блискучі успіхи молекулярної біології дали поштовх до розвитку
особливого розділу, який дістав назву б і о х і м і ч н а |
е м б р і о л о г і я . |
Т е р а т о л о г і я — це розділ ембріології, |
який вивчає вади |
розвитку. |
|

ТЕОРІЇ ЕМБРІОЛОГІЇ
Проблема розвитку людини до її народження на світ завжди викликала надзвичайну цікавість. "Звідкіля я взявся ?" — одне із перших запитань, яке ставить дитина. З іменами Гіпократа (460-377 pp. до н.е.) і Арістотеля (384-322 pp. до н.е.) пов'язано створення перших наукових уявлень з ембріології.
Згідно теорії Гіпократа плід утворюється через змішування чоловічого сім'я і жіночої статевої клітини. Він вважав, що всі члени організму виникають одночасно і незалежно один від одного. Ця ідея знову відродилась і стала домінуючою на протязі 17-18 століть і отримала назву префо рмі зму .
Згідно цієї теорії кожен зародок в мініатюрі з самого початку є уже цілком сформованим, має всі частини тіла і йому залишається тільки рости. Серед преформістів було дві течії: а н і м а л і с т и і о ві ст и . Анімалісти або сперматики вважали, що зародок преформований у сперматозоїді, який вигодовується в яйці. Овісти думали, що зародок у мініатюрі знаходиться в яйцеклітині, а сперма лише стимулює його ріст.
Арістотель вперше заклав підвалини теорії е п і г е н е з у , згідно якої частини зародка не закладені в невидимо малих розмірах, а розвиваються і ростуть в певній послідовності. Гарвей цю тезу доповнив віталістичним поглядом про те, що матеріал материнського зачатка набуває форми майбутнього організму під впливом нематеріальних сил — "ентелехій", які закладені в сім'ї батька. К.Вольф у віці 26 р. в своїй докторській дисертації блискуче обгрунтував концепцію епігенезу. Ідея розвитку шляхом прогресивного росту і диференціації швидко витіснила теорію преформізму. Однак, боротьба теорій преформізму і епігенезу стала особливо запеклою в кінці XIX століття у зв'язку з розвитком експериментальної ембріології і, так званої механіки розвитку, яка претендувала на каузальне пояснення онтогенезу. Встановлення того факту, що дорослий організм цілком складається із клітин і продуктів їхньої життєдіяльності, проклало шлях до відкриття основного положення ембріології: організм будь-якої нової істоти розвивається із однієї клітини, яка утворилась у результаті об'єднання батьківських і материнських статевих клітин при заплідненні.

Разом із тим, успіхи молекулярної біології та генетики в другій половині XX століття доказали наявність в статевих клітинах детермінованих хімічних і біологічних структур (молекул ДНК), які визначають відтворення основних видів та індивідуальних особливостей організму, який розвивається.
Обидві, довгий час конкуруючі теорії (преформізму та епігенезу), можуть бути використані для утвердження генетичної теорії розвитку.
Статеві клітини не мають готових структур для майбутнього зародка, але мають постійний набір генів — генетичну програму для конкретного виду тварин, включаючи особливості для кожного індивідууму. Одночасно в процесі ембріогенезу виникають структури від молекулярного до органного рівнів, яких немає в статевих клітинах, що підтверджує деякі положення епігенезу. Отже, статевим клітинам передаються за спадком не готові структури, а інструкція для їх виробництва. Ця інструкція не "ентелехія", а програма специфічних синтезів, закодованих в молекулах ДНК.
В даний час процес розвитку розглядається як реалізація тих можливостей, які закладені в спадковому коді ДНК статевих клітин. Хоча, для здійснення цих процесів розвитку необхідна наявність певних умов середовища, вони незмінно реалізуються шляхом самоди-ференціювання на основі тих видовоспецифічних закономірностей, які були філогенетично напрацьовані і закріплені впродовж еволюції органічного світу.
Основні процеси ембріогенезу
Із заплідненої яйцеклітини (зиготи) розвивається багатоклітинний організм. В зиготі гени репресовані (неактивні) і знаходяться у вигляді гетерохроматину.
Г е т е р о х р о м а т и н — це репресований генетичний матеріал (ДНК
хромосом зв'язана з білками гістонами). Початок розвитку характеризується е к с п р е с і є ю (активацією) певних груп генів, тобто звільнення частин ДНК від асоційованих з нею гістонів. Еух р о м а т и н — це експресовані (функціонуючі) гени. Першими дере-пресуються гени, які обумовлюють збільшення кількості клітин (проліферацію) і регулюють загальний метаболізм клітин. На стадії гаструляції активуються першими тканинноспецифічні гени. В

процесі органо- і гістогенезу, включаються інші гени, які регулюють більш специфічні функції диференційованих клітин.
В зиготі закладена можливість формування цілого організму. Окремі клітини, які утворюються внаслідок декількох перших поділів після запліднення, зберігають цю здатність. Такі клітини називаються т о т и п о т е н т н и м и . По мірі розвитку клітини поступово втрачають свою здатність до утворення клітин всіх типів, які є в дорослому організмі. Скорочення можливостей вибору (втрата тотипотентності) клітини, яка розвивається, називається д е т е р м і н а ц і є ю . Детермінація — процес, в результаті якого група клітин вибирає тільки один із багатьох можливих шляхів розвитку. В основі детермінації лежить експресія генів, в результаті яких дана клітина (або група клітин) стає спеціалізованою клітиною певного типу.
Зовнішнім проявом детермінації є диференціація. Якщо детермінація супроводжується поступовим обмеженням потенцій клітин зародка до розвитку в різних напрямках, то під диференціцією розуміють морфологічну чи функціональну експресію тієї частини геному, яка залишилась в розпорядженні даної клітини або групи клітин. В результаті диференціації, клітини набувають деяких нових властивостей (спеціалізуються), але одночасно втрачають частину своїх попередніх можливостей. З біохімічної точки зору диференціація — процес, за допомогою якого дана клітина вибирає для себе один або декілька спеціалізованих шляхів синтезу (наприклад: еритроцити синтезують гемоглобін, а клітини кришталика — білок кришталін). Функціональну диференціацію розглядають як розвиток здатності до скорочення у міоцитах, або провідності у нейроцитах.
З морфологічної точки зору кінцева диференціація виражається в утворенні багаточисельних спеціалізованих клітин і структур. Отже, різниця між клітинами, які мають однаковий набір генів, визначається диференційованою репресією та експресією генів. Різні клітинні типи експресують різні гени.
У зв'язку з тим, що синтез білків диктується експресією генів, то гени діляться на дві категорії. Гени першої — функціонують в усіх клітинах

організму, визначаючи їх основний метаболізм і нечутливі до безпосередніх впливів, які викликають диференціацію. Гени другої категорії керують синтезом тих білків, які визначають різницю між клітинами в період розвитку (тобто їхню спеціалізацію) і реагують на чинники, які викликають диференціацію (індуктори).
Виникає запитання, а хто здійснює експресію одних і репресію інших генів? При дробленні зиготи дочірні клітини отримують ідентичні набори хромосом (46 для людини), однак, цитоплазма не завжди ділиться між дочірними клітинами рівномірно ні за кількістю, ні за якістю. Встановлено, що нерівномірний розподіл різних ділянок цитоплазми заплідненого яйця між дочірними клітинами, зумовлює диференціацію останніх у клітини різного типу. Отже, цитоплазма приводить у дію механізм, який здійснює вплив на ядро для включення (експресії) одних і виключення (репресії) інших генів.
Суттєвою рисою диференціації є те, що диференціація клітин, як правило, відбувається після їхньої проліферації. Клітини, які швидко розмножуються, є низькодиференційованими (наприклад: мезенхімні клітини). Навпаки, високодиференційовані клітини, зазвичай, втрачають здатність до проліферації (наприклад: нейроцити, кардіоміоцити, еритроцити).
Диференціація незворотня і здійснюється в одному напрямку — від менш диференційованої до більш диференційованої структури.
М о р ф о г е н е з — втілення плану просторової організації зародка. Морфогенез здійснюється при реалізації різних процесів: росту, індукції, направленої міграції клітин, міжклітинної взаємодії, смерті клітин.
Р і с т — збільшення маси і лінійних розмірів за рахунок зростання кількості клітин, морфофункціональних одиниць органів, власне органів, систем органів. Збільшення маси без поділу клітин спостерігається при гіпертрофії в нормальних (гіпертрофія міометрію при вагітності) і патологічних умовах. В організмі виробляються багаточисельні гуморальні чинники (фактори росту), які стимулюють ріст і проліферацію різних типів клітин.
Конкретні типи клітин, які утворились внаслідок детермінації, формують тканини. Процес, в результаті якого окремі тканини в ході диференціації

набувають характерного для них вигляду, називається г і с т о г е н е з о м . Із клітин різної тканинної приналежності формуються органи. В органогенезі важливе значення має процес і н д у к ц і ї . Під індукцією розуміють вплив однієї тканини зародка (індуктора) на іншу тканину — ціль, в результаті чого відбувається утворення якісно нової структури. Індуктор може давати інструкцію тканині — цілі для диференціації в конкретну структуру або дозволяти її. Прикладом первинної ембріональної індукції є процес формування нервової системи із дорсальної ектодерми під впливом індуктора — хордомезодерми. Відкриття організатора і феномена первинної ембріональної індукції є результатом двадцятирічних експериментально-ембріологічних досліджень Ганса Шпемана, який за це відкриття у 1935 році став лауреатом Нобелівської премії. Усі наступні індукційні процеси, що відбуваються в зародку, називаються вторинними індукціями. В процесі розвитку зародка відбувається неодноразові переміщення, або м і г р а ц і я окремих або груп клітин із однієї частини в іншу. Нерідко ембріональні клітини виникають і розмножуються в одному місці, а диференціюються і функціонують в іншому (наприклад: ганобласти утворюються в жовтковому мішку, а мігрують
всім'яники і яєчник; клітини нервового гребеня виселяються і диференціюються
вмелаьгацити епідермісу, хромафінні клітини надниркових залоз, нейроцити вегетативних та спинномозкових вузлів; клітини дерматома мезодерми виселяються під покривний епідерміс і формують сполучнотканинну основу шкіри).
Природу |
клітинних взаємодій в морфогенезі пояснює концепція |
п о з и ц і й н о ї |
і н ф о р м а ц і ї . При розвитку загальний план будови тіла |
визначається дуже рано. Пізніше, на протязі всього періоду формування органу або цілого організму, деталі морфогенезу уточнюються за допомогою сигналів позиційної інформації. Згідно названої концепції, клітина знає своє місце в координатній системі зачатку органу і диференціюється у відповідності з цим положенням. Позиційну інформацію клітина отримує від інших клітин. Більше того, клітина досягає кінцевої диференціації за умови своєчасного отримання нею серії послідовних сигналів позиційної інформації. Ділянка, в

межах якої ефективно діють сигнали |
позиційної інформації, називається |
|
|
протязі низки наступних поділів, |
|
м о р ф о г е н е т и ч н и м п о л е м |
. На |
|
клітини морфогенетичного поля зберігають пам'ять про своє початкове призначення.
Постійна активність гомейозисних генів визначає в клітині пам'ять про позиційну інформацію. Гомейозисні гени — родина споріднених генів, які містять гомеобокс і визначають форму тіла. У ссавців ця родина представлена 38 генами, які згруповані в 4 комплекси — НОХА, НОХВ, НОХС і НОХД (Human Gene Mapping Workshops — робоча група картирування генів людини). Ці групи генів розміщені в 2, 7, 12 і 17 хромосомах. Експресія цих генів відбувається в ембріогенезі і визначає організацію загального плану тіла. Експресія цих генів контролює розділення тіла ембріона за координатами на морфогенетичні зони. Транскрипти гомейозисних генів є присутні в головному і спинному мозку, в нирках і серці з 5-го до 9-го тижня розвитку.
С м е р т ь клітин — необхідний компонент ембріонального ровитку.
Природна (генетична) смерть клітин на ранніх етапах онтогенезу реалізується шляхом а п о п т о з у . Наприклад: від 25 % до 75 % загальної популяції нейробластів гине на певних етапах розвитку нервової системи.
СТАТЕВІ КЛІТИНИ АБО ГАМЕТИ
Статеві клітини на відміну від соматичних мають гаплоїдний набір хромосом. Всі хромосоми гамет, за винятком однієї статевої, називаються аутосомами. В чоловічих статевих клітинах людини та ссавців статеві хромосоми X або Y, а в жіночих — тільки X. Статеві клітини на відміну від соматичних мають здатність до асиміляції (злиття) і якщо вона не відбулася, то чоловічі і жіночі гамети гинуть.
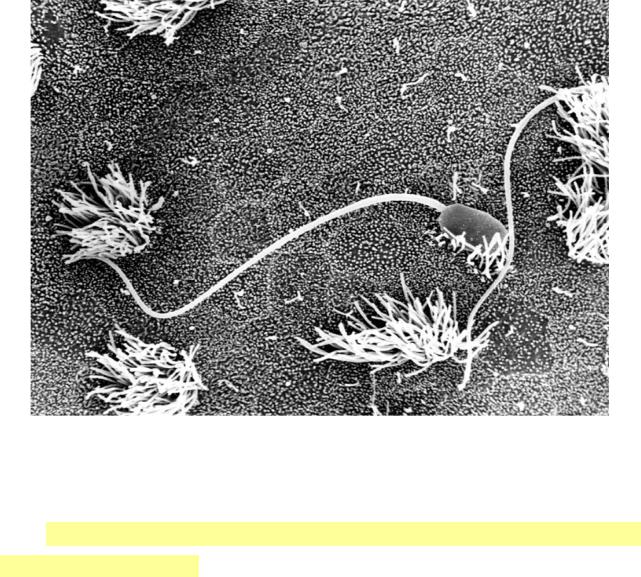
Ч о л о в і ч і с т а т е в і к л і т и н и — сперматозоїди, або спермії, розвиваються в дуже великій кількості. В спермі людини, яка виділяється при еякуляції, міститься від декількох мільйонів сперматозоїдів, у риб і птахів — до декількох мільярдів. Розміри сперматозоїдів коливаються у широких
межах: від 20 мкм у кролика до 5000 мкм у тритона. Сперматозоїди мають здатність до активного руху. Форма їх може бути різною. У хребетних сперматозоїди мають джгутикову форму ( мають хвіст, що по своїй суті є органелою спеціального призначення – джгутиком).
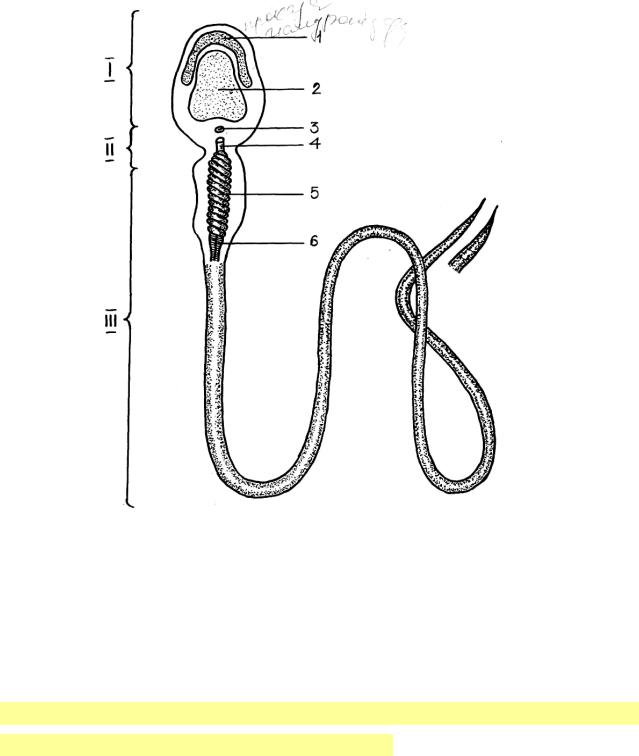
Будова с п е р м а т о з о ї д а . В ньому розрізняють головку, шийку і
хвіст.
І – голівка: 1- акросома, 2 – ядро, ІІ – шийка: 3- проксимальна центріоля, ІІІ – хвіст: 5 – мітохондріальна піхва, осьова нитка
|
Г о л о в к а с п е р м а т о з о ї д а |
побудована із невеликого щільного |
|
|
|
ядра з гаплоїдним набором хромосом |
та вузькою смужкою цитоплазми. В |
|
передній частині є чохлик, в якому розміщується акросома. Чохлик і акросома є похідними комплексу Гольджі. В акросомі локалізуються видоспецифічні ферменти гіалуронідаза і протеази, які здатні розщеплювати
оболонки, якими покрита яйцеклітина.
Апікальна частина голівки спермія, в якій знаходиться акросома і яка має дещо загострену форму називається перфораторіум.
В шийці сперматозоїда розміщуються проксимальна і дистальна центріолі, від останньої починається осьова нитка, яка продовжується у хвостовий відділ.
Сперматозоїди на поверхні війчастого епітелію сім’явиносних шляхів.
Х в о с т о в и й відділ сперматозоїда складається із початкового, головного і кінцевого. В початковому відділі довкола осьової нитки локалізуються мітохондрії. Головний відділ хвоста побудований із центральної пари мікротрубочок з осьовою ниткою і периферійно локалізованими дев'ятьма парами мікротрубочок, покритий вузькою смужкою цитоплазми, яка зникає у кінцевому відділі.
Сперматозоїди тварин різняться між собою, головним чином, за будовою головки. У амфібій головка сперматозоїда довга і загострена, у ссавців — сплющена.