

ogurtsov_a_n_vvedenie_v_biofiziku_fizicheskie_osnovy_biotekh
.pdfКаждая из нитей двойной спирали состоит из пентоз и фосфатных групп, нуклеиновые основания ориентированы внутрь двойной спирали. В ДНК обычно используются четыре нуклеиновых основания: аденин, гуанин, цитозин и тимин. Аденин образует пару с тимином, формируя две водородные связи. Гуанин спаривается с цитозином, формируя три водородные связи. В РНК тимин заменён урацилом. Нуклеиновые основания имеют подобные химические свойства и различаются, главным образом, ориентацией акцепторов и доноров водородных связей по периметру оснований. Кроме двух основных уотсон-криковских пар возможны и другие типы спаривания, кроме того, в специальных случаях для расширения диапазона возможных типов спаривания нуклеотидов используются также и модифицированные азотистые основания.
Главная функция нуклеиновых кислот – хранение и обработка генетической информации. В то же время, рибосома, которая является наиболее сложной и важной молекулярной машиной в клетке, состоит преимущественно из РНК. Отсепарированные и высушенные рибонуклеиновые кислоты имеют фибриллярную форму и похожи на волокна хлопка.
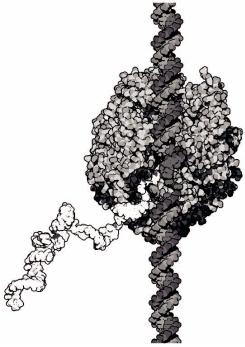
Основная (В-форма) ДНК, несмотря на достаточно высокую стабильность структуры, обеспечиваемую множественными водородными и гидрофобными взаимодействиями, обладает значительной гибкостью. Это свойство обеспечивает необходимую модификацию структуры ДНК при связывании белков со специфическими участками ДНК. В ДНК, в
отличии от белковых α- спиралей, нет водородных связей, параллельных основной оси. Это свойство позволяет молекуле ДНК изгибаться при объединении с белками,управляющими эволюцией ДНК (рисунок 66).
Высокая гибкость ДНК обеспечивает плотную упаковку ДНК в нуклеосомы хроматина – ДНК –белкового комплекса, из которого формируются хромосомы эукариот. В ходе процессов репликации и транскрипции ДНК две нити двойной спирали должны разъединяться с тем, чтобы позволить "внутренним" краям оснований спариваться с соответствующими комплементарными основаниями, формируя новую полинуклеотидную цепь.
200
Рисунок 66 –Изгиб ДНК при связывании с С-доменом ТАТА-связывающего белка (TATAbox-binding protein (TBP))
Процесс раскручивания двойной спирали и разъединения двух спиралей называется денатурацией (или плавлением) ДНК. Денатурация может быть индуцирована искусственно in vitro нагреванием раствора, содержащего ДНК. При повышении температуры усиливается тепловое движение в молекулах и разрушаются слабые нековалентные связи, которые формируют двойную спираль. Спирали разъединяются и расходятся вследствие электростатического отталкивания между отрицательно заряженными сахарофосфатными остовами. Вблизи температуры денатурации небольшое увеличение температуры раствора вызывает резкое, практически одновременное, разрушение множественных слабых нековалентных взаимодействий( которые удерживают двойную спираль) вдоль всей длины ДНК, что проявляется в резком изменении ультрафиолетового поглощения раствором (рисунок 67(а)).
Температура плавления, Tm , при которой разъединяются две спирали ДНК, определяется несколькими факторами. Молекулы, которые содержат большее количество C≡G пар плавятся при более высокой температуре, поскольку пары C≡G, имеющие три водородные связи, более стабильны, чем пары А=Т, в которых только две водородные связи. Таким образом, по температуре плавления можно оценить концентрацию
201
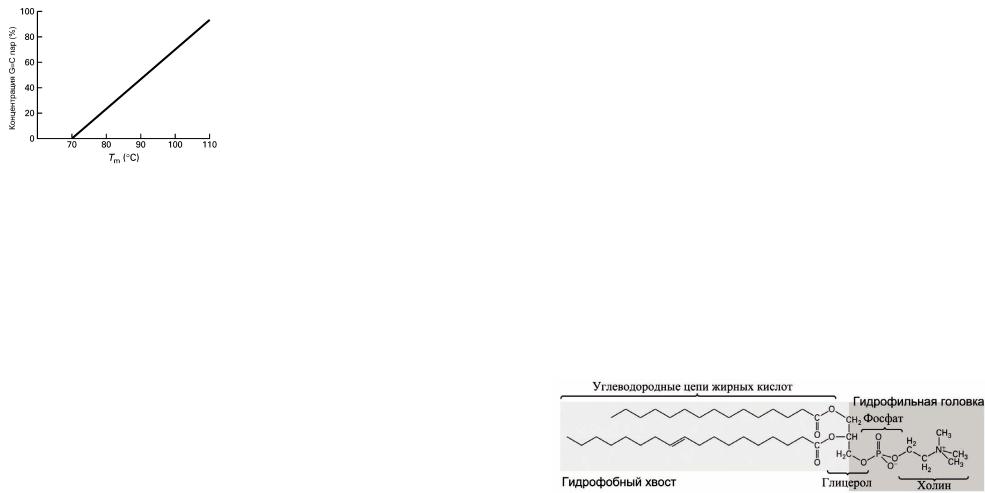
C≡G пар (рисунок 67(б)).Справедливо эмпирическое соотношение
Tm =TAT + (TCG −TAT )x ,
где x –доляC ≡G пар, TAT ≈ 70°С и TCG ≈110°С –температуры плавления полинуклеотидов, состоящих исключительно из А=Т или C≡G пар при значениях x = 0 и x =1,соответственно.
|
а |
б |
Рисунок |
67 – Температура денатурации ДНК: а –поглощение ультрафиолета, |
|
б –зависимость |
концентрацииG |
≡C парот Tm |
Концентрация ионов в растворе также влияет на Tm , поскольку отрицательно заряженные фосфатные группы обеих спиралей экранированы положительными ионами. Если снизить концентрацию положительных ионов в растворе, то уменьшается и степень экранирования. Это увеличивает силы отталкивания между сахарофосфатными остовами и снижает Tm . Наконец, резкое изменение рН денатурирует ДНК даже при низких температурах. При низких значениях рН (кислая среда) нуклеиновые основания протонируются и, следовательно, приобретают положительный заряд, отталкивая друг друга. При высоких значениях рН (щелочная среда) основания теряют протоны, становятся отрицательно заряженными и опять-таки отталкиваются друг от друга.
Одноцепочечные молекулы ДНК, которые получаются в результате денатурации, образуют случайным образом сформированные клубки без определенной структуры. Понижение температуры, увеличение ионной
концентрации или нейтрализация рН вызывают самопроизвольную ренатурацию двух цепей ДНК в двойную спираль. При этом, однако,
важно, что две некомплементарные цепи ДНК не ренатурируют, а
остаются в виде хаотичных клубков. Еще более важно, что они не мешают двум комплементарным цепям ДНК найти друг друга в растворе и ренатурировать.
Процессы денатурации и ренатурации ДНК являются основами гибридизации ДНК –методики , которая используется для определения взаимного соответствия двух ДНК-образцов, и для детектирования и изолирования специфических молекул ДНК в растворе, содержащем различные ДНК последовательности.
6.3. ЛИПИДЫ ИПОЛИСАХАРИДЫ
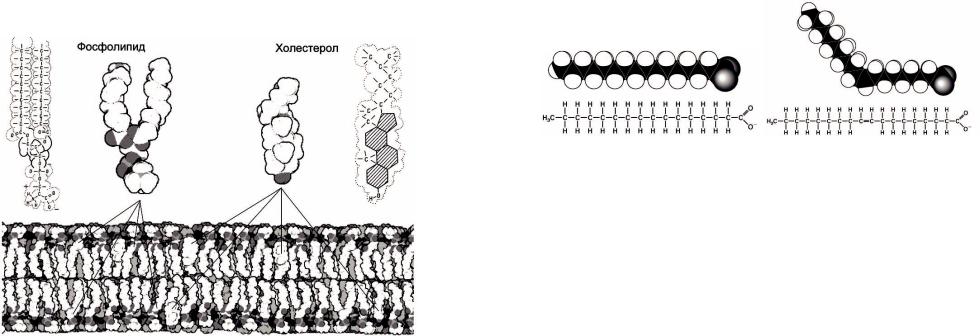
Примечательно, что самые большие пространственные структуры, которые формируются в клетке, образованы не из макромолекул, таких, как белки или нуклеиновые кислоты, а агрегацией липидов в биомембраны– обособленную жидкую фазу. Липиды, которые используются клеткой, устроены так, чтобы осуществлялась самопроизвольная агрегация, в результате которой в клетке формируется клеточная инфраструктура. В молекуле липидов объединены два противоположных химических качества. Они одновременно содержат и полярную или даже заряженную гидрофильную группу атомов, которая стремится раствориться в воде, и одну или несколько гидрофобных углеводородных цепей, которые выталкиваются из воды (рисунок 68). Такие молекулы называются амфифильными.
Рисунок 68 – Фосфатидихол– ипичный |
фосфолипид |
202 |
203 |
Двойственная природа липидных молекул приводит к их |
ненасыщенные углеродуглеродные связи, то в таких местах образуются |
самоорганизации в мембранные структуры, в которых заряженные (или |
жесткие кинки (рисунок 70(б)). |
полярные) головки обращены в сторону воды, а углеводородные хвосты |
|
упакованы внутри мембраны( рисунок 69). |
|
Рисунок 69 – Молекулы фосфолипидов (фосфатидилсерин) и холестерола из которых происходит самосборка биомембраны. Темно-серым цветом обозначены полярные участкимолекул
Наиболее распространенные типы липидов в природе это фосфолипиды и гликолипиды. В основе их конструкции лежит молекула глицерола, имеющая три гидроксильные группы, вместо которых могут быть присоединены три другие группы. Две из них, как правило, это жирные кислоты, присоединенные к глицеролу через карбоксильную группу. Углеводородные хвосты этих жирных кислот имеют 16–24 атома углерода. Вместо третьей гидроксильной группы к глицеролу присоединяется фосфатная группа или другая полярная (или заряженная) группа. Если жирнокислотные хвосты имеют в своем составе
204
|
а |
б |
Рисунок 70 – Влияние |
двойной С=С связи на форму жирной кислоты: |
|
а–пальмитат |
(ионизированная форма пальмитиновой кислоты), б – олеат (ионизиро- |
|
ваннаяформа |
олеиновойкислоты |
) |
Жирнокислотные хвосты с кинками гораздо хуже упаковываются в упорядоченную структуру, и, поэтому, биомембрана с ненасыщенными углеводородными цепями в липидах имеет более низкую температуру фазового перехода из физиологического жидкокристаллического в замороженное гель-состояние, чем у биомембраны с насыщенными С–С связями в липидах.
Холестерол и другие стеролы устроены иначе. Они состоят из нескольких жестко связанных гидрофобных углеводородных колец протяженностью такой же, как и углеводородные хвосты фосфолипидов. Гидроксил на одном из концов обеспечивает гидрофильность, ориентируя холестерол в мембране. Холестерол добавляется к мембранам в разных пропорциях для того, чтобы модифицировать свойства биомембран. Поскольку молекула холестерола является жесткой, то добавление холестерола ингибирует( затрудняет) движение соседних липидов, увеличивая тем самым вязкость мембраны и делая ее менее проницаемой для малых молекул.
Липиды используются клетками для формирования мембранных структур клетки. Мембраны непроницаемы для ионов и больших молекул,
205
начиная с сахаров и полипептидов, но легко проницаемы для молекул воды, кислорода, азота. Примечательно, что молекулы, содержащие много атомов углерода, достаточно легко проникают через мембраны. Именно поэтому алкоголь легко распространяется по организму, пересекая все барьеры.
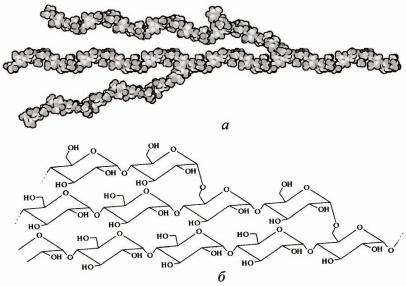
Полисахариды. Полисахариды являются наиболее гетерогенными структурами из четырех типов молекулярных образований, которые используются в природе.
Моносахариды – мономеры полисахаридов – имеют множество гидроксильных групп и могут соединяться друг с другом, образуя гликозидную связь между гидроксильной группой одного моносахарида и альдегидной или кетогруппой другого моносахарида, благодаря чему происходит полимеризация моносахаридов в полисахарид. При этом возможно образование бесчисленного множества структур.
В природе множество различных линейных и разветвленных структур полисахаридов синтезируется для различных нужд (рисунок 71).
Рисунок 71 – Полисахариды: а– схемамолекулы полисахарида, б – схема связей вмолекуле гликогена
Например, |
объединение молекул глюкозы посредством |
(β1 →4) |
связи образует |
длинные линейные молекулы целлюлозы, из |
которых, |
например, образованы волокна хлопка. Однако же, если использовать несколько другой тип связи, а именно (α1→4) связь, полисахаридные молекулы образуют спирали, которые образуют рыхлые гранулы
крахмала.
Широко используется также ветвление полисахаридов. Гликоген является примером такого использования ветвления –он представляет собой дендромер, состоящий из постоянно разветвляющихся полисахаридных цепей глюкозы. Гликоген используется для хранения глюкозы, поэтому его древовидная (дендромерная) компактная форма обеспечивает, тем не менее, свободный доступ ко многим свободным концам ветвей для отсоединения, по мере необходимости, индивидуальных моносахаров.
Большое количество гидроксильных групп в полисахаридах образуют водородные связи с другими донорами или акцепторами, обеспечивая существование двух важных типов структур в живых организмах.
Водном случае, отдельные полисахаридные цепи ассоциируются с большим количеством молекул воды, образуя клейкий гель. В таком виде углеводы покрывают большинство наших клеток, образуя их клейкую защитную оболочку. Гликопротеины слизи дают представление о внешнем виде и свойствах такого образования.
Вдругом случае, углеводные цепи прочно ассоциируются друг с другом посредством сети водородных связей, образуя прочные волокна, внутри которых практически нет воды. В таком виде полисахариды используются для формирования макроскопической инфраструктуры клеток и межклеточных образований и для запасания энергии. Некоторые из наиболее впечатляющих биологических структур, созданных природой, включая прочные стволы гигантских деревьев и жесткие водонепроницаемые панцири членистоногих, обязаны своей силой и прочностью полисахаридам.
206 |
207 |
6.4. ЭВОЛЮЦИОННАЯСПЕЦИФИКА |
СТРОЕНИЯИ ФУНКЦИОНИРОВАНИЯ |
БИОСТРУКТУР
Если бы кто-нибудь поставил перед нами задачу сконструировать живую клетку, мы бы, вероятно, не пошли бы тем наиболее экономным и бережливым путем, который использовала природа. Рассмотрим, к примеру, компьютер. Он состоит из микроскопически синтезированных кремниевых микросхем, пластикового корпуса, металлических проводов, соединяющих устройства, флуоресцентных веществ, нанесенных на стекло и бомбардируемых электронами, чтобы получить свечение. Все эти компоненты произведены используя различные технологические процессы, различные базовые принципы,часто в разных частях света.
Клетки более однородны– они используют всего несколько технологий синтеза и только нескольких базовых типов макромолекул для того, чтобы построить множество различных бионаноструктур. Это является одновременно и ценным качеством, и ограничивающим фактором. Биологические молекулы имеют свои ограничения – им необходимо водное окружение с соответствующей температурой, рН и концентрацией солей. Причина такого ограниченного количества использованных "базовых технологий" –эволюционный процесс путем естественного отбора. Эволюция наложила жесткие граничные условия на то, каким образом биологические молекулы изменялись в ходе эволюции, всячески стимулируя модификацию уже существующих структур и как можно реже "изобретая" действительно новые молекулярные решения. Эволюционный процесс происходит в процессе передачи генетической информации от поколения к поколению. На каждом этапе возможны небольшие изменения генотипа, и потомство получается слегка отличным от родителей. При этом действительно необходимо, чтобы изменения были малыми. Если изменение затронет один из жизненно важных процессов, то потомство не выживет. Клетки и организмы должны поддерживать линию жизни, которая тянется к ним от первых, первобытных клеток. Если какое либо поколение не сможет создать жизнеспособное потомство, всё то, что было накоплено предыдущими поколениями, все те биологические "открытия" и "изобретения", которые
208
были "сделаны", будут утеряны. Поэтому эволюция гораздо более ограничена в "свободе", чем традиционные технологии человеческой цивилизации.
Если мы создали компьютер, который не функционирует должным образом, например, в ходе тестирования новых микросхем или клавиш клавиатуры, мы просто выбросим его и вернемся к проектированию нового образца. Мы просто потеряем определенное количество времени и денег и более ничего. Если же произвести изменение какого-либо критически важного молекулярного компонента в клетке, то такое изменение должно быть всегда правильным, иначе клетка погибнет, и эволюционный процесс прервется.
Особенности сохранения изменений в ходе естественной эволюции. Конечно же, эволюция продолжается, несмотря на такие печальные последствия изменения критически важных систем. Свидетельством этому является то многообразие форм жизни, которое окружает нас. Клетки имеют несколько уровней защиты, которые позволяют экспериментировать с новыми молекулярными структурами.
Во-первых, структура данного белка может быть продублирована в геноме. Поэтому одна из копий может быть модифицирована без опасности потерять исходную информацию. Дупликация генов в ДНК чрезвычайно распространенное явление. Например, 200 миллионов лет назад ген, кодирующий гемоглобин, белок, который переносит кислород в крови, был продублирован. Это позволило второй форме гемоглобина адаптироваться для выполнения других функций, в то время как исходная форма продолжала выполнять свою функцию в крови. Новая форма постепенно выработала большее сродство к кислороду, связывая его более сильно, чем исходный гемоглобин. Сейчас этот специализированный гемоглобин используется во время внутриутробной жизни плода в его крови,перехватывая кислород из крови матери.
Во-вторых, эволюции помогает закон больших чисел. Клетки редко существуют в одиночку. Обычно, колония бактерий, косяк рыбы, стая птиц или стадо скота является биологически адекватной единицей. Внутри такой естественной популяции больше возможностей для эксперименти-
рования. Влияние случайной летальной мутации нивелируется, если
209
оставшаяся часть популяции выживет. А отдельные особи с редко встречающимися улучшениями могут впоследствии доминировать в следующих поколениях. Постепенно эти изменения приводят к развитию популяции, образуя новые варианты организмов и, в конце концов, к появлению совершенно новых особей.
Вирус иммунодефицита человека (ВИЧ) демонстрирует эволюционную силу популяции и тот прогресс, который становится возможным вследствие эволюционной оптимизации. Когда ВИЧ репродуцируется, он использует фермент ДНК-полимеразу для копирования своего малого генома. Работа этого фермента не надежна, он делает гораздо больше ошибок, чем подобные ДНК-полимеразы других клеток. Такой брак в работе, тем не менее, является большим преимуществом. В инфицированном организме каждый день ассемблируется 10миллиардов вирусов. Многие из них имеют мутации в геноме, причем многие из этих мутаций являются летальными. Но популяция вирусов настолько большая, что значительная часть вирусов остается нормальной и продолжает существование популяции, причем некоторые из мутировавших вирусов оказываются даже более жизнеспособны, чем исходные вирусы. Например, лечение каким-либо лекарственным препаратом против ВИЧ приводит к гибели нормальных вирусов, но мутировавшие вирусы выживают. Через неделю такой лекарственноустойчивый штамм уже будет доминировать в популяции.
Это эффектный пример эволюции в действии, хотя и во много раз более ускоренной по сравнению с теми темпами эволюции, которые нам более привычны. Естественным популяциям высших организмов необходимы сотни и тысячи лет, чтобы зафиксировать эволюционные изменения. Это объясняется высокой надежностью работы их ДНКполимераз и большим временем жизни каждой особи. А популяция ВИЧ может модифицироваться за считанные дни вследствие небрежной работы её реплицирующего фермента и огромного числа вирусов в популяции.
Биотехнология биомолекулярных комплексов. Эволюционная изменчивость обеспечивается мутациями и дупликацией генов. Большое число вариантов тестируется в популяции, медленно улучшая и
оптимизируя каждый компонент. Эволюционная оптимизация создала
210
такие утонченные и нежные наномашины, целенаправленное проектирование которых в конструкторских бюро человеческой цивилизации трудно себе даже представить. Например, у белков часто широкий диапазон подвижностей их частей является частью их функционирования: они используют захват цели и сложные перемещения частей для изменения и управления активностью. Более того, эти перемещения не являются переходом между несколькими фиксированными состояниями, подобно двум состояниям настенного электрического выключателя. Вместо этого эти движения приспособлены к условиям постоянного и случайного теплового движения индуцированного водным окружением, поэтому эти движения должны рассматриваться как структурированный ансамбль многих функциональных конформаций. Повторить, спроектировать аналогичную наномашину–это задача колоссальной сложности, вряд ли посильная человечеству сегодня, поскольку конструирование машин и механизмов требует полного знания и описания всех возможных конформаций в каждом состоянии. А для эволюционного "конструирования" эта задача является легкой. Эволюция делает одновременно много небольших изменений и сохраняет в потомстве только те из них,которые работают.
Эволюция не проектирует ничего перед началом конструирования – вместо этого она создает много, очень много прототипов и сохраняет наилучшие. Эволюция, однако, ограничена необходимостью передачи полученных усовершенствований через потомство. После того, как критически важная часть оборудования доведена до совершенства и находится в постоянной эксплуатации, очень сложно удалить её, или заменить её, или пытаться модифицировать её, поскольку при этом очень большой риск разрушения всей системы (в нашем, биологическом, случае
– смерть клетки). Это, в частности, справедливо для таких ключевых молекулярных процессов, как считывание и использование генетической информации, производство необходимой в клетке энергии, и размножение, каждый из которых критически зависит от согласованной работы десятков и сотен молекулярных машин.
Это привело к выдающейся унификации, единообразию и подобию, в
работе на молекулярном уровне таких ключевых процессов во всех
211
биологических системах. Все они построены из подобных основных компонентов, открытых в свое время эволюцией и использующихся с тех пор во всех последующих организмах.
Эволюционные ограничения свойств биомолекул. Биомолекулы идеально приспособлены к работе в биологических условиях, которые относительно мягкие по сравнению с теми условиями, в которых приходится работать машинам в нашем макромире. А это накладывает существенные ограничения на диапазон возможных функций бионаномашин и на те условия, в которых они могут работать. Практически все биомолекулы созданы таким образом, чтобы работать в водной среде. Большинство сконструированы для работы при температуре 37° С, нейтральной рН и физиологической концентрации солей в цитозоле. В специальных случаях могут быть проведены определенные модификации для того, чтобы расширить диапазон стабильности, но основные ограничения для ковалентно связанных органических молекул, которые должны зачастую самоассемблироваться в более сложные комплексы, достаточно жестко ограничивают диапазон приемлемых условий.
Необходимо учитывать еще одну существенную особенность природных биомолекулярных механизмов. В большинстве случаев биологическая эволюция создала бионаномеханизмы с относительно малым временем жизни. Например, большинство белков существуют только одни сутки. Этот подход прямо противоположен подходу в макромашиностроении, где машины и агрегаты создаются для длительного функционирования, в то время, как биомашины предназначены для одноразового использования при выполнении какой-либо одной задачи. Даже те биоструктуры, которые на первый взгляд кажутся неизменными, такие, например, как кости, и те перманентно дизассемблируются, восстанавливаются и перестраиваются. В макро-машиностроении такого нет. И хотя при таком подходе происходит перерасход энергии и других ресурсов на постоянную перестройку биосистем, это оправдано с точки зрения адекватной реакции биосистемы на изменяющиеся внешние условия, что характерно для жизни вообще. Возможно именно такой, биологический, подход к созданию искусственных объектов, которые, по
212
окончании использования, будут самоуничтожаться, поможет решить экологические проблемы сегодняшней индустрии.
6.5. ПРИМЕРЫБИОМОЛЕКУЛЯРНЫХ КОМПЛЕКСОВ
Рассмотрим несколько наиболее впечатляющих примеров бионаномолекулярных комплексов.
Фермент тимидилат синтаза. Фермент тимидилат синтаза осуществляет механосинтез углерод-углеродных связей (рисунок 72).
Рисунок 72 – Тимидилат синтаза: а –схема молекулы, кофактор( белый) расположен в левом верхнем кармане фермента, б –исходное расположение молекул
перед каталитическим |
актом, метильная |
группа |
связана |
с кофактором, |
|
в – расположение молекул после |
катализа, |
метильная |
группа |
присоединена к |
|
нуклеотиду; 1 –кофактор |
, 2 –метильная |
группа, 3 –тимидин |
|
|
|
213
Инициирование создания такой связи может быть обеспечено специальными молекулярными решениями. При этом совершенно недостаточно сблизить атомы углерода на необходимое расстояние. Для успешного проведения углеродного механосинтеза при биологических условиях атом углерода должен быть активирован, и та область, куда должен быть присоединен этот атом, также должна быть приготовлена для образования связи. Тимидилат синтаза осуществляет специфическую реакцию механосинтеза углерода, присоединяя новую метильную группу к нуклеиновому основанию. Метильная группа, показанная на рисунке черным цветом, активируется, присоединяясь к молекуле кофактора, показанной на рисунке белым цветом. Кофактор устроен таким образом, чтобы удерживать метильную группу, образуя метастабильный комплекс, но быть в более стабильном, низкоэнергетическом состоянии без нее. Когда кофактор отдает метильную группу, кофактор переходит в это, энергетически более выгодное положение, делая тем самым невозможным обратный перенос метильной группы. Тимидилат синтаза точно размещает в пространстве молекулу нуклеотида и активированный углерод и принудительно осуществляет перенос метильной группы. Простая близость нуклеотида и метильной группы не достаточна, вместо этого фермент должен организовать идеальное для протекания реакции молекулярное окружение. Нуклеотид при этом так окружен функциональными группам фермента, что симметрия его электронного облака изменяется в пользу реализации переноса углерода.
РНК полимераза. Следующий пример природной бионаномашины
– РНК полимераза (рисунок 73).
Молекула ДНК переносит генетическую информацию. Эта информация хранится в чрезвычайно плотноупакованном виде. Одна бактериальная клетка размером в микрометр хранит в себе около 70 килобайт информации в своем геноме. Обычный компактдиск на одном микрометре хранит только один бит информации. Генетическая информация хранится в форме, которая химически стабильна и продублирована, что обеспечивает ее восстановление в "аварийных" ситуациях. Структура ДНК является и "архивом технологических
214
регламентов" для сборки всех структур организма, и активным участником и регулятором синтеза матричных РНК.
Генетическая информация хранится внутри двойной спирали ДНК в виде последовательности нуклеотидов спаренных водородными связями.
Рисунок 73 – ТранскрипцияДНК |
ферментом РНКполимераза в молекулу РНК |
РНК полимераза копирует информацию с ДНК в про-мРНК. Поверхность двойной спирали ДНК также содержит информацию, управляющую самим процессом инициации, элонгации и терминации транскрипции. Эта " экстрагенетическая" информация считывается белками, которые присоединяются и "обертываются" вокруг спирали ДНК. Даже степень жесткости и напряженности конкретного участка ДНК используется для управления местоположением, перемещением и функционированием молекул,которые взаимодействуют с ДНК.
215

Рибосома. Рибосома( рисунок 74) – это самодостаточная фабрика, в |
закреплен в липидной мембране (мембрана показана на рисунке серым |
|
которой происходит биосинтез белка. Эта фабрика осуществляет операции |
прямоугольником). |
|
модульной сборки всех необходимых макромолекул, считывания |
|
|
генетической информации с линейного носителя ( мРНК) |
и |
|
соответствующего упорядочения 20 аминокислот в белковую цепь. |
|
|
Рисунок 74 – Схемарибосомы
Белковая нить любой длины и с любым порядком аминокислот может быть синтезирована просто в результате предварительного формирования соответствующей мРНК. Рибосомы универсальны. Любой белок может быть синтезирован на рибосоме, используя стандартный набор стартовых и стоповых инструкций и стандартный генетический код.
АТФ синтаза. АТФ синтаза (рисунок 75)является одновременно и вращательным мотором, и генератором. АТФ синтаза трансформирует энергию, превращая электрохимическую энергию в химическую, и наоборот. В нижней части рисунка изображен" мотор", который приводится во вращение электрохимическим градиентом, и который
216
Рисунок 75 – СхемаАТФ синтазы: 1 –ротор, 2 –статор
Мотор состоит из ротора, образованного циклически расположенным кольцом белков, и статора. Статор управляет потоком протонов через мембрану и использует энергию этого потока для вращения ротора.
В верхней части рисунка изображен" мотор", который приводится в движение химической энергией гидролиза АТФ. Этот мотор расположен в цитозоле и состоит из кольца из шести белков, окружающих эксцентричный ротор. Гидролиз АТФ изменяет конформацию белков, окружающих ротор,стимулируя,тем самым,вращение ротора.
Вся эта сложная конструкция используется для работы в двух направлениях.
217

(1) Электрохимический мотор в мембране может вращать |
Для того чтобы просто держать в руках баскетбольный мяч необходимо |
|
химический мотор, который в этом случае будет осуществлять реакцию |
около двух триллионов (2×10 12) миозиновых" |
шажков". Однако бицепсы |
синтеза АТФ из АДФ,то есть будет генератором АТФ. |
человека содержат в миллионы раз больше молекул миозина, поэтому |
|
(2) Химический цитозольный мотор за счет энергии гидролиза АТФ |
только небольшая часть миозина в мускулах участвует в генерации силы в |
|
будет вращать электрохимический мотор, который будет работать в |
данный момент времени. |
|
качестве насоса, перекачивающего протоны через мембрану и |
Белок опсин. Белок опсин( рисунок |
77)является светочувстви- |
увеличивающего градиент протонов по обе стороны мембраны. |
тельным белком. |
|
Актин и миозин. Актин и миозин образуют "машину", произ- |
|
|
водящую мышечное сокращение (рисунок 76). |
|
|
|
|
|
|
Рисунок 77 – Схемамолекулы мембранногобелка |
опсина |
|
Рисунок 76 – Схемарасположенияактиновых |
нитейимиозина в саркомере |
Поскольку биомолекулы при ассоциации взаимодействуют своими |
||||
|
|
|
|
поверхностями, они легко распознают изменения в конформациях |
||
Мышечные клетки заполнены чередующимися филаментами |
поверхности соседей. Поэтому большинство биосенсоров преобразуют |
|||||
миозина (серые "снопы") |
и актина (темные" спирали"). |
Химическая |
внешний световой или химический сигнал в изменение формы |
|||
энергия АТФ превращается в механическую работу в молекулах миозина. |
поверхности. Опсин содержит в себе светочувствительную молекулу |
|||||
Множество миозиновых головок" карабкаются" вдоль актиновых волокон. |
ретиналя( ретинола), |
погруженную внутрь глобулы. Ретиналь, поглощая |
||||
В сокращающейся мышце каждая молекула миозина может совершить за |
фотон,претерпевает |
цис-транс переход( рисунок 78). |
||||
одну секунду пять "шагов" |
длиной10нм |
вдоль актинового филамента. |
|
|
|
|
|
218 |
|
|
|
219 |
|