

палеонтология бондаренко
.pdfПоперечное сечение кораллитов может быть округлым, эл липтическим, многоугольным, а их форма и размеры нередко из меняются в пределах одной колонии. Наряду с крупными кораллитами появляются и мелкие, а иногда пузыревидные образования, слагающие сплошной промежуточный скелет. Внутренние полости кораллитов могут сообщаться друг с другом с помощью пор, со единительных трубок, соединительных пластин; в двух послед них случаях формируется несплошной промежуточный скелет.
Во внутренней полости кораллитов находятся вертикальные и горизонтальные элементы. Септы могут быть гребневидными, массивными, чешуевидными, шипообразными и пластинчатыми! Днища табулятоморфных кораллов имеют форму горизонтальных полных или прерывистых пластин, а также воронкоили пузыре видных образований. Поперечник отдельных кораллитов 0,1-10 мм, а колоний — до 1,5 м.
Табулятоморфные кораллы известны почти повсеместно, встречаясь со среднего кембрия до неогена включительно. Они вели неподвижный донный образ жизни, обитали в нормальноморских бассейнах и участвовали в палеозойском рифостроении.
Подкласс Tabulatomorpha включает три надотряда: Chaetetoidea, Tabulatoidea, Heliolitoidea.
Надотряд Chaetetoidea (O-N). Скелет хететоидей состоял из плотно прилегающих друг к другу мелких, почти капиллярных кораллитов, слагавших массивные колонии (рис. 117). Размеры колоний иногда могли достигать 3 м и более. Внутри кораллитов наблюдаются горизонтальные днища и единичные септы в виде
Рис. 117. Надотряд Chaetetoidea (O-N)
а — внешний вид колонии с плоскостями перерыва роста; б, в — поперечное и продольное сечения
218
септальных выступов. Рост септальных выступов завершается раз делением материнского кораллита на две и более дочерние особи. Поперечные сечения кораллитов округлые, многоугольные, ме андрически изгибающиеся, звездчатые. Для хететоидей характер ны общие плоскости остановки роста, поэтому колония иногда раскалывается на серию скорлуповатых пластин.
Надотряд Tabulatoidea (G2-P). Табулятоидеи имели кустистостелющиеся, кустистовертикальные, массивные и цепочечные ко лонии. Кораллиты роговидные, цилиндрические или призмати ческие, их поперечное сечение квадратное, многоугольное, ок руглое, эллиптическое, полулунное. Септы мелкие шипообразные, реже в виде чешуи, пластин или отсутствуют. Число септ непос тоянное. Днища горизонтальные, реже воронко- и пузыревидные. У некоторых табулятоидеи внутренние полости соседних кор&ч- литов сообщались с помощью пор, соединительных трубок и со единительных пластин. В составе надотряда выделяется 7 отря дов; ниже описано пять.
Отряд Auloporida (62 -Р). Наиболее примитивный отряд, име ющий кораллиты роговидной, реже цилиндрической формы. Они соединены друг с другом в местах почкования, образуя кустис тые, обычно стелющиеся по субстрату колонии (рис. 118). Днища и септы развиты слабо или отсутствуют.
Отряд Lichenariida (O-S,). Массивные колонии, как и в отря де Favositida, имеющие вид пчелиных сот, но стенки кораллитов сплошные, без соединительных пор и каналов.
Отряд Halysitida (02 -S). Цепочечные колонии из вертикаль но расположенных кораллитов, имеющих овальное сечение, срас тающихся узкими сторонами и образующих одно- и многорядные цепочки. Септальные образования и горизонтальные днища раз виты хорошо (см. рис. 118).
Отряд Favositida (02 -Р). Массивные колонии сложены призма тическими кораллитами (сотовые кораллы). Форма колоний раз нообразна — от лепешковидных до цилиндрических и массивноветвистых. Соединительные образования представлены порами, реже каналами, обычно закономерно расположенными на стенках. Септы разнообразные: пластинчатые, шипообразные, чешуевидные. Днища — от горизонтальных до пузырчатых. Уникальные находки фоссилизированных полипов с 11-13 щупальцами в кораллита.х сняли дискуссию о том, не являются ли фавозитиды, а имеете с ними остальные табулятоидеи и гелиолитоидеи губками (рис. I I1')
Отряд Syringoporida (02 -Р,). Кустистые колонии пк U»II щ вертикальных цилиндрических кораллитов (рис. 120). Кор.шишм сообщались друг с другом с помощью соединительных фуИочп-.. Септы и днища хорошо развиты. Обычно днища шцишкшиишыс, реже горизонтальные.
28* Л1 )

Рис. 118. Отряд Auloporida (62 -Р) и отряд Halysitida (02-S)
а-в — Aulopora (G-P): а, 6 — общий вид стелющейся колонии, в — схема распо ложения кораллитов на субстрате; г-е — Catenipora (03 -S): г — внешний вид колонии, д, е — поперечное и продольное сечения; ж — Halysites (03-S) — попе речное сечение
Надотряд Heliolitoidea (02 -D2 ). Колонии массивные разнооб разной формы, состоящие из кораллитов и промежуточного ске лета. Кораллиты обычно цилиндрические с двенадцатью, реже шестью септами и многочисленными днищами. Промежуточный скелет по одной версии представлял собой видоизмененные ко раллиты (гетероморфные компоненты); по другой — подошвен ные и боковые разрастания кораллитов (цененхима). У отряда Proporida (03 -S) промежуточный скелет состоял из пузыревид ных образований, а у отряда Heliolitida (03 -D2 ) — из полигональ ных трубок с более частыми днищами, чем в кораллитах (рис. 121).
Геологическая история табулятоморфных кораллов началась в среднем кембрии с возникновения роговидных кораллитов аулопорид без септ и днищ или с их очень слабым проявлением. В ордовике от аулопорид произошли сотовые колонии без пор
Рис. 119. Отряд Favositida (02 -Р)
а-в — Favosites (03 -D,): а - объемная реконструкция фрагмента колонии; б, в- поперечное и продольное сечения; г - фоссилизированные полипы с щупальца ми; ранний силур, Канада (Copper, Plusquellec, 1993); д, е - известковые трубки червей-комменсалов среди кораллитов фавозитид. Д — днища, п — поры, с — шипообразные септы, ш — щупальца
Рис. 120. Отряд Syringoporida (02 -Р,)
а-&- Syringopora (Ог С): а - внешний вид колонии, б, в - поперечное и про дольное сечения
220
в |
к |
д |
к |
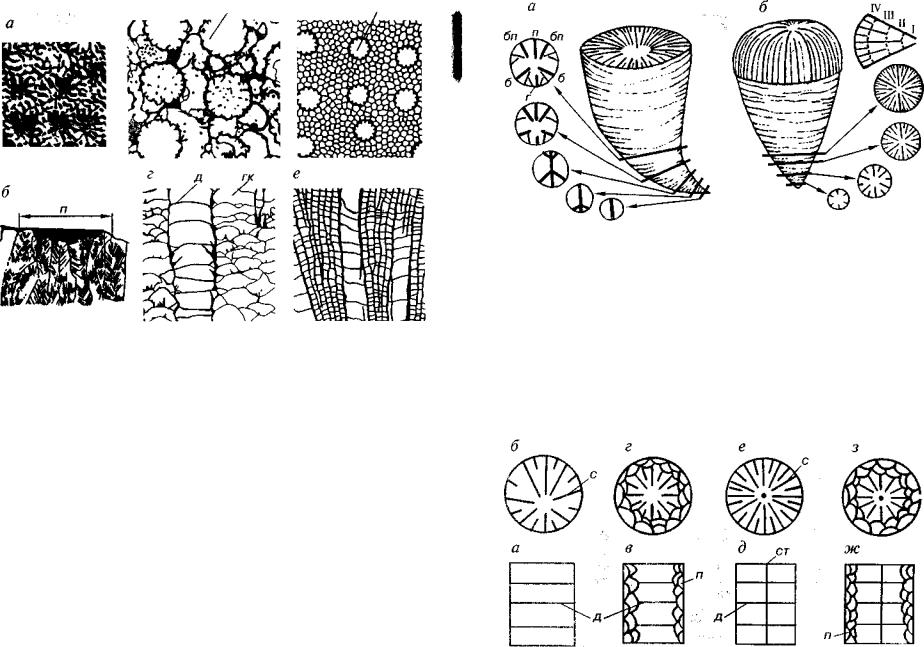
Рис. 121. Надотряд Heliolitoidea (02 -D2 )
а, б — Coccoseris (Оэ); в, г — Propora (03 -S); д, е — Heliolites s.lato (03 -D2 ); а, в, д — поперечные и б, г, е — продольные сечения, гк — гетероморфные ком поненты, д — днища, к — кораллиты, п — внутренняя полость кораллита
(Lichenariida, O-S,) и кустистые колонии с соединительными труб ками (Syringoporida, 02 -Р,). Лихенарииды в течение среднего-поз днего ордовика дали начало цепочечным колониям (Halysitida, 02 -S), массивным сотовым колониям с соединительными обра зованиями (Favositida, 02 -Р) и массивным колониям с гетероморфными компонентами (Heliolitoidea, 02 -D2 ). В ордовике, вероят но, также от лихенариид возникли хететоидеи.
Подкласс Четырехлучевые кораллы, или Ругозы. Subclassis Tetracoralla, или Rugosa
(греч. tetra — четыре; koratlion — коралл)
Палеозойские одиночные и колониальные организмы с из вестковым скелетом. Форма одиночных тетракораллов — рого видная, цилиндрическая, призматическая. Они, как правило, пок рыты эпитекой, представляющей внешний морщинистый слой, что и определило второе, не менее распространенное название этого подкласса — Rugosa (лат. ruga — морщина, морщинистый).
Рис. 122. Заложение септ у четырехлучевых (а) и шестилучевых (б) кораллов а — двусторонне-симметричное заложение септ с образованием главной (г), про тивоположной (п), боковых (б) и противоположных боковых (бп); б— циклическое
заложение септ — I—IV циклы
Длина одиночных ругоз обычно не более 25 см при попереч нике до 6 см, но некоторые позднепапеозойские ругозы достига ли почти длины клыка мамонта. Поперечник кораллитов в коло ниях до 4 см, а самих колоний — до 1,5 м.
Септы закладываются закономерно. Обычно сначала возни кает одна септа, в дальнейшем она распадается на две: короткую (главную) и .длинную (противоположную), затем появляются 4 боковые септы. В результате образуется 6 секторов, но только в
Рис. 123. Схема строения однозонных (а, б), двухзонных (в-е) и трех юппич (.« , о
|
четырехлучевых кораллов |
' |
д — днища, п — пузырчатая ткань, с — септы, ст — IUHIIMII. |
222 |
223 |

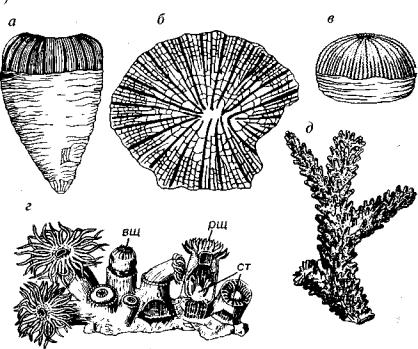
Рис. 124. Одиночные четырехлучевые кораллы (О-Р)
а — внешний вид, б, в — однозонный коралл Amplexus (С); г, д — двухзонныи коралл Caninia (С); е, ж — двухзонныи коралл Bothrophyllum (С2_3); 3, " — трехзонный коралл Neopaliphyllum (S); к, л — пузырчатый коралл ChavsakiafS^-Dj); м-о — крышечный коралл Calceola (D,_2). к — крышечка, оп — осевые пузыри, пп — периферические пузыри
четырех из них закладываются новые септы, так как около проти воположной септы дополнительные септы не возникают (рис. 122). На взрослой стадии все септы образуют один или два цикла, в последнем случае формируются большие и малые септы, череду ющиеся друг с другом. Иногда у позднепалеозойских ругоз число
224
Рис. 125. Колониальные четырехлучевые кораллы (02 -Р)
а, б — однозонный коралл Favistina (02_3); в, г — двухзонныи коралл Hexagonaria (D2_3) (ориг. Л. М. Улитиной); д, е — трехзонный коралл Petalaxis (С2)
циклов достигало 3-5. В точках заложения септ образуется про странство — фоссула, которое фиксируется расхождением септ и углублением днищ. Лучше всего развита фоссула около главной септы. Способ заложения септ, наличие фоссул, утолщение глав ных и соседних с ними септ и ряд других признаков резко нару- ч шают радиальную симметрию ругоз, превращая ее в бирадиальную или двустороннюю.
Поперечное сечение одиночных кораллов и кораллитов ко лоний может быть круглым, многоугольным, трехили четырех угольным. Скелетные элементы кораллитов (септы, днища, пузы ревидные образования, столбики) весьма разнообразны по строе нию и сочетанию друг с другом.
Общепринятые критерии для разделения тетракораллов на от ряды отсутствуют, в их эволюции прослеживается усложнение стро ения за счет появления новых скелетных элементов (рис I).) 125) У форм, возникших в раннем ордовике, развиты только uiimim (однозонные кораллы, О-Р), иногда отсутствуют и они V и'ч руки, которые известны с позднего ордовика, имелись JIIIIIIII.I и «жоно
стенок пузыревидные образования |
диссеиимсты (лиуч интыс |
|
кораллы, О, I'). С силура известны наиболее СИПАМИ у( фпгипые |
||
29 |
2С97 |
2,25 |
i ri i KI кораллы, у которых были развиты днища, пузыревидные об разования и простой или сложный столбик (трехзонные кораллы, S-P). Иногда вся внутренняя полость коралла заполнена пузыре видными образованиями (пузырчатые кораллы, S-D). Кроме того, выделяется группа одиночных крышечных кораллов (S-D). Четырехлучевые кораллы появились в ордовике и существовади по пермь; при этом колониальные формы возникли позже (средний ордовик), чем одиночные (ранний ордовик). Ругозы обитали в основном в верхней сублиторали морских тегоговодных бассей нов, могли участвовать в рифообразовании.
Подкласс Шестилучевые кораллы. Subclassis НехасогаМа
(греч. hexa — шесть; korallion — коралл)
Одиночные и колониальные организмы, особенно широко распространенные в современных тропических и субтропических морях. Это преимущественно стеногалинные формы, хотя неко торые из них, например актинии, встречаются и в бассейнах по ниженной солености типа Черного и Белого морей. Вокруг щелевидного рта располагаются щупальца, число которых обычно крат но шести. В гастральной полости находится шесть или двенадцать мягких перегородок, нижние края которых свободно свисают вниз.
В состав этого подкласса входят как бесскелетные формы (ак тинии), так и те, которые имеют известковый скелет (склерактинии).
Отряд Scleractinia (T-Q). Одиночные и колониальные совре менные и ископаемые кораллы (рис. 126). Форма одиночных ко раллов дисковидная, коническая, грибообразная, цилиндричес кая, червеобразная с поперечником до 30 см. Форма колоний мас сивная, массивно-ветвистая или кустистая, высота колоний до 1 м, диаметр до 3 м. Поперечное сечение кораллитов в колониях круг лое или многоугольное. Септы закладываются закономерно (см. рис. 122). Сначала появляется 6 септ, затем возникает еще 6. В результате формируется 12 секторов, в которых образуется 12 септ
ит.д. По мере роста кораллита септы удлиняются и возникает от 2 до 6 циклов септ, приподнятых над чашкой. Во внутренней по лости кораллитов наблюдаются днищеподобные и пузыревидные образования, а иногда и столбик. У некоторых склерактинии ске лет состоит из кораллитов и промежуточного скелета, имеющего различное строение. Одиночные формы, подобно тетракораллам, могут иметь хорошо развитую эпитеку, которая в отличие от них не доходит до верхнего края, уступая место краевой зоне септ, выступающих наружу за пределы чашки.
Склерактинии появились в триасе; в среднем триасе они были
иостаются до сих пор рифостроящими организмами, с ними
Рис. 126. Подкласс Hexacoraila (T-Q)
а~в — одиночные формы: а, б — Montlivaultia (J-K); в — Cyclolites (К-Р2 ); г, д — колониальные формы: г — внешний вид современной формы, д — Acropora (P2-Q)
вщ — втянутые щупальца, рщ — расправленные щупальца, ст — столбик
связано образование органогенных известняков. Современные рифостроящие шестилучевые кораллы приурочены к мелководью тепловодных бассейнов, где они живут на глубинах не свыше 90 м. Кроме того, в современных морях известны также глубоководные и холодноводные коралловые банки вплоть до глубин почти 1500 м. Одиночные склерактинии живут гораздо глубже — до 6000 м.
Подкласс Восьмилучевые кораллы. Subclassis Octocoralia
(греч. octo — восемь; korallion — коралл)
К этому подклассу относятся преимущественно колониаль ные животные, обитающие в морских бассейнах с нормальной соленостью. Они живут на всех глубинах, достигая глубоковод ных желобов ультраабиссали. Внешнее разнообразие восьмилучевых кораллов очень велико: они представлены различно окрашен ными морскими перьями, красными, черными и голубыми ко раллами, красными морскими органчиками и т.д. (рис. 127). Это
2 9 * |
227 |
226
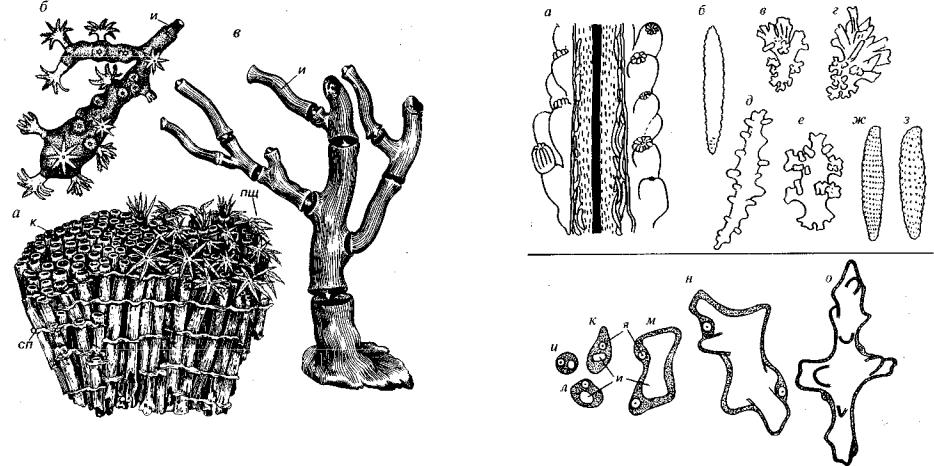
Рис. 127. Подкласс Octocoralla (V?, O-S, K-Q)
a — Tubipora(Q); б — Corallium (K-Q); в — Isis s.lato ( P - Q ) . H — известковые стержни, к — кораллит, пщ — перистые щупальца, сп — горизонтальные соеди нительные пластины
организмы прикрепленные, хотя некоторые морские перья могут передвигаться, подпрыгивая, или частично зарываться в осадок с помощью мускулистого основания.
Восьмилучевые кораллы имеют восемь перистых щупалец и восемь мягких мезентериальных перегородок в пищеварительной полости. Среди них есть формы скелетные и бесскелетные. Ске лет имеет мезоглеальное положение и состоит из органических (роговых) и известковых спикул с различными примесями, рас положенных изолированно или слившихся в плотную или «губ чатую» ткань (рис. 128). Между кораллитами обычно располага ется промежуточный скелет, представляющий собой результат деятельности общей мягкой ткани между ними. Размеры коло ний октокораллов свыше 2 м в высоту.
228
Рис. 128. Спикулы восьмилучсвых кораллов (Schiock and Twenhofel, 1953; Bengsion, 1981; Woodland, 1906)
a — фрагмент веточки современного коралла, на срединном срезе в центре виден органический стержень, вокруг него мелкие, а далее — крупные спикулы; на внешней поверхности полипы с щупальцами; б-ж — современные спикулы; з — силурийская спикула; и-о — последовательные стадии внутриклеточного образо вания известковой спикулы. и — известковый элемент, я — ядро клетки
Сведения о находках восьмилучевых кораллов отрывочны. Видимо, они появились в венде, где найдены отпечатки колони альных организмов, несколько напоминающие морские перья. Спикулоподобные элементы, сходные со спикулами октокорал лов, встречены в отложениях ордовика и силура (см. рис. 128), а достоверные остатки известны с мела и поныне. Суммируя ска занное, геологический возраст октокораллов: венд?, ордовик — силур, мел — ныне.
229
ТИП ГРЕБНЕВИКИ. PHYLUM CTENOPHORA
(греч. kteis, ktenos — гребень; phoros — несущий)
Гребневики вместе с книдариями относятся к разделу ради альных двухслойных животных. Они объединяют бесскелетных одиночных морских животных, у которых имеется восемь греб ных пластинок. У основной группы гребневиков эти пластинки, ориентированные радиально, способствуют активному планктон ному образу жизни. Тело прозрачное, радиально-симметричное, овальное или мешковидное (рис. 129). Ротовое отверстие распо лагается на одном его конце, а на противоположном находится орган равновесия, или аборальный орган.
Для происхождения трехслойных животных наибольший ин терес представляет небольшая группа гребневиков, перешедших к донному ползающему образу жизни. При этом тело становится двусторонне-симметричным, а главное, закладываются зачатки третьего зародышевого листка — мезодермы, что позволяет свя зывать появление трехслойных животных с гребневиками. В ис копаемом состоянии гребневики не известны; в последнее время появились недостоверные указания на находки отпечатков греб невиков в девоне.
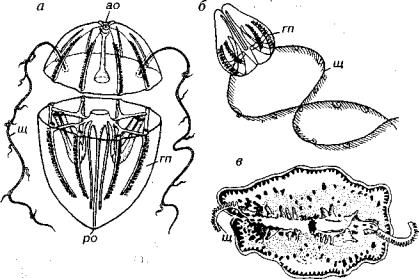
Рис. 129. Тип Ctenophora (современность)
а — схема строения гребневика; б — радиально-симметричный гребневик; в -4- двусторонне-симметричный гребневик, ао — аборальный орган, гп — гребные пластинки, ро — ротовое отверстие, щ — щупальца
230
РАЗДЕЛ ДВУСТОРОННЕ-СИММЕТРИЧНЫЕ, ИЛИ ТРЕХСЛОЙНЫЕ.
DIVISIO BILATERIA, ИЛИ TRIBLASTICA
(лат. Ы — два, дважды; lateralis, lateris — бок, сторона)
Двусторонне-симметричные трехслойные животные, у кото рых закладываются три зародышевых листка (см. рис. 78). В ос новном это формы водные, хотя многие черви, большинство чле нистоногих и некоторые моллюски вышли на сушу, а насекомые освоили и воздушную среду. Положение ротового отверстия на эмбриональной и взрослой стадиях, а также способ закладки ме зодермы явились основой для выделения двух подразделов: первичноротые и вторичноротые. Трехслойные животные известны начиная с венда.
ПОДРАЗДЕЛ ПЕРВИЧНОРОТЫЕ.
SUBDIVISIO PROTOSTOMIA
(греч. protos — первый; stoma — рот)
Первичноротые животные характеризуются тем, что ротовое отверстие взрослого организма располагается на месте бластопора зародыша, а мезодерма закладывается телобластическим спо собом (см. рис. 78). К этому подразделу относят от 9 до 26 типов, в учебнике описаны только пять.
ТИП ПРИАПУЛИДЫ. PHYLUM PRIAPULIDA
К приапулидам относятся низшие черви, представленные в современной фауне семью родами, число родов ископаемых так же около семи.
Тело приапулид удлиненное, несегментированное, длиной от 2 до 10-15 см, с элементами радиальной симметрии. На переднем конце имеется расширенная часть — хоботок, а на заднем у неко торых форм присутствует хвостовая жабра (рис. 130). Хоботок иногда снабжен снаружи радиально расположенными шипиками и крючками, передний конец хоботка может заворачиваться внутрь гели I Ыщтарительная система сквозная, животные преимущесТИППН1 чинные. Простота организации приапулид проявляется в прими i пином строении нервной системы, отсутствии кровенос ной < in Юмы, дыхание осуществляется всей поверхностью тела. Жишннмг рпштыкшолмс, личинка не трохофорная.
231

Рис. 130. Тип Priapulida (G2-Q)
а — реконструкция Ottoia (62 ); б — Priapulus (Q); в, г — Halicryptus (Q): в — передний конец тела, г — схема расположения крючьев на переднем конце (а — Gould, 1989; б-г — Жизнь животных, 1968, т. 1)
Приапулиды ведут исключительно зарывающийся образ жиз ни, обитая в плотных песчаных и песчано-глинистых грунтах. Животные «забуриваются» в грунт с помощью сильно развитой мускулатуры. Сначала расширяется хоботок и увеличивается по перечник хода или норки, а затем туловище подтягивается впе ред. За 1 час червь может продвинуться на расстояние 1 м. Харак тер движения тесно взаимосвязан с радиальной симметрией тела.
Приапулиды появились не позднее чем в кембрии. Они явля лись постоянным компонентом сообществ, обитавших в море на плотных песчано-илистых фунтах. Об этом свидетельствуют на ходки приапулид в среднекембрийских сланцах Бёрджес (см. рис. 252). Средний кембрий — современность.
ТИП КОЛЬЧАТЫЕ ЧЕРВИ. PHYLUM ANNELIDES
(лат. annellus — колечко)
Кольчатые черви обитают в водной и наземной среде. Их тело имеет метамерное строение, так как оно состоит из множества сегментов — «колечек», заключенных в кожно-мускульный ме шок. В каждом сегменте повторяется набор органов почти всех систем. У аннелид хорошо развиты пищеварительная, кровенос ная, половая, нервная, мышечная, выделительная и иногда дыха тельная системы. Органы чувств концентрируются на переднем конце тела. Примитивные аннелиды имели на каждом сегменте параподии — выросты кожно-мускульного мешка.
Рис. 131. Тип Annelides (V-Q)
а — Serpula (S-Q); б— крышечка; в, г — Spirorbis (O-Q); д — схема расположения кристаллов кальцита в известковой трубочке червя; е — сколекодонты — хитиновый челюстной аппарат; ж — следы ползания и проедания грунта; з — Dickinsonia (V)
Водные кольчатые черви иногда ведут неподвижный образ жизни и в этом случае строят агглютинированные, роговые, хи тиновые или известковые трубки (рис. 331). Форма трубок раз лична: от удлиненных слабоизогнутых до неправильно изгибаю щихся или почти шгоскоспиральнозавитых. При массовых посе лениях известковые трубки рода Serpula и близких ему родов могут давать серпулиты. Иногда в ископаемом состоянии сохраняются
232 |
30-2697 |
т т о |
|
|

крышечки, подобные таковым у современных форм, а также хи тиновый челюстной аппарат — сколекодонты.
Наиболее древние кольчатые черви были бесскелетными ор ганизмами. Начиная с венда известны разнообразные следы пол зания, проедания грунта и зарывания в него. Это результат жиз недеятельности различных организмов, в том числе и аннелид. Следы ползания и проедания грунта называются биоглифы и яв ляются одной из разновидностей иероглифов. Кольчатые черви играют значительную роль в процессе биотурбации, т.е. в про цессе переработки осадка, при зарывании в грунт и пропускании осадка через кишечник.
С кембрия и до наших дней встречаются аннелиды с извес тковым скелетом. Современные морские формы живут на раз личных глубинах вплоть до ультраабиссали. Они, в частности помпейский червь, входят в сообщества денсали.
Кольчатые черви являются предками членистоногих и мол люсков, с которыми их сближают единый тип развития зародыша (эмбриогенез) и сохранение в той или иной степени метамерности, что свойственно членистоногим и некоторым низшим мол люскам. Эмбриональное развитие завершается формированием личинки трохофорного типа (см. рис. 78).
ТИП ЧЛЕНИСТОНОГИЕ. PHYLUM ARTHROPODA
(греч. arthron — сустав; pous, podos — нога)
Трехслойные первичноротые двусторонне-симметричные жи вотные, произошедшие от кольчатых червей. Тело членистоногих имеет метамерное строение. Оно состоит из 8-180 сегментов, раз лично сросшихся между собой, и членистых конечностей (рис. 132). Современные формы населяют воду, сушу и воздух. К ним при надлежат раки, крабы, скорпионы, клещи, пауки, насекомые (рис. 133). Это наиболее многочисленный из всех типов, на его долю приходится около 3 млн видов, что составляет более по ловины от общего числа видов царства животных; главенствую щая роль приходится на долю насекомых.
У членистоногих присутствуют следующие системы: пищева рительная, нервная, кровеносная, дыхательная, половая, мышеч ная, выделительная — и хорошо развиты органы чувств, в том числе простые и сложные глаза. Сложные глаза состоят из мно жества простых с общим внешним покровом (голохроические) либо каждый простой глазок заключен в самостоятельную капсу лу (шизохроические).
234
Рис. 132. Схема строения основных представителей членистоногих
а, б — подтип Trilobitomorpha, класс Trilobita: а — подкласс Polymera, б — под класс Miomera; в, г — подтип Chelicerata: в — эвриптерус, г — паук; д-з — подтип Crustaceomorpha, класс Crustacea: д — панцирь рака, е —- известковый домик усоногого рачка, ж — известковая створка остракоды, з — хитиновая створка листоногого рачка; и, к — подтип Tracheata, класс Insecta: и — муравей,
к — крылатое насекомое
Тело защищено хитиновым скелетом в виде панциря, створок, щитков и т.д. Скелет в различной степени может пропитываться углекислым кальцием или фосфатом кальция. Жесткий хитино вый скелет препятствует росту животного, поэтому для большин ства членистоногих характерна линька, при этом панцирь или
зо* |
235 |
Рис. 133. Образ жизни и условия обитания членистоногих а — трилобиты; б-д — ракообразные: б — ракушковые рачки, в — листоногие
рачки, г — усоногие рачки, д — рак; е-з — хелицеровые: е — эвриптерус, ж — мечехвост, з — паук; и-л — трахеинодьацащие: и — многоножки, к, л — насекомые
створки сбрасываются. Мягкое тело растет преимущественно между линьками. Некоторые членистоногие не линяли, их скелет над страивался и увеличивался в размерах.
При разделении членистоногих на подтипы основное внима ние уделяется строению головного отдела и особенностям конеч ностей этого отдела, а также специфике органов дыхания. Ниже описаны четыре подтипа: трилобитообразные, ракообразные, хе лицеровые и трахейные. Членистоногие появились в докембрии, достоверные находки известны начиная с венда; в геологическом отношении наиболее важны трилобиты.
|
ПОДТИП ТРИЛОБИТООБРАЗНЫЕ. |
|
SUBPHYLUM TRILOBITOMORPHA |
•.,;,, |
Класс Трилобиты. Classis Trilobita |
|
(лат. tri — три; греч. lobos — доля) |
К подтипу вымерших трилобитообразных животных относят до десяти различных классов. Наиболее многочислен и разнооб разен класс трилобитов, существовавший только в палеозое. В ископаемом состоянии от них сохраняются панцири и отпечатки, лишь в исключительных случаях можно наблюдать строение ко нечностей, глаз, пищеварительного тракта и т.д.
пц ро
Рис. 134. Схема строения трилобитов а — вид сверху; б— реконструкция трилобита с брюшной стороны; в — продольный
разрез; г-е — типы лицевых швов: г — переднещечный, д — заднещечный, е — угловощечный. а — антеннулы, ао — анальное отверстие, бб — боковые борозды, бг — борозды глабели, бд — базальные дольки, бл — боковые лопасти, или плев ры, бр — боковые ребра, г — глабель, гк — глазные крышки, гщ — головной щит, дк — двуветвистые конечности, ж — желудок, зв — задняя ветвь лицевых швов, зк — затылочное кольцо, к — кишечник, кк — краевая кайма, кш — краевые шипы, м — мембрана, о — осевая часть панциря, ош — осевые шипы, п — плевры, пб — продольные борозды, пв — передние ветви лицевых швов, пц — панцирь, ро — ротовое отверстие, с — сегменты, то — туловищный отдел, фл — фронтальная лопасть глабели, х — хвостовой шип, хщ — хвостовой щит, щу —
щечный угол, щш — щечные шипы
236