

Биомы, Гл
.9.pdfГлава 9
ЗОНАЛЬНЫЕ ТИПЫ БИОМОВ СУШИ
Áиом представляет собой сочетание конк-
ретных экосистем. Зональные типы биомов формируются при определенном соотношении тепла и влаги, включают растительные сообщества и животное население, неразрывно связанные друг с другом. Как крупные экологические единицы биосферы они, в свою очередь, могут быть подразделены на более мелкие типы – региональные типы биомов.
Зональные типы биомов – совокупности естественных экосистем со сходной структурой и обликом сообществ, сформировавшиеся в процессе исторического развития и отражающие гидротермический потенциал территории. Их границами являются рубежи, за пределами которых экосистемы данного зонального типа не могут существовать из-за несоответствия условиям среды (Огуреева, Котова, 1997).
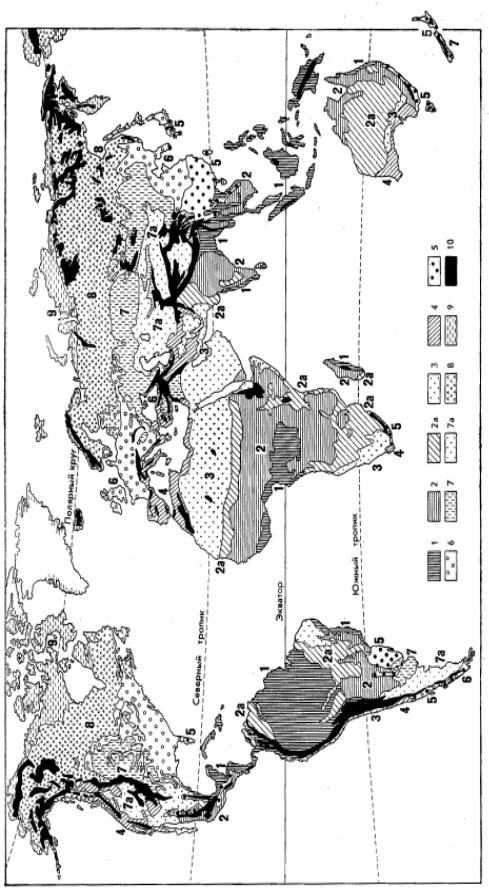
Биогеографическая картина мира, основанная на выделении биотических царств суши, отражает уникальность флоры и фауны, являющуюся следствием дивергентного пути эволюции (см. гл.8). Выделение зональных типов биомов подчеркивает черты сходства органического мира, формирующегося в условиях одинаковой обеспеченности теплом и влагой. Об общей картине распределения зональных типов биомов суши дают представление карты растительности и почв (рис. 9.1, 9.2).
Наглядной моделью распространения зональных типов ландшафтов служит гипотетический материк (см. рис. 2.14). Сопоставление с картой растительных и почвенных зон Земли показывает, что эта модель достаточно полно отображает географию зональных типов биомов в целом (табл. 9.1). Зоны в таблице перечислены в том же порядке, в каком дано их описание в тексте.
Перейдем к характеристике зональных типов биомов суши. В качестве основы взя-
ты представленные в табл. 9.1. системы зональных типов ландшафтов и растительных зон Земли. При описании биомов использовались работы В. В. Алехина, Л. В. Кудряшова, В. С. Говорухина (1961), Н. А. Бобринского (1951), Г. Вальтера (1968), А. Г. Воронова, Н. Н. Дроздова, Е. Г. Мяло (1985), У. Зедлага (1975), А. Г. Исаченко и А. А. Шляпникова (1989), П. Д. Ярошенко (1975) и др.
9.1. Биомы жаркого пояса
Жаркий пояс Земли простирается к северу и югу от экватора в тропические широты; в каждом полушарии его ограничи- вает годовая изотерма 20 îС. Важнейшим фактором зональной дифференциации в жарком поясе является увлажненость (см. гл. 2). В соответствии с закономерностями атмосферной циркуляции и режимом выпадения атмосферных осадков формируется зональный ряд биомов: дождевых тропических лесов, тропических листопадных лесов, саванн и пустынь. Переходное положение между биомами жаркого и умеренного поясов занимают биомы субтропических зон.
9.1.1. Дождевые тропические леса
Биомы дождевых тропических лесов – одни из самых древних и богатых на Земле (рис. 9.3). Их ареал охватывает несколько царств: Неотропическое, Эфиопское, ИндоМалайское, Австралийское. Количество видов растений и животных здесь больше, чем во всех других биомах вместе взятых.
По климатическим особенностям различа- ют леса экваториального и субэкваториального поясов, муссонных дождей, горных склонов. Наиболее полно свойства вечнозеленых дождевых тропических лесов (гилей) выражены в бассейне Амазонки, по берегам Гвинейского залива и в бассейне Конго, на Малаккском полуострове, островах Малайского архипелага, на северо-вос- токе Австралии (рис. 9.4). Главный фактор, определяющий формирование вечнозеле-
146
Ва.Г у:льтер
растительныхема онз поЗемли,
.Ðèñ .9 Ñõ.1
тропические– еполувечнозелены
склоновгорных пояса(леса туманов);2
лесадевые инизменностей
дожтропические
1 – евечнозелены
ропическиесубт :зоны
иТропические
-ñóá
тропиче– иские
убтропикс 3ов;
ропикт иов
îíûç
частичноов, овникизлак
арникуст
олк кючих
слизаро
иаванны
ствсте сенные
åñà,
са;ле –2а лесухие
листи еопадны
тепунк .)7а
растительно– отместь, вченная
(в.ш угихдр стахме
(35°)30° .ñ è.ø .þ
олькт доо
ñîâþ
оронуст лпо
èåù âñÿ
юстира
пустыи прони,
скиетропиче лупустынипо
-вечнозеле
äÿìè;æ –5
сласти дозимними
чаяю обаридные
ждо вклдями),
зимними(с
ластьоб олистнойжестк стирастительно
–4 ходнаяпере
скаяиче :зоны
иУмеренная тарк
периодомзимним
умереннойлеса сзоны
летнезелены– лиственныее
прохладнымм 6климатом;
постояннос нывлаж
лиственныеные умереннолеса зонытеплой зонили
совлю то -30
ств поорону
òñÿþ
стира(про
хс зимойолодной
нипусты
по– илупустыни
оныз прерии,(степи, 7апампа);
мереннойу
овникизлак
îíà);ç –7
ояпок льная(немора
северноготундры ария;полуш -10 сгоры
северноголесов ария;полуш –9 зонаарктическая
ххвойны
пустыня;холодная –8 зонабореальная
–Тибете
.ø â),
35° .ñ è.ø .þ
альпийскийчая нивальныи поясай
растительности,поясами ювкл
миразличны
147
148
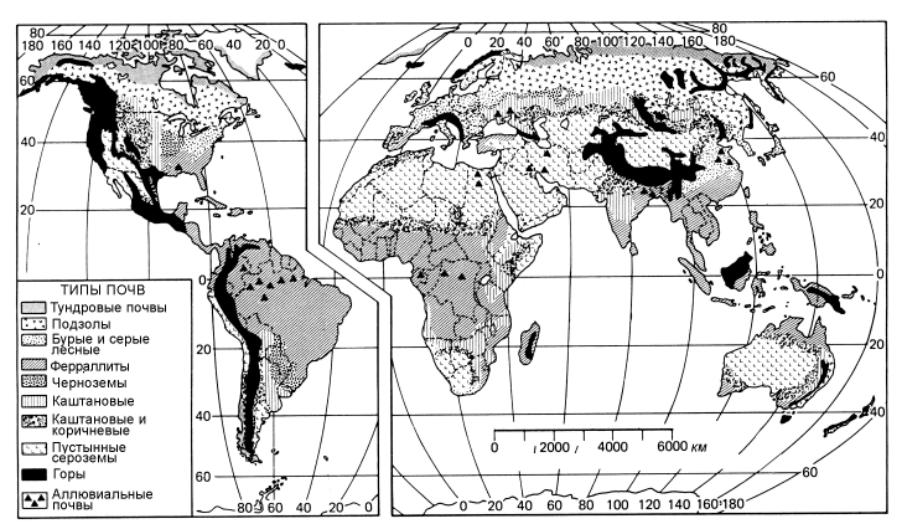
Рис. 9.2. Зональные типы почв, по Earl D. Shaw
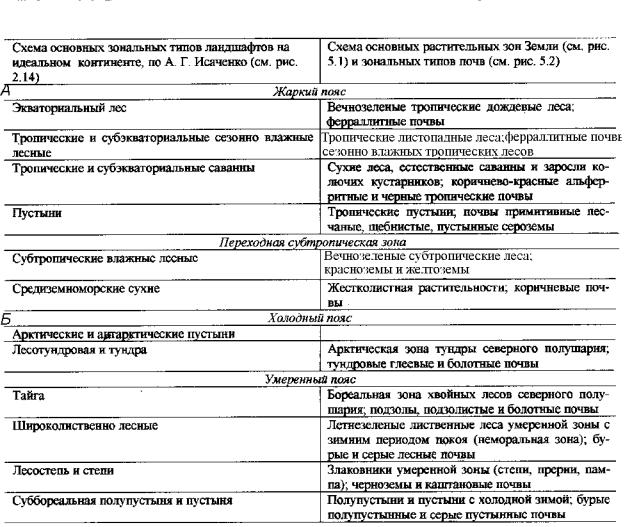
Таблица 9.1. Сопоставлениеосновныхзональныхтиповландшафтовнаидеальном континенте, растительных и почвенных зон Земли
ных дождевых тропических лесов, – сохраняющийся на протяжении миллионов лет благоприятный теплый и влажный климат. Обилие тепла и влаги обеспечивают круглогодичную вегетацию растений. Сезонная ритмика нечеткая.
Средние месячные температуры воздуха от 25 до 35 îС, их годовая амплитуда всего 2 – 4 îС (суточные амплитуды больше годовых). Атмосферных осадков выпадает от 1500 до 4000 мм, а на наветренных склонах гор до 10 000 мм и более. Для существования гилей, однако, более важно не общее количество осадков, а распределение их в году. В субэкваториальном поясе дождевой климат имеет летний пик осадков, когда солнце стоит в зените (субэкваториальный муссон). Эти зенитальные дожди вызывают резкий подъем уровня воды в реках и затопление низменностей. С переходом в тропи- ческие широты продолжительность зимнего сухого сезона увеличивается. Климати-
ческой границей дождевых лесов является продолжительность периода засухи более двух месяцев.
Длительность дня и ночи вблизи экватора в течение года приблизительно одинакова. Это постоянство имеет большое зна- чение для суточного ритма жизни растений и животных. Красочное описание суток во влажном тропическом лесу дает А. Уоллес: “Рано утром, в первые два часа после восхода солнца, небо было неизменно безоблачным, термометр стоял на 22 – 23 °С, тяжелая роса или следы ночного дождя, отягощавшие листья, быстро испарялись под палящими лучами солнца, поднимавшегося как раз на востоке и быстро приближавшегося к зениту. Вся природа дышала свежестью, молодая зелень и почки цветов быстро распускались... Жара усиливалась с каждым часом и около 2 час. дня достигла 33 – 34 °С, к каковому времени умолкали голоса всех птиц и зверей. Листья, такие све-
149

Рис. 9.3. Девственный тропический лес в Бразилии, по A. Schimper
Рис. 9.4. Ареал дождевых тропических лесов, по П. Ричардсу
150
жие и сочные утром, блекли и свешивались |
доточились наиболее устойчивые, но бес- |
вниз, лепестки цветов осыпались. В июне |
плодные соединения алюминия и железа. |
и июле около полудня обыкновенно выпа- |
Продуктивность лесов огромна. Запас |
дает сильный дождь, приносящий желан- |
древесины свыше 5000 ц/га (в Бразилии – |
ную прохладу. Туча приближается всегда |
до 17 000 ц/га), а годовой прирост 350 – |
одинаковым, весьма любопытным манев- |
500 ц/га и более. Парадокс влажных тропи- |
ðîì. |
ческих лесов заключается в том, что пыш- |
Сначала прохладный морской бриз, по- |
ная растительность развивается на мало- |
являющийся около 10 час. утра и усилива- |
мощных бедных почвах. Это становится |
ющийся по мере того, как поднимается |
возможным благодаря очень быстрому кру- |
солнце, начинает ослабевать и, наконец, |
говороту биогенных солей. Отмершая мас- |
прекращается совершенно. Тогда жара и |
са ветвей и листьев быстро, в течение не- |
электрическое напряжение воздуха стано- |
скольких недель, разлагается и минерали- |
вятся просто невыносимыми. Тяжелое рас- |
зуется. В лесах мало опавшей листвы и не- |
слабление овладевает всеми, даже обитате- |
редко видны пятна оголенной земли. В раз- |
ли леса выдают это своими движениями. |
ложении опада большое участие принима- |
Но вот на востоке показывается белый ту- |
ют термиты; главная же роль, по-видимо- |
ман и начинает собираться в кучевые обла- |
му, принадлежит грибам. |
ка, нижний край которых становится все |
Корневая система деревьев не прони- |
темнее и темнее. Весь восточный горизонт |
кает на большую глубину, вся она сосредо- |
внезапно чернеет; эта мгла распространя- |
точена в первых десятках сантиметров ма- |
ется все большой и больше, заслоняя под |
ломощного слоя почвы, где образует фильтр, |
конец и солнце. Затем мощный порыв вет- |
поглощающий растворимые питательные |
ра с туманом проносится по лесу, раскачи- |
вещества. Важную роль в питании растений |
вая вершины деревьев, сверкает ослепи- |
играет микориза. Высокие деревья с повер- |
тельная молния, раздается удар грома и раз- |
хностной корневой системой мало устойчи- |
ражается стремительный ливень. Такие гро- |
вы. Досковидные корни – специальное при- |
зы быстро проходят и оставляют за собой |
способление, подобно контрфорсам у готи- |
неподвижные исчерна-синие тучи, остаю- |
ческих соборов они поддерживают высокие |
щиеся на небе до самой ночи. Вместе с тем |
стволы (рис. 9.5). |
вся природа освежена; лишь лепестки цве- |
Особенности почв влажных тропичес- |
тов и листья кучами валяются под деревья- |
ких лесов следует учитывать при их хозяй- |
ми. К вечеру природа пробуждается к но- |
ственном освоении. Издавна здесь приме- |
вой жизни, которой полон каждый куст, каж- |
нялась подсечно-огневая система земледе- |
дое дерево. На другой день солнце снова по- |
лия: лес вырубался, сжигался, участок об- |
является на безоблачном небе, возобновляя, |
рабатывался для выращивания тех или |
таким образом, круговорот тропического |
иных сельскохозяйственных культур (рис. |
дня, в течение которого перед нашими гла- |
9.6). Но запасы питательных веществ в по- |
зами как будто проходят весна, лето и |
чве ограничены, и они быстро вымывают- |
осень…” |
ся дождями. Через пару лет возделанный |
Почвы влажных тропических лесов |
участок приходится забрасывать. Губитель- |
формируются на мощной бесплодной фер- |
на сплошная вырубка лесов, особенно на |
раллитной (Al2O3, Fe2O3) коре выветрива- |
склонах, которая ведет к смыву почв и ин- |
ния. Основной запас питательных веществ |
тенсивной эрозии. Около 100 000 км2 òðî- |
сосредоточен в живых растениях. Полага- |
пических лесов подвергается деградации |
ют, что растения получили этот запас мил- |
ежегодно в результате антропогенного воз- |
лионы лет назад в процессе биохимическо- |
действия. Обнажение бесплодной феррал- |
го разложения материнских горных пород |
литной коры выветривания на больших |
богатого минералогического состава. В ко- |
площадях – угроза того, что влажные тро- |
нечном счете, в коре выветривания сосре- |
пические леса могут утратить способность |
151

Рис. 9.5. Досковидные корни, придающие большую устойчивость высоким стволам деревьев с поверхностной корневой системой, по А. Кернеру
152

Рис. 9.6. Выжигание тропического леса под сельскохозяйственное угодье и его последствия:
Справа – эрозия склонов после уничтожения лесной растительности
êсамовосстановлению и на их месте возникнут вторичные растительные сообщества с более бедным видовым составом и низкой продуктивностью.
Богатство биома влажных тропических лесов находит выражение в огромном разнообразии видов, жизненных форм и в сложной структуре лесного сообщества. 70% видового разнообразия растений составляют деревья (цветковые). Участие трав незначительно, обычно они растут на опушках леса. Преобладают травы, своими размерами не уступающие деревьям, бананы достигают высоты 6 м. Их подземные органы живут неопределенно долго.
Число видов деревьев на 1 га может превышать 100; переходя от одного дерева
êдругому, исследователь каждый раз обнаруживает новый вид. Лесные сообщества не отделены друг от друга четкими границами, а связаны постепенными переходами. Такое явление называется горизонтальной континуальностью растительного покрова.
Так же сложен вопрос о вертикальном сложении леса: между 30 – 40-метровыми гигантами, образующими верхний полог, и деревьями в 3 – 5 м существуют виды деревьев, образующие кулисообразные полога, перекрывающие друг друга (рис. 9.7). Это дает основание говорить о вертикальной континуальности, которая усиливается благодаря массе внеярусных растений – лиан и эпифитов.
Число видов эпифитов и лиан в дватри раза превосходит число видов деревьев (рис. 9.8). Особую жизненную форму
представляют деревья-душители, большинство из которых относится к роду Ficus. Благодаря птицам их семена попадают на ветви крон деревьев верхнего яруса. На первой стадии развития росток ведет жизнь эпифита. Однако сразу начинается быстрый рост корней: один корень устремляется в землю и снабжает растение водой и питательными солями; ответвления воздушных корней охватывают ствол хозяина и душат его (рис. 9.9).
Влажный тропический лес создает своеобразный микроклимат под своим пологом. Кроны деревьев смыкаются; в лесу ца-
Рис. 9.7. Вертикальная структура дождевого тропи- ческого леса, по Г. Вальтеру:
Разные виды деревьев высотой от 4 – 5 до 30 – 35 м образуют переходы, затрудняющие выделение четких элементов вертикальной структуры лесного сообщества
153

рит зеленый сумрак, редкий луч солнца пробивается сквозь густую листву. Здесь влажно и душно, как в оранжерее. Условия обитания накладывают отпечаток на строение растений. Стволы гладкие и стройные; кора тонкая и светлая. Корка, как правило, не образуется. У многих деревьев досковидные и ходульные корни. Почки защищены очень слабо, почечные чешуи отсутствуют. Форма и размер листьев в нижних ярусах леса мало варьируют: обычно они большие и цельнокрайние. В более высоких ярусах леса листья вырабатывают приспособления для защиты от яркого солнца и потери влаги – их размеры уменьшаются, они становятся кожистыми, блестящими, с толстой кутикулой. Представление о “вечнозелености тропических лесов” создается потому, что смена листьев в течение года происходит постепенно: одни листья разворачиваются
Ðèñ. 9.8. Орхидеи-эпифиты, по А. Кернеру из почек, другие опадают. Наряду с этим у некоторых деревьев может происходить одновременное массовое сбрасывание листвы, хотя рядом стоящие деревья того же вида могут оставаться зелеными.
В нижнем ярусе влажность воздуха постоянно высокая. Защиты от испарения растениям здесь не требуется. Напротив, у них есть ряд приспособлений, которые облег- чают транспирацию: тонкие листья с большой поверхностью и большим количеством устьиц; листья, в которых отдача влаги обеспечивается капельными устьицами на концах листовых жилок, из которых выдавливаются капли воды.
Многие тропические деревья цветут и плодоносят круглый год. На одном дереве имеются цветочные почки и цветки, зеленые и спелые плоды. Иногда деревья зацветают дружно раз в 10 лет. Учи- тывая, что деревья одного

вида в тропическом лесу растут на большом |
двудольных деревьев нужно назвать цейбу |
удалении друг от друга, приходится только |
(Ceiba) – дерево, возвышающееся над всем |
догадываться, благодаря какому сигналу они |
лесным пологом; бертоллецию (Bertolletia |
зацветают одновременно, обеспечивая тем |
excelsa), плоды которой называют бразиль- |
самым возможность перекрестного опыле- |
скими, или американскими, орехами; гевею |
ния. Одной из особенностей плодоноше- |
(Hevea brasiliensis), дающую каучук; деревья |
ния у некоторых видов растений является |
с ценной красной древесиной – махагонию |
каулифлория, т. е. образование цветков, а |
и цезальпинию. К нижнему ярусу относит- |
затем и плодов непосредственно на ство- |
ся шоколадное дерево (Theobroma cacao); |
лах или толстых ветвях. Наиболее извест- |
дынное дерево (Carica, около 40 видов), |
ный пример – шоколадное дерево(рис. 9.10). |
сладкие плоды которого содержат фермент |
|
папаин, благоприятствующий пищеваре- |
|
нию. Деревья всех ярусов переплетают ли- |
|
аны. Среди эпифитов выделяются своим |
|
разнообразием орхидеи, папоротники, виды |
|
из семейства Bromeliaceae. В заводях Ама- |
|
зонки растет знаменитая гигантская кув- |
|
шинка виктория регия (рис. 9.13). |
|
Кроны густого тропического леса яв- |
|
ляются средой обитания множества зверей |
|
и птиц с ярким оперением (рис. 9.14). По- |
|
чти все они проводят большую часть жиз- |
|
ни на деревьях, не опускаясь на землю. |
|
Приспособление характерное для животных |
Рис. 9.10. Плоды шоколадного дерева (какао) на ство- |
неотропиков – цепкохвостость. Цепкие хво- |
ле и ветвях дерева. Рядом продольный разрез плода |
сты обезьян (рис. 9.15), опоссумов, древес- |
|
ных муравьедов и дикообразов, енотов-но- |
Животный мир влажных тропических |
сух помогают им ловко передвигаться с вет- |
лесов характеризуется огромным богат- |
ки на ветку. Типичными представителями |
ством и разнообразием: более 10 000 000 |
фауны Неотропического царства в гилее яв- |
видов, в основном насекомых и других бес- |
ляются неполнозубые – ленивцы. Их когти |
позвоночных (рис. 9.11). Благоприятные ус- |
срослись в серповидные крючья, с помощью |
ловия обитания – тепло, влажно, обилие зе- |
которых животные висят на ветвях безо вся- |
леного корма, масса убежищ, способствуют |
кого напряжения мускулов (рис. 9.16). |
формированию сообществ со сложными |
Впрочем, представление о том, что в |
биотическими отношениями. Круглый год |
тропическом лесу на каждом шагу встреча- |
климатические условия благоприятны для |
ются интересные птицы и звери, не соот- |
размножения и развития. Основная жизнь |
ветствует действительности. Девственный |
сосредоточена в кронах деревьев; у земли |
лес кажется мертвенно тихим: в густой чаще |
лес кажется безжизненным. |
трудно передвигаться и в то же время легко |
В Неотропическом царстве дождевые |
спрятаться. Лишь невидимые стаи обезьян |
тропические леса занимают огромные про- |
и птиц производят подчас невероятный |
странства на Амазонской низменности, где |
шум. Изредка встречаются равнинный та- |
их описал еще А. Гумбольдт, введший в на- |
пир и похожие на кабанов пекари. Крупные |
уку термин “гилея” (рис. 9.12). А. Л. Тахтад- |
грызуны замещают здесь копытных; наи- |
жян (1978) выделяет особую Амазон-скую |
больших размеров достигает капибара |
флористическую область, которая включает |
(Hydrochoerus capybara), до полутора мет- |
500 эндемичных родов и не менее 3000 ви- |
ров в длину и массой до 60 кг. Хищники |
дов. Здесь центр распространения пальм (до |
представлены кошачьими – ягуаром и оце- |
200 видов), из которых 60 эндемичны. Из |
лотом. |
155