

Страница
---------------------------------------------------------------------------------------------------------------------
1. Происхождение высших растений. Морфологические аспекты эволюции (дифференциация тела; многообразие габитуса и жизненных форм как результат эволюции в различных условиях среды; побег как осевой орган и эволюция процесса его нарастания; два пути формирования листовых органов – филлоидный и теломный).
Происхождение наземных, высших растений ознаменовало новую эпоху в жизни нашей планеты. Первые достоверные наземные растения, известные только по спорам, датируются началом силурийского периода. До сих пор ведутся споры на тему происхождения высших растений. Существуют разные точки зрения, такие как, что высшие растения произошли либо от зелёных, либо от красных водорослей.
Момент выхода из воды на сушу стал поворотным в жизни растений, им пришлось осваивать совершенно новые условия обитания. Приспособление к новым условиям обитания повлекло за собой изменения в строении тела, размножении и образе жизни растений. По мере возникновения трудностей, растения выработали различные ткани, а клетки, слагающие их, подверглись дифференциации, в результате чего стали выполнять более узкие функции лучше, чем недифференцированные клетки низших растений.
Первая возникшая трудность – добыча воды из влажной почвы и прикрепление к ней, так как всасывать воду всей поверхностью тела, как в водной среде, было уже нереально. Должны были появиться органы прикрепления к почве, позднее превратившиеся в корни. Должны были возникнуть и осевые органы, несшие спорангии со спорами, а позднее и органы ассимиляции - листья.
Одного прикрепления к субстрату недостаточно для жизни в сменившихся условиях, полученную из влажной почвы воду нужно провести на всю высоту растения, так появилась необходимость в проводящей системе.
Соприкосновение с воздушной средой вызвало к жизни покровные ткани, защищающие растение от излишних испарений и механических воздействий, необходимость газообмена, дыхания привела к образованию устьиц, расположенных среди клеток эпидермы, а значительно меньшая плотность воздуха в сравнении с водой привела к образованию механических тканей. Для обеспечения фотосинтеза потребовалось разделение функций клеток растения. Так возникла ассимилирующая ткань.
Необходимость в дифференциации возникла уже у первых наземных растений. Корень, стебель, листья, а наличие специализированных органов резко отличает высшие растения от низших (водорослей).
Габитус от лат. hábitus — внешность, наружность, вид, облик, образ.
Жизненной формой растений называется совокупность всех признаков, определяющих их внешний облик и отражающих приспособление к условиям обитания. В первую очередь это касается признаков вегетативных органов, обеспечивающих жизнь растения и его связь со средой.
Жизненную форму у высших растений с эколого-морфологической точки зрения можно определить, как своеобразный общий облик (габитус) определенной группы растений (включая их подземные органы), возникающий в их онтогенезе в результате роста и развития в определенных условиях среды. Исторически этот габитус развился в данных почвенно-климатических и ценотипических условиях как выражение приспособленности растений к этим условиям.
По Раункиеру (1934 г.) жизненная форма возникает в историческом развитии в результате приспособления растений к климатическим условиям страны, что может служить индикатором ее климата.
Своеобразие различных жизненных форм возникает в онтогенезе растений в результате более – менее сложной и длительной цепи возрастных морфогенетических изменений. Однолетние сеянцы ели или сосны еще не деревья. В большей степени это относится ко многим травянистым многолетникам: стержнекорневые в первые годы жизни, они затем могут стать корневищными, кистекорневыми, столонообразующими, клубнеобразующими и т.п. Таким образом, жизненная форма часто меняется в онтогенезе растений.
Существенные различия наблюдаются нередко в росте и развитии одного и того же вида в разных экологических условиях, что приводит к появлению разных жизненных форм в пределах вида.
В еще большей степени изменение жизненной формы иногда наблюдается у растений при интродукции их за пределы естественного ареала.
Изучение жизненных форм различных фитоценозов способствует более глубокому познанию структуры, динамики, исторического развития растительных сообществ и их взаимосвязи со средой. Жизненная форма является адекватным выражением условий жизни организмов, поэтому их изучение в ценозах – надежное средство экологической оценки местообитания.
Эколого-морфологическая классификация Серебрякова (1964) построена на признаках формы роста, длительности жизни надземных вегетативных органов, наличии одревеснения стеблей. Она включает древесные, полудревесные и травянистые растения.
Древесные растения имеют многолетние одревесневающие надземные стебли. К ним относятся деревья, кустарники и кустарнички. Деревья - многолетние древесные растения с хорошо выраженным главным стеблем (стволом), превышающим 3 м в высоту (Берёза, сосна). Кустарники - многолетние древесные растения, у которых главный ствол хорошо выражен лишь в начале жизни, затем теряется среди равных ему нескольких скелетных стеблей, возникших из спящих почек, а позже отмирает, их высота от 1 до 6 м (Крушина, шиповник). Кустарнички - многолетние древесные растения, у которых главный стебель имеется лишь в начале онтогенеза, а затем сменяется боковыми надземными стеблями из спящих почек его наземной части (Вереск, брусника).
Полудревесные растения - многолетники, стебли которых на большей части своей длины травянистые и ежегодно отмирают, а одревесневают лишь в основании. К ним относятся полукустарники и полукустарнички. У полукустарников высота многолетних одревесневших нижних участков стеблей не превышает 20-30 см, а неодревесневших - 15 - 20 см (Черника).
Травянистые растения имеют неодревесневающие стебли. К ним относятся многолетние, двулетние и однолетние травы. Продолжительность жизни многолетних трав превышает 2 года (Ландыш майский, Пырей ползучий). Для завершения жизненного цикла двулетних трав требуется два вегетационных периода, при этом цветение и плодоношение происходят на втором году жизни (Морковь, Капуста). Жизненный цикл однолетних трав длится один вегетационный период (Мятлик однолетний, Марь белая).
Побег, как и корень, - основной орган растения. Вегетативные побеги в типичном случае выполняют функцию воздушного питания, но имеют ряд других функций и способны к разнообразным метаморфозам. Спороносные побеги (в том числе и цветок) специализированы как органы репродуктивные, обеспечивающие размножение.
Побег образуется верхушечной меристемой как единое целое и, поэтому, представляет собой единый орган того же ранга, что и корень. Однако по сравнению с корнем побег имеет более сложное строение. Вегетативный побег состоит из осевой части – стебля, имеющего цилиндрическую форму, и листьев – плоских боковых органов, сидящих на стебле. Кроме того, обязательной частью побега являются почки – зачатки новых побегов, обеспечивающие нарастание побега и его ветвление, т.е. образование системы побегов. Главную функцию побега – фотосинтез – осуществляют листья; стебли – преимущественно несущие органы, выполняющие механическую и проводящую функции.
Главная черта, отличающая побег от корня, - его облиственность. Участок стебля, от которого отходит лист (листья) называется узел. Участки стебля между соседними узлами – междоузлия. Узлы и междоузлия повторяются вдоль оси побега. Таким образом, побег имеет метамерное строение, метамером (повторяющимся элементом) побега являются узел с листом и пазушной почкой и нижележащее междоузлие.
Первый побег растения – его главный побег, или побег первого порядка. Он образуется из зародышевого побега, заканчивающегося почечкой, которая формирует все последующие метамеры главного побега. По положению эта почка – верхушечная; пока она сохраняется, данный побег способен к дальнейшему росту в длину с образованием новых метамеров. Кроме верхушечной, на побеге образуются боковые почки. У семенных растений они находятся в пазухах листьев и называются пазушными. Из боковых пазушных почек развиваются боковые побеги, и происходит ветвление, за счет которого увеличивается общая фотосинтезирующая поверхность растения. Формируется система побегов, представленная главным побегом (побегом первого порядка) и боковыми (побегами второго порядка), а при повторении ветвления – боковыми побегами третьего, четвертого и последующих порядков. Побег любого порядка имеет свою верхушечную почку и способен к нарастанию в длину.
Почка – это зачаточный, еще не развернувшийся побег. Внутри почки заключена меристематическая верхушка побега – его апекс. Апекс представляет собой активно работающий ростовой центр, который обеспечивает формирование всех органов и первичных тканей побега. Источником постоянного самообновления апекса являются инициальные клетки апикальной меристемы, сосредоточенные на кончике апекса.
Дихотомическое -> Моноподиальное -> Симподиальное
Лист в эволюции растений возникал 2 раза. В девоне возник энационный лист, называемый также филлоидом и микрофиллом. Он возник как чешуевидный вырост на побеге, служивший для увеличения площади фотосинтезирующей поверхности. Этот вырост нужно было снабжать водой и забирать из него продукты фотосинтеза, поэтому в него проникла проводящая система. Сейчас такие листья характерны для плауновидных и псилотовидных.
Во второй раз возник теломный лист или макрофилл. Он возник на базе группы теломов, расположенных в одной плоскости, которые уплощились и срослись. Данный вид листа характерен для хвощей, папоротников, голосеменных и цветковых.
---------------------------------------------------------------------------------------------------------------------
2. Подкласс Марсилеивые – общая характеристика на примере марсилии - происхождение; распространение; основные черты строения; особенности размножения и полового процесса.
Водные, разноспоровые растения. Гаметофиты всегда раздельнополые, редуцированные, развивающиеся в оболочках мегаспор и микроспор. Мегаспорангии и микроспорангии сосредоточены в общих сорусах, заключенных в спорокарпии.
Порядок Marsileales относятся растения, ведущие водный или земноводный образ жизни и характеризующийся, как правило, разноспоровостью. Как и у всех разноспоровых растений, у них раздельнополые гаметофиты, развивающиеся внутри оболочек мегаспор и микроспор. Мужской гаметофит редуцирован значительно сильнее, чем женский, и представлен 1-2 проталлиальными клетками и двумя очень утолщёнными антеридиями. Женский гаметофит крупнее, содержит в нижней части питательные вещества, не заключенные в клетки питательной ткани и 1-5 архегониев. Перед созреванием сперматозоидов клетки мужского гаметофита расплываются.
Так как растения водные или земноводные, оплодотворение происходит в водной среде, а развивающийся зародыш спорофита на первых этапах жизни питается за счёт веществ из женского гаметофита.
Порядок содержит одно семейство – Marsileaceae.
В пределах СССР, Средней Азии и Поволжья распространена Марсилия четырёхлистная Marsilea quadrifolia. От тонкого корневища марсилеи отходят придаточные корни и листья с длинными черешками, которые заканчиваются четырьмя листочками, по форме напоминающие листочки клевера. Их своеобразная особенность – способность ко «сну». В темноте листочки складываются и повисают вниз. В нижней половине черешка на более или менее длинной ножке сидят спорокарпии, по форме и размерам похожие на мелкое семя фасоли. Оболочки спорокарпия тверды и прочны, выдерживают пересыхание и механическое воздействие. Оболочка спорокарпия имеет листовое происхождение, возникла из четырёхлисточного спороносного сегмента листа. Внутри спорокарпия по его периферии проходит кольцом тяж хрящевидной ткани, к которому двумя рядами прикрепляются сорусы мега и микроспорангиев. Те и другие спорангии покрыты очень тонкой прозрачной оболочкой, стенкой спорангия, обычно лишённой кольца и не выполняющей защитной роли, так как функцию защиты берёт на себя стенка спорокарпия.
Раскрывается зрелый спорокарпий будучи помещённым в воду. Хрящевидная ткань при этом набухает, заставляет раскрыться спорокарпий на две створки, и из него, постепенно разворачиваясь, выходит эта ткань в виде длинного тяжа, по обе стороны которого располагаются сорусы. Каждый сорус покрыт индузиумом, тонкой прозрачной оболочкой, которая сильно редуцированна.
Мегаспора марсилеи наполнена питательными веществами, а ядро оттеснено в верхний конец, где имеется сосочковидный вырост и там оболочка тоньше.
Микроспроры имеют тетраэдрическую форму, позднее становясь округло шаровидными.
Спора всегда прорастает в воде, даже если вид земноводный, образовавший спорокарпии вне воды. Водные виды тоже всегда образуют спорокарпии вне воды, остающиеся жизнеспособными до попадания в воду. Способность спорокарпия набухать и раскрываться в воде может сохраняться долгие годы – иногда и десятилетия.
Женский гаметофит марсилеи представлен практически одним архегонием, развивающимся в месте сосочковидного выроста, надрывающего тонкую оболочку мегаспоры, а остальная часть мегаспоры погружена в водянистую протоплазму клетки. Кроме того, мегаспора с развивающимся гаметофитом может быть одета слизистым студенистым покровом, предохраняющим растение.
Мужской гаметофит состоит из одной проталлиальной клетки и двух очень упрощённых антеридиев, дающих малое число (16) сперматозоидов. Всё развитие этого растения занимает 24 часа, таким образом быстрое развитие развитие раздельнополых папоротников хорошо иллюстрирует смысл разноспоровости: маленькие, предельно просто устроенные гаметофиты быстро проходят эту ненадёжную в условиях наземного существования стадию и образуют зародыш спорофита, для которого готово большое количество питательных веществ, не использованных женским гаметофитом, но накопленных предыдущим поколением – спорофитом.
Развитие зародыша протекает быстро, он состоит из листа, корня, стебелька и ножки.
Спорокарпии распространяются как потоками воды, так и с говном водоплавающих птиц, через кишечник которых спорокарпий проходит неповреждённым. Бывает и вегетативное размножение у этих папоротников.
---------------------------------------------------------------------------------------------------------------------
3. Анатомические аспекты эволюции высших растений (эволюция основных тканей; основные типы стелей и их эволюция). Эволюционные аспекты полового процесса (жизненный цикл высших растений и основные эволюционные тенденции; гаметангии высших растений и их эволюция; равноспоровость и разноспоровость).
При выходе на сушу сформировались покровные тканми, которые играют барьерную функцию между внутренней жидкой средой организма и газообразной атмосферой. Потом, для транспирации, появились устьчные аппараты. Потом механические ткани для опоры и защиты растения, и проводящие, дабы провести вещества на всю длину растения.
Стель - центральный цилиндр.
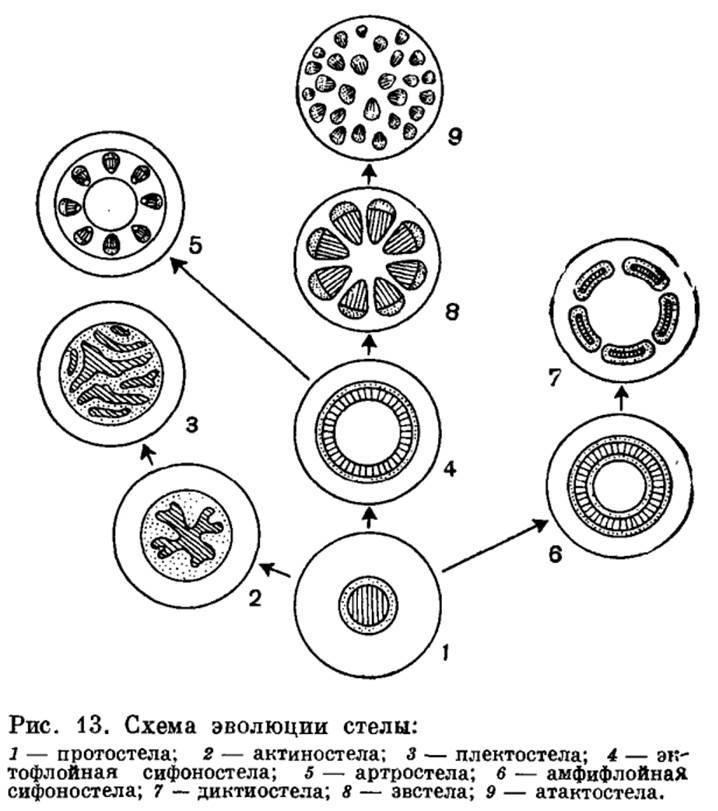
Всего существует 9 типов стелей.
1) протостель (Риниофиты)
Наиболее примитивный тип стели. В которой проводящая ткань образует сплошную массу и центральное ядро, состоящее из ксилемы, полностью окружено слоем флоэмы. Протостела не только очень проста по структуре, но и представляет собой исходный тип, от которого в процессе эволюции произошли все другие типы стелы.
2) актиностель (Некоторые примитивные Плауны)
Более подвинутый тип с радиальными выступами ксилемы.
3) плектостель (Плауны посложнее)
Результат дальнейшей специализации актиностелы. Тут ксилема разбита на отдельные пластинки, расположенные параллельно или радиально друг к другу.
4) сифоностель (Папоротники)
Имеет трубчатое строение и обладает центральной паренхимной частью, или сердцевиной. Эволюция сифоностелы сопровождается появлением паренхимных участков (лакун) в самом проводящем цилиндре. Благодаря лакунам создается непрерывная связь коры и сердцевины.
5) артростель (Хвощи)
Имеет членистое строение и характеризуется наличием центральной полости и протоксилемных лакун (каринальных каналов).
6) амфифлойная сифоностель (Папоротники)
Осевой цилиндр, в котором есть сердцевина с внутренней эндодермой, кольцо внутренней флоэмы, кольцо ксилемы и наружное кольцо флоэмы, граничащее с наружной эндодермой.
7) дигтиостель (Папоротники)
Когда сифоностела разбита на сеть или ряды продольных тяжей, получается рассеченная стела, или диктиостела.
8) эустель (Большинство семенных растений)
Возникает в результате рассечения сифоностелы. характерная для большинства семенных растений.
9) атактостель (Травянистые двудольные и большинство однодольных)
Проводящие пучки эвстелы разбросаны по сердцевине и коре так, что сходство с единой сетью или кольцом теряется совершенно. Ввиду своеобразия подобной стелярной организации она выделяется в особый тип стелы, названный атактостелой. Атактостела развилась в результате крайнего рассечения эвстелы и представляет собой в эволюционном отношении крайне специализированную форму центрального цилиндра.
спорофит 2n => спорангий 2n => материнские клетки спор 2n =/R!=> спора => гаметофит n => антеридий n (/архегоний n) => сперматозоид n (яйцеклетка n) =О!=> зигота 2n => спорофит 2n
Два пути эволюционных тенденций:
1) гаметофитный тип
2) спорофитный
У мхов гаметофит преобладает.
Чем более растение развито, тем редуцированнее гаметофит. У цветковых растений мужские гаметофиты столь малы, что помещаются внутри оболочки пыльцевого зерна и состоят всего лишь из нескольких клеток. Женский гаметофит цветковых (зародышевый мешок) помещается внутри семяпочки и состоит в наиболее распространённом случае (в случае так называемого нормального моноспорического женского гаметофита) из 7 клеток (содержит 8 — либо 7 после слияния двух ядер в центральной клетке и образования вторичного ядра). У папоротников, хвощей и плаунов гаметофит представляет собой небольшое, но самостоятельное растение, иногда называемое заростком. Заростки папоротников и хвощей фотосинтезирующие, однолетние. Заростки плаунов живут под землей, развиваются в течение многих лет и питаются за счет симбиоза с грибами.
У высших растений есть равноспоровые и разноспоровые. Равноспоровые имеют одинаковые по форме и размерам, и происхождению спорангии и споры дают начало обоеполым гаметофитам.
Разноспоровость образует разные типы спор - мегаспоры и микроспоры. Мегаспоры обычно значительно крупнее микроспор. Мегаспоры дают начало женским гаметофитам с архегониями, микроспоры дают начало мужским гаметофитам с антеридиями.
---------------------------------------------------------------------------------------------------------------------
4. Подкласс Сальвиниевые – специфичные черты строения как результат эволюции в водной среде, общая характеристика на примере сальвинии и азоллы - происхождение; распространение; основные черты строения; особенности размножения и полового процесса.
Salvinidae. Характеризуется наличием водных разноспоровых растений. Гаметофиты всегда раздельнополые и редуцированные, развиваются внутри оболочек микроспор и мегаспор.
Микроспорангии и мегаспорангии заключены в разных сорусах, спорокарпиев нет.
Порядок Salviniales представлен двумя семействами: Salviniaceae - Сальвиньевые и Azollaceae – Азолловые.
Эти растения развивают плавающий ветвящийся стебель, несущий распростёртые на поверхности воды листья. Корни есть только у Азолловых, у сальвиниевых корневидные листья. Сорусы закрыты шаровидным индузиумом, а при созревании спор клетки тапетума и содержимое недоразвитых спор образуют пенистую массу – массулу.
Распространены как в тропиках, так и в умеренных областях.
Семейство Salviniaceae содержит 1 род Salvinia, состоящий из 8 видов, распространённых в основном в Африке, однако Salvinia Natans растёт преимущественно в южных районах Евразии. Это небольшое растение обладает ветвящимся тонким стеблем, плавающим на поверхности воды, от него отходят мутовки листьев. Отходят по два листа. Корней у сальвинии нет, есть вместо них подводные, корневидные листья. Они покрыты щетинистыми волосками и выростами на эпидермисе, препятствующие смачивание листа. Если растение погрузить в воду, лист покроется серебристым слоем воздуха и сразу всплывёт, стоит только его отпустить. Корневидные листья расчленены на нитевидные доли, густо усаженные твёрдыми волосками, они выполняют не только функцию всасывания воды, но и функцию якоря, стабилизатора, держа растение в одном положении.
Кроме того, на корневидных листьях образуются сорусы спорангиев.
Таким образом корневидные листья спороносящие и защищают спорангии.
Сорусы сальвинии закрыты шаровидным двухслойным индузиумом. Они содержат ТОЛЬКО мегаспорангии или только микроспорангии. Микроспорангии мельче, в них образуется 64 микроспоры. В мегаспорангии возникает 32 клетки, способные дать мегаспоры, но получается только одна. Остальные разрушаются и вместе с клетками тапетума дают периплазмодий – питательную среду, для формирования единственнной мегаспоры, приобретающей к моменту созревания третью толстую и прочную оболочку – перину. Периплазмодий образуется и при созревании микроспор, но только из клеток тапетума.
Зрелые сорусы опускаются на дно и разрушаются, споры высвобождаются и всплывают к поверхности воды и здесь прорастают.
Из микроспор развивается редуцированный мужской гаметофит из 2 проталлиальных клеток и двух антеридиев, по 4 сперматозоида с каждого.
Мегаспоры начинают прорастать внутри своей оболочки и не выпадает из неё. Зрелый женский гаметофит, находясь в оболочке мегаспоры представлен крупной базальной клеткой, наполненной питательными веществами, в которой появляется много ядер, но оболочек нет, и не формируется никаких тканей, на которой развивается 3 или 5 архегониев.
Эта часть гаметофита высовывается из-под оболочки мегаспоры в месте разрыва её и мегаспорангия и зеленеет. При этом он значительно увеличивается, становится больше мегаспоры. Так как развитие происходит в воде, затруднений для сперматозоидов нет.
Зародыш спорофита формирует ножку, первый лист и стебель, корень совсем не развивается или очень рано исчезает, сливаясь с ножкой.
Растение может размножаться обрывками стебля, вегетативно.
Бурно разрастившись где-нибудь сальвиния будет мешать рыбакам и судам, образуя запруды.
Семейство Azollaceae. Содержит 1 род Azolla и 6 видов, растущих в тропиках и реже в умеренных областях планеты. Оно тоже плавает на поверхности воды. Ветвящийся тонкий стебель, от которого отходят придаточные корни и два ряда листьев. Каждый лист двулопастной, а его верхняя, не погружённая в воду лопасть, состоит из нескольких рядов клеток и несёт устьица на верхней поверхности. Нижняя лопасть однослойна, погружена в воду и нужна для её всасывания.
Особенностью Азоллы является её симбиоз с водорослью Anabaena azollae. Анабэна врастает в специальные полости молодого листа Азоллы. За счёт симбиоза Азолла снабжается азотом. Во Вьетнаме её специально разводят в качестве удобрения, дабы рис рос лучше. По способности накапливать азот папоротник не уступает бобовым.
Спорангии развиваются на первом боковом листе каждой ветки, на нижнем, погружённом в воду сегменте. Обычно образуются 2 или 4 соруса, всегда содержащие или только микроспоранги и только мегаспорангии. Микросорус значительно крупнее мегасоруса, закрыт индузиумом, и несёт большое число микроспорангиев, в свою очередь содержащих по 64 микроспоры, соединяющимися пенистым и твердеющим веществом из расплывшихся клеток тапетума, причём в отличие от сальвинии это вещество собирается комочками или массулами.
Мегасорусы гораздо мельче, чем микросорусы и содержат обычно один мегаспорангий, в котором образуется 32 мегаспоры, но ольео одна развивается до конца. А сестринские клетки, расплываясь образуют с клетками тапетума 4 массулы, в одну погружена мегаспора остальные располагаются сверху в виде грушевидных поплавков. Массулы снаружи окружены остатками оболочки мегаспорангия.
У микроспор вслед за образованием массул в микроспорангиях на их поверхности развиваются якоревидные крючки, или глохидии, с очень тонким основанием, легко колеблющегося от малейшего движения воды. Этими глохидиями массулы с микроспорами заякориваются на неровной оболочке мегаспор и таким образом прорастание мужского и женского гаметофита рядом друг с другом.
Мужской гаметофит Азолли развивается внутри оболочки микроспоры и внутри микроспорангия. Зрелый гаметофит предельно упрощён: он состоит из одной проталлиальной клетки и одного антеридия, с него 8 сперматозоидов.
Женский гаметофит также внутри оболочки мегаспоры, образуется крупная базальная клетка, она становится многоядерной. И ещё одна клетка поменьше сверху, она и даёт начало гаметофиту женскому. Из верхней клетки развивается бесцветная ткань, разрывающая оболочку мегаспоры и тогда на её поверхности, развивается архегоний. Если оплодотворения не произойдёт, может развиться ещё 1-2 архегония. Нижняя большая клетка служит резервуаром для пищи гаметофита и зародыша.
---------------------------------------------------------------------------------------------------------------------