

1.Лесоводство и лесоведение, различие между ними.
Лесоведение – это учение о лесе, о его природе, о закономерностях происходящих в лесу. Лесоведение основывается на дендрологии и физиологии, лесной метеорологии, почвоведение, фитопатологии, энтомологии и таксации леса. В лесоведение рассматриваются географические аспекты, закономерности возобновления и распространения леса. Объектами лесоведения являются как естественные леса, так и искусственные леса.
Лесоводством принято называть теорию и практику лесного хозяйства или деятельности, направленной на наиболее выгодное и постоянное использование леса в интересах людей. Научная дисциплина о природе леса, его биологии и экологии, закономерностях динамики в пространстве и во времени. Является составной частью лесоводства, его естественно-исторической основой. В связи с признанием многосторонней роли леса выделилось в самостоятельную научную дисциплину. лесоведение опирается на биологические науки - физиологию, дендрологию, генетику, фитопатологию, энтомологию и др.
Учебный курс лесоводства обычно начинают с раздела о природе леса – лесоведения.
2. Структура древостоя, классы Крафта, деревья-лидеры и подгон.
Особенности каждого древостоя, как и насаждения в целом, зависят от сочетания экологических факторов, от состава древостоя, его происхождения и истории роста. Но есть и общие характерные для всех древостоев черты, объясняемые спецификой взаимных отношений в лесу между деревьями, видами, возрастными поколениями, ярусами. Чаще всего взаимоотношения проявляются через внешнюю среду. Их называют косвенными. Наиболее существенным видом косвенных отношений является конкуренция. Имеют место и прямые взаимоотношения: симбиотические, паразитические, физиологические (срастание корней), химические (аллелопатия), механические (охлестывание).
Конкуренция является главной причиной дифференциации деревьев по размеру, своеобразия их формы, темпов отпада. В древостое обычно приводят классификацию Г. Крафта, предложенной в конце 19 в. Согласно этой классификации, все деревья по положению кроны в пологе делятся на классы господствующих и угнетенных деревьев.
1 – широкие мощные кроны, наиболее развиты (10%); 2 – хорошо развитые (20-30%); 3– стесненные, крона сдавлена с боков, но вершина свободная (примерно 33%) – господствующая часть; 4 – угнетенные, крона до половины полога (10-15%); 5 – заглушенная, крона под живыми деревьями (10 – 15%).
Распознать классы Крафта можно в раннем возрасте (8 – 9 лет) с вероятностью до 65 – 70%, можно диагностировать будущих лидеров (вероятность 90 – 96 %).
ПОДГОН деревья или кустарники, способствующие ускорению роста и улучшению формы ствола главной древесной породы. Создает боковое оттенение главной породы, предотвращает рост нижних ветвей и способствует очищению ствола от сучьев. Подгонами могут быть древесные породы (береза, вяз, клен, липа и др.), которые отличаются густооблиственной кроной, и кустарники (лещина, жимолость, крушина, бузина и др.).
В смешанных насаждениях роль подгона по отношению друг к другу могут играть и деревья главных пород при совместном произрастании быстро и медленнорастущих. В подгон особенно нуждаются медленнорастущие породы, в первую очередь дуб. Подгон появляется в результате естественного возобновления, но может вводиться искусственно.
3.Лесной фитоценоз, яруса, подлесок, подрост, напочвенный покров.
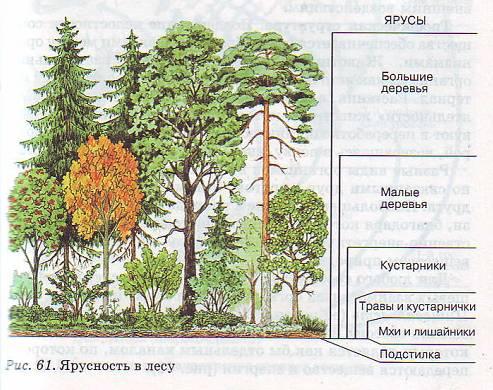
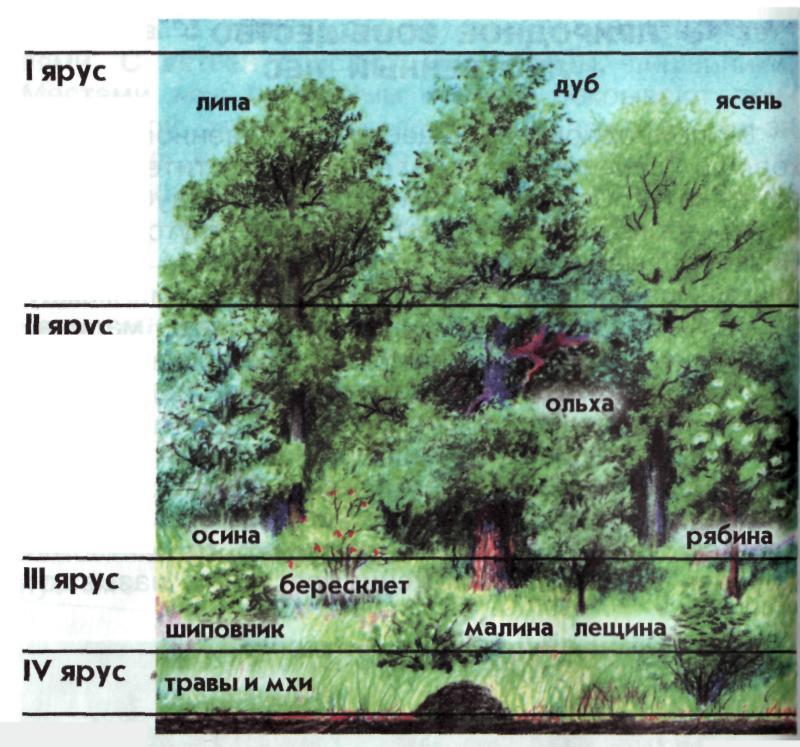
В состав лесного фитоценоза помимо древостоя входят подрост, подлесок, живой напочвенный покров. Подростом называется совокупность молодых деревьев, находящихся под пологом старого древостоя или на вырубке после его удаления, способная заменить старый древостой. Иногда древостоем называют всякое молодое поколение, в том числе появившееся на вырубке или гари от налета семян. Подлеском называют совокупность кустарников, кустарничков и полукустарников, реже деревьев, произрастающих под пологом леса, не способных достигнуть высоты верхних ярусов и образовать древостой. Появлению подроста и подлеска способствует прореживание древостоями рубками ухода или постепенными. Живой напочвенный покров – совокупность мхов, лишайников, травянистых растений и кустарничков, произрастающих под пологом леса, на вырубках и гарях. На первых этапах жизни леса под густым древостоем живой покров может отсутствовать, т.к. почва в это время покрыта слоем опада главным образом из хвои и листьев. Особенностью леса является подстилка – напочвенный слой, образующийся из растительного опада. Благодаря подстилке, ее разложению в условиях особого микроклимата наблюдается своеобразный режим почвенных процессов. Лесной фитоценоз отличается своеобразной вертикальной и горизонтальной структурой. По вертикали он делится на ярусы, сложенные разными жизненными формами растений и видами. Горизонтальная структура – наблюдается мозаичность, объясняемая разнообразием состава древостоев, групповым размещением деревьев, неоднородностью микрорельефа и почвы.
Яруса представлены на картинке
4.Определение леса по В.Н.Сукачеву и в понимании И.С.Марченко - лекции.
Тип леса — участок леса или их совокупность, характеризующиеся общим типом лесорастительных условий, одинаковым составом древесных пород, количеством ярусов, аналогичной фауной, требующие одних и тех же лесохозяйственных мероприятий при равных экономических условиях (определение В. Н. Сукачёва).
Лес (по академику В.Н.Сукачеву,1953 г) – это сообщество древесных растений, организованных конкурентной борьбой за существование, т.е. древостой это основной элемент. И.С. Марченко написал книгу «Биология лесных экосистем». Он объяснил и предложил новую концепцию понятия лес. Лес – сообщество растений, которые взаимодействуют между собой своими полями и энергиями и взаимодействуют с энергией земли (гипотеза).
В 1974г. Появилась ключевая работа, изменившая представление о лесе. Юодвалькис и Кайрюкштис выяснили следующее: при составлении моделей роста лесных культур ели в опытах с разной густотой посадки 1 – 10000/1 га, установили, что в период за 2 – 3 года до смыкания крон в рядах растения резко снижали прирост боковых ветвей в 2 раза. Вторы объяснили снижение прироста сменой конкурентной борьбы на взаимную толерантность.
5. Лес как явление географическое. Лес и почва.
Географическая среда отображает все многообразие экологических условий и определяет характер леса. Все особенности лесного хозяйства обязательно связаны с географическими особенностями. Разнообразные формы леса могут быть поняты, пишет Г.Ф. Морозов, только в связи с внешней географической средой, к которой прежде всего относят климатические, эдафические, почвенные, гидрологические, ландшафтные и другие особенности. С учетом географических факторов были составлены классификации зон растительности. Наиболее известны классификации A. Гумбольдта (начало XIX в.), русского климатолога и ботаника B.П. Кеппена (1901). В начале XX в. экспедиция В. В. Докучаева установила географичность почв, ее связь с климатом и другими факторами почвообразования. В современных правилах рубок главного пользования равнинные леса европейской части России по географическим признакам разбиты на округа: притундровый, северотаежный, среднетаежный, южнотаежный, хвойно-широколиственный и лесостепной. Леса разных географических зон существенно различаются.
Почва снабжает растения элементами питания и водой, обеспечивает их физическую устойчивость, является средой обитания животных. А растения и животные - причины образования и условия существования почвы. Почва влияет на состав, темпы роста, качество древесины, на процессы возобновления растений, служит основой классификации их местообитания. Влияние почвы существенно зависит от рельефа и горной породы.
Рельеф перераспределяет влагу атмосферных осадков, тепло, свет, биологические элементы. Существует классификация рельефа: макро-, мезо- и микрорельеф. Микрорельеф способствует выживанию подроста, например ели, ее всходов и сеянцев. Существует горизонтальная расчлененность почвы внутри насаждений: деление на парцеллы. Парцеллярная структура объясняется неоднородностью состава, выпадением сухостоя, рубками ухода, болезнями. В основном положительное влияние леса на почву заключается в увеличении ее глубины, в содержании органики, защите почвы от эрозии, сбережении влаги, смягчение микроклимата. Общая оценка: лес повышает плодородие почвы, извлекая и аккумулируя зольные элементы их грунта. Уничтожение леса в любых условиях приводит к оскуднению почвы – заболачиванию, пересыханию, обеднению.
6.Биотические факторы и лес, биоразнообразие и устойчивость лесов
Благодаря естественному отбору сообщество получает мощный гомеостатический механизм, который позволяет противостоять изменениям и сохранять равновесие (Р. Риклефс, Ю. Одум, Р. Уиттекер). В ходе естественной сукцессии возрастают глубина почвы и запас элементов питания в ней, увеличиваются темпы роста, биомасса и видовое разнообразие экосистемы, возрастает устойчивость сообщества. Это — естественные процессы. Однако в настоящее время естественные процессы существенно нарушаются. Одной из причин снижения устойчивости лесов является уменьшение разнообразия флоры и фауны, разнообразия видового, генетического и экосистемного в основном из-за неблагоприятных последствий неправильного хозяйства, прежде всего из-за погони за повышением продуктивности лесов любыми способами в ущерб природоохранным и социальным функциям. Значительно уменьшается разнообразие флоры и фауны после сплошной рубки. В древостое по вырубке количество видов таежного крупнотравья составляет к 100 годам около 30 % по сравнению с мало нарушенным лесом (П.Д.Ярошенко, 1997). Особенно большие потери всех видов флоры и фауны наблюдаются после рубки в редких местообитаниях. Результатом увлечения сплошными рубками и монокультурами, преимущественно сосны и ели, стало уменьшение площади широколиственных, дубовых, пихтовых и кедровых лесов с их своеобразной и ценной флорой и фауной. Сплошная вырубка сухостоя уменьшает количество и разнообразие видов насекомых и грибов, необходимых для разложения древесины и сохранения почвенного плодородия. Поэтому в ряде стран, например в Германии, сухостой рубят лишь там, где есть потребность в его использовании. На международной конференции во Франции в 2002 г. подчеркивалась необходимость замены хвойных культур естественными хвойно-широколиственными лесами, которые некогда преобладали в Европе. Еще в прошлом веке Г. Ф. Морозов перечислял достижения старого лесоводства, которые направлены на поддержание устойчивости: предпочтение естественному возобновлению перед искусственным, осторожность и постепенность действий, предпочтение смешанным и сложным насаждениям перед чистыми и простыми, сохранение местных рас, групповой метод возобновления и др. Обоснование изменений в привычной лесохозяйственной практике, направленных на повышение устойчивости и биоразнообразия лесов, на сохранение естественной сукцессии лесов, — одна из главных проблем лесоводства. Она обсуждается на международных конференциях, в которых принимает участие и Россия. Необходимо и в России менять некоторые установки в лесохозяйственной практике для сохранения устойчивости и биоразнообразия лесов.
Под биоразнообразием лесов понимают все находящиеся в лесах жизненные формы, включая деревья, растения, животные, грибковые образования и микроорганизмы, а также их назначение в природе. Характеризующиеся сложностью и богатством эти формы предоставляют многие жизненно важные услуги для людей.
Тем не менее, люди разрушают биоразнообразие лесов с угрожающей скоростью. Превращение лесов в сельскохозяйственные угодья, чрезмерный выпас скота, нерациональное управление, введение инвазивных чужеродных видов, создание инфраструктуры, горнодобывающих и нефтяных месторождений, антропогенные пожары, загрязнение воздуха и изменение климата — все это оказывает свое негативное воздействие на биоразнообразие лесов. Эта деградация приводит к сокращению жизненного цикла лесов и отрицательно влияет на возможность человека использовать продукцию леса.
Девственные и естественные леса по сравнению с антропогенными лесами к большинству факторов обладают значительно большей устойчивостью. Девственные леса, сохранившиеся на относительно небольших площадях, нуждаются в охране как ценнейший генофонд лесов планеты. Всестороннее изучение девственных и естественных лесов как природных моделей устойчивых насаждений поможет уйти от массового шаблона в лесохозяйственной деятельности, нередко приводящего к необратимым ошибкам, которые оборачиваются миллионными убытками, пустыми затратами труда и времени, моральным ущербом и ущербом для лесных экосистем, который не всегда учитывается.
Задача приблизить по устойчивости антропогенные леса к естественным в целях снижения доли опеки (уходов, защиты и охраны) перед лесоводами ставилась еще Г Ф. Морозовым, который завещал вести дело так, чтобы постоянство пользования было налицо, но чтобы лес не терпел заметного ущерба в своей биологической устойчивости, по возможности был и самостоятелен в смысле возобновления и отличался от стихийного большей производительностью. «Наша задача, — говорил он, — ведет к умению сохранить в должной мере стихийное начало, дополнив его такими актами с нашей стороны, которые ослабляли бы вред от уменьшения подвижного равновесия».
7.Появление и развитие леса на ранее непокрытых лесом территориях, изменение густоты с возрастом
Появление леса на вырубках, сенокосах, бывших пашнях, гарях начинается с поселения. Иногда до сотен тыс/га. К возрасту 50-60 лет остается не более 1 тыс., к спелости - 300.
Объясение естественного изреживания насаждений. В динамике изреживания 10 тыс. растений погибают. В редких насаждениях погибают меньше. На единице площади помещается ограниченный объем фотосинтезирующего аппарата (объем хвои, масса хвои) – биоматерия содержится в пологе древостоя, движется в верх оставляя внизу погибшие ветки. Ослабленне деревья отмирают и количество их сокращается. По объему кроны деревья разделяются на 5 классов (ввел Крафт):
1.Наиболее развитые(10%)
2.Хорошо развитые(20-30%)
3.Стесненные – крона сдавлена сбоков, но вершина свободна(33%)
4.Угнетенные – крона до ½ полога(10-15%)
5.Заглушенная – крона под живыми деревьями(10-15%)
Распознать классы Крафта можно уже в самом раннем возрасте (8-9 лет) с вероятностью 60-70%, диагностировать будующих лидеров. Будующие аутсайдеры 90-96%
В этом возрасте формируется ранговая структура древостоя и неумолимо начинает действовать ранговый закон роста (По Маслакову): «Деревья просто с возрастом, увеличив свои размеры, начиная с 8-10 лет до 40, оставаясь в основном крупными или мелкими, средние деревья меняют свои ранги, как вверх, так и вниз.»
По этому закону, в раннем возрасте можно удалять все мелкие растения и оставлять не более 1 тыс. самых развитых. Из них вырастает лес с запасом в 2 раза больше чем без регулирования густоты.
ГУСТОТА ДРЕВОСТОЯ плотность стояния деревьев в древостое, характеризующаяся числом деревьев на единице площади (1 га) с учетом толщины стволов. От густоты древостоя зависят рост деревьев в высоту и по диаметру, развитие крон, очищение стволов от сучьев и др. Чем больше густота древостоя, тем интенсивнее отпад, меньше средние высота и диаметр деревьев. В естественных насаждениях первоначальная густота составляет несколько тысяч растений на 1 га. С возрастом густота древостоя. уменьшается и к спелости насаждения достигает нескольких сотен деревьев. Густота древостоя зависит от условий местопроизрастания. В спелых древостоях одинакового породного состава густота может быть меньшей в лучших лесорасти-тельных условиях и большей - в худших. Нормы густоты древостоя установлены для основных лесообразующих пород, групп типов леса и возрастных групп и приведены в таблицах хода роста древостоев. Древостой с густотой деревьев 25 % и менее от нормы можно считать редким, 26-50 % -изреженным, 51-75 % - средней густоты, 76—100 % - густым, 101 % и более - очень густым. Густоту древостоя чаще оценивают в молодом возрасте (до 10 лет) и при необходимости проводят мероприятия по дополнению или сохранению деревьев в редких и изреженных древостоях и разреживанию излишне густых древостоев для ускоренного роста перспективных деревьев. В таксации лесных массивов этот показатель используется редко (см. Густота защитного лесного насаждения).
8. Индивидуальное развитие дерева. Полог древостоя, крона дерева, классификация деревьев по Крафту. Ранговый закон Е.Л.Маслакова
Когда появляются леса, образуется полог. Полог – это пространство, занятое сомкнувшимися в горизонтальном и вертикальном направлениях кронами деревьев, образующими древостой. Деревья растут, и полог движется вверх, оставляя внизу на стволах отмирающие ветви. Ослабленные деревья также отмирают.
Крона дерева – это верхняя часть дерева, состоящая из ствола, ветвей и сучьев, в ней содержится фотосинтетический аппарат.
По объему кроны немецкий лесовод Крафт разделил живые деревья на 5 классов, которые так и называют «классы Крафта»:
1 – наиболее развитые (доминанты), примерно 10%;
2 – хорошо развитые (субдоминанты), около 30%;
3 – средние (крона сдавлена с боков, но вершина свободна), около 30–40 %;
4 – угнетенные (крона до ½ полога), 10–20%;
5 – заглушенные (крона под пологом), 10–15%.
Объем кроны – это багаж, с которым дерево движется в будущее. Распознать (диагностировать) классы Крафта можно уже в возрасте около 10 лет по размерам растения – высоте и диаметру стволика, объему хвои.
В возрасте около 10 лет формируется ранговая структура древостоя и начинает функционировать ранговый закон роста деревьев в древостое (Маслаков, 1984), в соответствии с которым начиная с этого возраста (8-10 лет) деревья растут, просто увеличивая свои размеры, оставаясь либо крупными, либо мелкими. Средние растения меняют свои ранги как вверх, так и вниз. Так, в групповых посадках сосны корреляционное отношение между площадями сечения деревьев в 10 и 40 лет составляет 0,88, а в 15 и 40 лет связь оказывается почти функциональной (Маслаков, 1984).
Зная этот закон, можно уже в самом раннем возрасте оставить только деревья-лидеры (с небольшим запасом), удалив остальные. Из них вырастет могучий и жизнестойкий лес с крупными деревьями и запасами древесины в 2–3 раза большими, чем средние естественные леса. При этом их старение отодвигается на десятки лет.
ОНТОГЕНЕЗ (от греч. происхождение возникновение) дерева, индивидуальное развитие дерева, имеющего семенное или вегетативное (поросль, корневища) происхождение, от зарождения до смерти. Поскольку средопреобразующее воздействие дерева зависит от того, в каком ярусе оно находится в данный момент онтогенез, для каждого онтогенетического состояния указан ярус.
Выделение ярусов проведено в соответствии с крупными подразделениями жизненных форм растений. Выделены следующие ярусы:
Проростки (p) - неветвящиеся растения, сформировавшиеся из семени в год его прорастания; имеют первичные корень и побег с семядолями, которые могут располагаться как над землей (у большинства растений), так и под землей (у дуба).
Ювенильные растения (j) обычно уже не имеют семядолей, но обладают детскими (инфантильными) структурами. Первичный побег (стволик) неветвящийся; листья ювенильной формы; корневая система состоит изглавного корня и небольшого числа боковых корней.
Проростки и ювенильные особи последовательно входят в состав яруса напочвенных мхов и лишайников (если онвыражен) и травяно-кустарничкового яруса; они характеризуются высокой теневыносливостью.
Имматурные растения (im) занимают промежуточное положение между ювенильными и взрослыми растениями. Побеговая система состоит из ветвей 2--4(5 го) порядков, крона еще не сформирована; общее число ветвей невелико, и диаметр стволика не более чем в 2 раза превышает диаметр крупных ветвей. Приросты стволика по длине и диаметру незначительно превышают приросты ветвей, в результате деревце имеет округлую или широкопирамидальную форму кроны.
Виргинильные деревья (v) имеют почти полностью сформированные черты взрослого дерева, но еще не приступили к семеношению. У них хорошо развиты ствол и крона, а прирост в высоту максимальный за весь онтогенез Величины текущего прироста ствола по длине значительно превышают таковые у крупных ветвей, что определяет удлиненную форму кроны с заостренной вершиной. Диаметр ствола превышает диаметр скелетных ветвей в 3 раза и более.
Молодые генеративные деревья (g1) имеют габитус взрослого и впервые приступают к семеношению. Органы семеношения локализованы в верхней части кроны, семян мало. Рост ствола в высоту интенсивный, порядок ветвления достигает 7--9 (10) и более. В нижней части ствола начинает формироваться корка.
Средневозрастные генеративные деревья (g2) имеют форму кроны -- от овальной или конусовидной с заостренной вершиной до округлой или тупо конусовидной.
Старые генеративные деревья (g3) практически прекращают рост в высоту, а прирост ствола по диаметру заметно уменьшается. Размеры кроны и корневой системы сокращаются из за отмирания части скелетных ветвей и якорных корней.
Сенильные деревья (s) у большинства видов имеют только вторичную крону, листья или хвоя могут быть ювенильного типа. Верхняя часть кроны и ствола отмирает, у лиственных деревьев и сосны часто остается живой нижняя половина или треть ствола, корневая система в значительной степени разрушена.
9. Рост сосны в ранние (4-10 лет) годы и влияние конкуренции (разного расстояния между растениями) на размеры стволов в 29-40 лет
Сосна обыкновенная является быстрорастущей породой. Максимальный прирост в высоту на лучших почвах наступает в возрасте 15—20 лет, на худших — в 25 лет. В возрасте 40—50 лет прирост в высоту замедляется, а затем и вовсе прекращается. По диаметру же дерево прирастает в течение всей жизни. Сосна доживает до 300—350 лет, редко до 400 лет и более.
В первые годы жизни на рост растений в основном влияют экологические факторы и эффекты материнского дерева , в особенности масса семени ,наблюдается интенсивная смена рангового положения высот растений в возрастном ряду (Маслаков,1980). Далее увеличивается влияние генетических факторов, наследуемость повышается, рост изучаемых растений стабилизируется и становится возможной оценка генетической обусловленности роста в возрасте 5-7 лет . Другие авторы отмечают стабилизацию роста после 10-15 лет и считают целесообразным начинать первые оценки скорости роста именно с этого возраста . В эти годы у сосны начинается период максимального роста, что и позволяет выделять лучшие потомства.
Наиболее детальные исследования роста деревьев сосны в молодняках проведены Е.Л. Маслаковым (1980, 1981, 1984, 1999), в результате которых был открыт «ранговый закон роста» деревьев в древостое. Согласно этому закону деревья с самого раннего возраста (8-10 лет) растут, в основном соблюдая сложившиеся в этом возрасте исходные пропорции.
Так, в 18 групповых посадках сосны корреляционное отношение между площадями сечения деревьев в 10 и 40 лет равно h = 0,88, а в 15 и 40 лет связь почти функциональна: h = 0,99. В других 15-летних культурах и в естественных молодняках такого же возраста деревья-лидеры определились уже в 5-7 летнем возрасте. Этот закон подтвердили исследования в республике Коми. В культурах закон проявляется сильнее. Так, в естественных молодняках только 30% деревьев сохранили свои ранги по высоте, тогда как в культурах их было уже 57%.Давно известно позитивное влияние сортировки посадочного материала. Размеры сеянцев влияли на рост сосны до 20-летнего возраста и отбор лучших растений повышал рост культур до 10%. В дальнейшем влияние сортировки, по-видимому, перекрывается действием ценотических факторов и не было прослежено.В селекции наиболее выгодны потомства со стабильным ростом.
10.Надежность диагностики в 4-10 лет лидеров и аутсайдеров у сосны
Можно отметить, в отличие от сосны, более точный прогноз развития для ели в условиях высокой начальной плотности ее ценозов, и раннюю диагностику ее роста начиная с 5-летнего возраста с выявлением деревьев-лидеров с надежностью 64% и с надежностью 70-75% в возрасте растений 7–10 лет.
Ранг 3 роста проявляется в редких или густых лесах. Если густота возрастает-размер уменьшается (размер корня тоже уменьшается).
Сосна 5-7 лет вероятность кроны – крупная 70-75%,МЕЛКАЯ 80-90%.
ОДНАКО за 30 лет вероятность увеличения кроны очень мала, коневая система сидит на полном уровне, корешки растут очень плотно.
Мерой соответствия и расхождения характера роста служили возрастные коэффициенты корреляции между размерами деревьев в 4, 5 7, 10 лет и их объемом в 29-40 лет. В молодом возрасте учитывали диаметр высоту и их произведение (Д2·Н), в старшем–объем ствола.
11. Ретроспекция роста ели: вероятность формирования крупных деревьев в возрасте 78 лет из крупных и мелких стволиков в 5-40 лет
диагностику роста дерева можно выяснить путем ретроспективного анализа ствола уже взрослого дерева вначале в самых простых сообществах древесных растений, какими являются лесные культуры. В качестве объекта для изучения данного вопроса желательны чистые высокопродуктивные лесные культуры, удовлетворяющие следующим требованиям:
- созданные одновозрастным посадочным материалом;
- с равномерным размещением посадочных мест;
- с максимальной сохранностью всех высаженных растений;
- в возрасте, приближающемся к возрасту рубки.
Связь между размерами стволов характеризует коэффициент корреляции, который закономерно увеличивался от r = 0,30 в 5 лет до r = 0,65-0,70 в 20 лет
Изучение роста модельных деревьев в крупном массиве культур ели, созданных посадкой и посевом семян в 1913 и 1919 г. по схемам Ф. А. Теплоухова, позволила установить следующее. В 70-78-летних культурах ели с высокой относительной полнотой (1,0-1,2) и густотой, достигавшей 2-5 тыс. шт./га в 20-30 лет, при разделении моделей в раннем возрасте по диаметру стволика на 3 и 6 градаций, можно дать следующие ретроспективные вероятности формирования из них деревьев с объемом ствола выше среднего (крупных деревьев). 1. Для растений с толстыми стволиками (15,6% количества растений в раннем возрасте) ретроспективные вероятности формирования из них крупных деревьев составят: в 5 лет 64%, в 7-12 лет 70-80%, в 20лет 70%, в 30 лет 80% и в 40 лет 100%. 2. Для растений с самыми толстыми стволиками (7,8% количества растений в раннем возрасте) указанные вероятности возрастают: в 5 лет 67%, в 7-12 лет 80-83%, в 20 лет 90%, в 30 лет и далее 100%. 3. Тонкие растения (до 60% количества растений в раннем возрасте) в любом возрасте, начиная с 5-7 лет, имеют мало шансов (менее 7%) сформировать крупные деревья, а самые тонкие (30% всех растений) их почти не имеют (ретроспективная вероятность формирования из них крупного дерева 2,7%). 4. Ранняя диагностика размеров деревьев у ели сибирской по отношению к их размерам в 70-78 лет в культурах возможна при следующих надежностях прогноза: - «мелкие» деревья (объем ствола ниже среднего) диагностируются в 5-7 лет по диаметру стволика менее 80% от среднего с надежностью прогноза 93-97%; - «крупные» деревья (объем ствола выше среднего) диагностируются в 7-10 лет по диаметру стволика 110% и более с надежностью прогноза 70-75% и улучшением прогноза до 85% при отборе по диаметру 125% от среднего и более.
12. Понятие о простых, сложных, равномерных и неравномерных древостоях. Показатели развития и их оценка. Понятие о полноте древостоя.
По форме древостой бывают простые и сложные. В простых древостоях кроны деревьев расположены в одном ярусе, т. е. высота деревьев относительно одинакова и колебания между высотами отдельных деревьев не превышают 10-15%. Такие древостой также называют одновысотными.
Сложными называют древостой, кроны деревьев которых расположены в двух или более ярусах, различающихся между собой по высоте, превышающей 10-15% от высоты большинства деревьев в каждом ярусе. Сложные древостой также называют многоярусными. Многоярусные древостой чаще бывают составлены из двух или нескольких пород, например, верхний (первый) ярус может образовывать сосна, нижний (второй) - ель. Сложную форму могут образовать и однопородные древостой, когда они составлены двумя или более возрастными группами, из которых более старая располагается в верхнем ярусе, а более молодая - в нижнем. Сложная форма древостоя объясняется не только ярусным строением, но и неравномерным размещением деревьев по площади в пределах участка.
Древостой с относительно равномерным размещением деревьев по площади участка относятся к простым, а с неравномерным, групповым размещением - к сложным формам древостоев.
Степень плотности стояния деревьев, характеризующую, в какой мере ими использованную занимаемое пространство, принято называть полнотой насаждения (заполненностью).
Так, если нет просветов между кронами – полнота 1,0; просветы занимают 20% – полнота 0,8; просветы занимают 30% – полнота 0,7 и т.д. >10-15 м, то деревья начинают раскачиваться, кроны трутся друг о друга, между ними появляются зазоры, и сомкнутость полога уже не годится для этого.
Поэтому для определения степени заполнённости ввели показатель абсолютной полноты, который стали измерять в виде общей суммы площади сечений деревьев на 1 га (∑g). 13.История и методика составления таблиц полных древостоев
В зависимости от древесной породы (светолюбивая, теневыносливая), условий роста (класса бонитета), размеров деревьев и состояния самого древостоя. Деревья светолюбивых пород по сравнению с теневыносливыми при прочих равных условиях образуют менее плотные древостои. При одних и тех же размерах деревьев насаждения при лучших условиях роста имеют на единице площади большее число стволов, чем растущие в худших условиях. Несмотря на то, что с увеличением возраста и размеров деревьев число их на единице площади уменьшается, сумма площадей их сечений и запас насаждения увеличиваются. Древостои, пройденные различными рубками (выборочными, санитарными, ухода), как правило, более изрежены, чем естественно формирующиеся.
Полнота — один из важнейших таксационных показателей, позволяющий определять запас насаждения, характеризовать его состояние и намечать хозяйственные мероприятия. Чтобы определить полноту древостоя, можно использовать данные о сумме площадей его сечений, запаса, числа деревьев на 1 га и степени сомкнутости полога. Полнота, устанавливаемая по сумме площадей сечений, называется таксационной и служит для определения запаса и прироста. Полнота определенная по степени сомкнутости крон – лесоводственной и служит для назначения хозяйственных мероприятий.
Различают абсолютную и относительную полноту. Абсолютная полнота насаждения выражается в квадратных метрах на 1-ra как общая сумма площадей сечений на высоте груди всех деревьев элемента леса или яруса (например, 32 м2) или как общая площадь горизонтальных проекций крон, образующих полог древостоя (например, 6000 м2).
В производственных условиях чаще определяют относительную полноту, выражаемую в десятых долях единицы. За единицу принимают полноту сомкнутого насаждения на 1 га, которая для данной породы, возраста (высоты) и условий местопроизрастания является максимальной. Такие насаждения называются нормальными. Согласно заданному условию нормальным считается насаждение, в котором в максимальной степени используются все природные возможности занимаемой им площади, т. е. такое, в котором нет лишних или недостающих деревьев. Если же в древостой без ущерба для окружающих деревьев можно добавить еще сколько-то, то полнота его окажется меньше 1,0. Так, к древостоям с полнотой 0,5 относятся те, в которых к уже имеющемуся количеству можно добавить еще столько таких же деревьев.
14. Стандартные таблицы полнот и запасов древостоев по лесорастительным зонам, относительная полнота
15.Закон развития одноярусных древостоев Г.С.Разина. Линии развития в зависимости от начальной густоты: а) сомкнутости крон б) суммарного объема крон
С первого года возникновения древостоя в нем происходит дифференциация деревьев по размерам – формируется их разнообразие по жизненному потенциалу. Причины этого процесса могут быть самые разные: сроки появления всходов, различия в возрасте, неравномерное расположение, микроусловия и др. Отстающие деревья постепенно отмирают – происходит самоизреживание древостоев. Каждое дерево появляется на свет с природной способностью расти и развиваться с максимальным использованием доставшихся ему индивидуальных особенностей и условий жизни в ценозе, развивая в первую очередь корневую систему и крону. Древостой, состоящий из множества разнообразных по жизненному потенциалу особей, приобретает интегральное свойство развиваться с полным использованием условий и стремится ускоренно осваивать доставшееся жизненное пространство. В этом проявляется стремление древостоя быстрее достичь индивидуальных пределов по всем таксационным показателям – линейным, площадным и объемным (Разин, Рогозин, 2012). Казалось бы, логика этих простых умозаключений вполне согласуется с идеями и законами экологии (Одум, 1986), однако ее законы не используются в полной мере в моделировании раз- вития древостоев. Рост древостоев весьма разнообразен, и впервые об изменении их роста сообщалось еще в начале XX в. Флюри и Гуттенбергом, ссылаясь на которых Н. В. Третьяков (1937) приводит пример, когда ель в 50 лет на трех участках имела высоту 13.5, 16.5 и 19.5 м, а в возрасте спелости таксировалась одним классом бонитета.
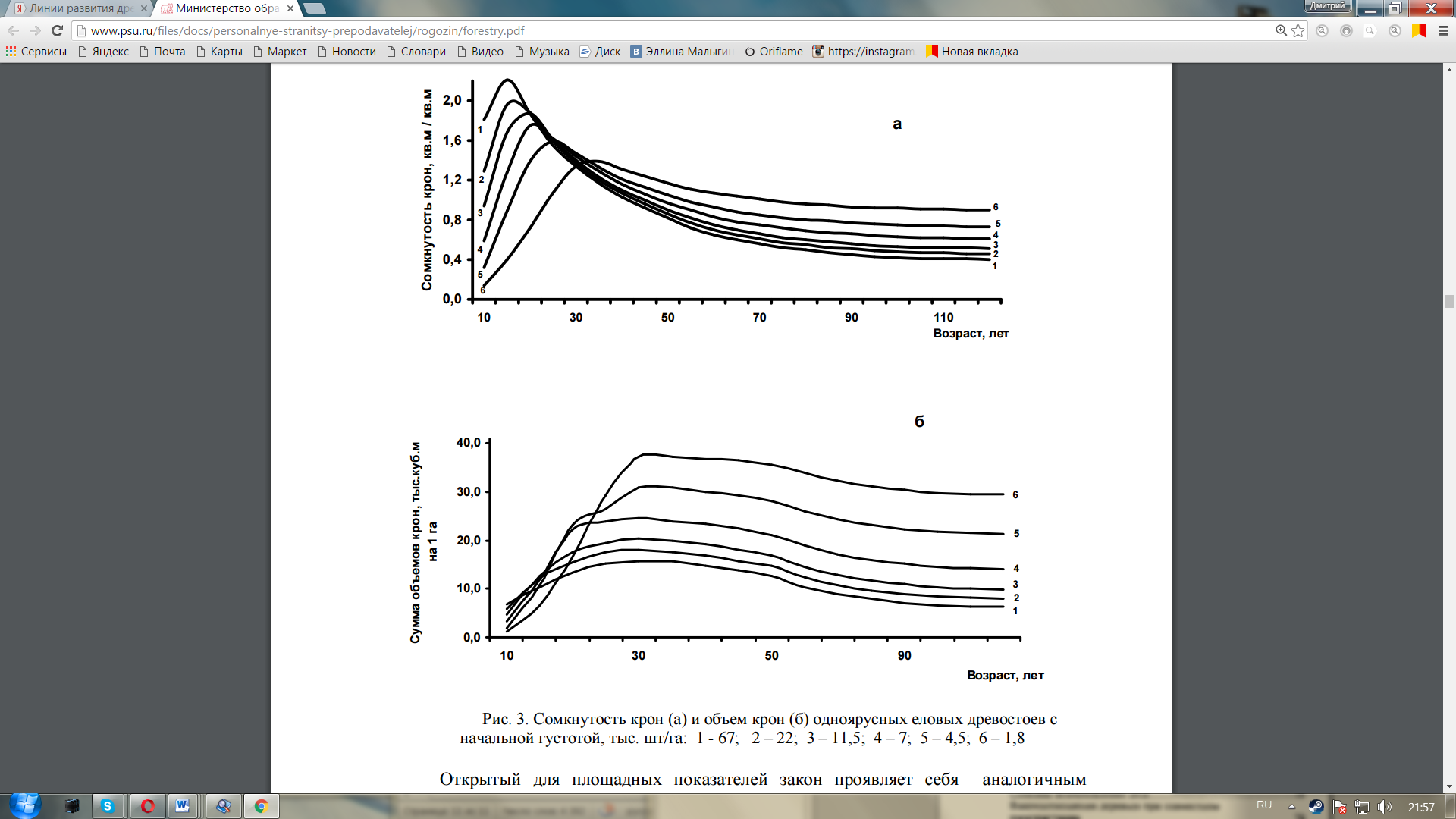
В серии работ (Разин, 1965, 1977, 1979, 1980, 1988, 2009), с их обобщением в статьях (Разин, Рогозин, 2009, 2010а, б) а также в настоящей книге применительно к древостоям культур показано, что рост, развитие и возрастная динамика древостоев подчиняются некоторым всеобщим биологическим законам. В древесных насаждениях эти законы имеют специфические особенности, которые заключаются в следующем.
1. Одноярусные древостои всех пород, составов, различной начальной густоты и неравномерности расположения деревьев имеют своей целью общее стремление всех деревьев яруса к достижению предельных линейных, площадных и объемных показателей в пределах сообщества. В результате почти любой древостой однажды в жизни становится сомкнутым и достигает индивидуального предельного состояния по этим показателям.
2. Достижение предельных показателей происходит в зависимости от густоты растений в сообществе и может быть сформулировано в виде «Закона динамики суммы горизонтальных проекций крон деревьев в простых древостоях», который состоит в следующем:
- чем больше начальная густота, тем раньше и при меньшей средней высоте древостои становятся сомкнутыми с предельной суммой площадей горизонтальных проекций крон деревьев и тем раньше и быстрее они снижают ее (размыкаются);
- чем меньше начальная густота, тем позже и при больших средних высотах древостои достигают предел сомкнутости и предел суммы площадей горизонтальных проекций крон деревьев, тем медленнее они снижают их и дольше остаются сомкнутыми и устойчивыми. Открытый для площадных показателей закон проявляет себя аналогичным образом в показателях объема. С некоторым запаздыванием по сравнению с сомкнутостью (на 10 –25 лет) древостои точно также достигают предельного состояния по сумме объемов крон и имеют индивидуальные линии своего развития по этому показателю в зависимости от начальной густоты (рис. 4.1б). Впечатляют полученные различия в суммах объемов крон, достигающие в крайних линиях 2-5 раз. Эти различия приводят к аналогичным различиям в объемах фотосинтезирующего аппарата, и если объемы крон отличаются в разы, то такими же будут различия ив объемах древесины.