

естественно
.PDF131
величину около 125 мВ. Мы говорим «около», «примерно» потому, что у клеток разного размера и типов этот потенциал может несколько отличаться, что связано с формой этих клеток (например, количеством отростков), а также с особенностями их мембран.
Все изложенное выше можно формально описать следующим образом. В покое клетка ведет себя как «калиевый электрод», а при возбуждении - как «натриевый электрод». Однако после того как потенциал на мембране достигнет своего максимального значения +55 мВ, натриевый ионный канал со стороны, обращенной в цитоплазму, закупоривается специальной белковой молекулой. Это так называемая «натриевая инактивация»; она наступает примерно через 0,5-1 мс и не зависит от потенциала на мембране. Мембрана становится непроницаемой для натриевых ионов. Для того чтобы потенциал мембраны вернулся к исходному состоянию-состоянию покоя, необходимо, чтобы из клетки выходил ток положительных частиц. Такими частицами в нейронах являются ионы калия. Они начинают выходить через открытые калиевые каналы. Вспомните, что в клетке в состоянии покоя накапливаются ионы калия, поэтому при открывании калиевых каналов эти ионы покидают нейрон, возвращая мембранный потенциал к исходному уровню (уровню покоя). В результате этих процессов мембрана нейрона возвращается к состоянию покоя (-70 мВ) и нейрон готовится к следующему акту возбуждения.
Таким образом, выражением возбуждения нейрона является генерация на мембране нейрона потенциала действия. Его длительность в нервных клетках составляет величину около 1/1000с(1 мс). Описанная последовательность событий приведена на рис.
Выполнению функций нейрона способствует синтез в его аксоплазме веществ-передатчиков — нейромедиаторов (нейротрансмиттеров): ацетилхолина, катехоламинов и др. Размеры нейронов колеблются от 6 до 120 мкм.
Число нейронов мозга человека приближается к 1011. На одном нейроне может быть до 10 000 синапсов. Если только эти элементы считать ячейками хранения информации, то можно прийти к выводу, что нервная система может хранить
132
1019 ед. информации, т. е. способна вместить практически все знания, накопленные человечеством. Поэтому вполне обоснованным является представление, что человеческий мозг в течение жизни запоминает все происходящее в организме и при его общении со средой. Однако мозг не может извлекать из памяти всю информацию, которая в нем хранится.
Для различных структур мозга характерны определенные типы нейронной организации. Нейроны, организующие единую функцию, образуют так называемые группы, популяции, ансамбли, колонки, ядра. В коре большого мозга, мозжечке нейроны формируют слои клеток. Каждый слой имеет свою специфическую функцию.
Клеточные скопления образуют серое вещество мозга. Между ядрами, группами клеток и между отдельными клетками проходят миелинизированные или немиелинизированные волокна: аксоны и дендриты.
Клеточные скопления образуют серое вещество мозга. Между ядрами, группами клеток и между отдельными клетками проходят миелинизированные или немиелинизированные волокна: аксоны и дендриты.
Одно нервное волокно из нижележащих структур мозга в коре разветвляется на нейроны, занимающие объем 0,1 ммЗ, т. е. одно нервное волокно может возбудить до 5000 нейронов. В постнатальном развитии происходят определенные изменения в плотности расположения нейронов, их объема, ветвления дендритов.
3.2.3. Глия
Помимо нейронов нервная ткань содержит клетки еще одного типа - клетки глии. Они выполняют опорную и защитную функции , а также в нейронофагии и называются глиальными клетками или глией. По численности их в 10 раз больше, чем нейронов , и они занимают половину объема ЦНС. Глиальные клетки окружают нервные клетки и играют вспомогательную роль. Глия не только выполняет опорные функции, но и обеспечивает многообразные метаболические процессы в нервной ткани, участвует в формировании миелиновой обо-
133
лочки и способствует восстановлению нервной ткани после травм и инфекций.
Между нейронами и глиальными клетками существуют сообщающиеся между собой щели размером 15-20 нм, так называемое интерстициальное пространство , занимающее 1214% общего объема мозга.
Глиальные клетки невозбудимы, во время деполяризации глиальных клеток проводимость их мембран не повышается.
Клетки нейроглии делятся на несколько типов. Клетки эпендимы выстилают желудочки головного мозга и спинномозговой канал и образуют эпителиальный слой в сосудистом сплетении . Они соединяют желудочки с нижележащими тканями.
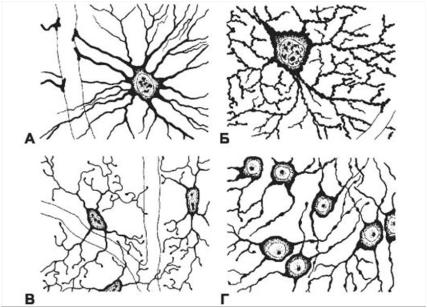
Клетки макроглии делятся на две категории - астроциты и олигодендроциты (рис. 30).
Протоплазматические астроциты локализованы в сером веществе ; от тела клетки, содержащей овальное ядро и большое количество гликогена , отходят сильно разветвленные короткие и толстые отростки.
Фибриллярные астроциты локализованы в белом веществе . Ядро у них также овальное, и тело клетки содержит много гликогена , но отростки длинные и менее разветвленные, некоторые ветви буквально упираются в стенки кровеносных сосудов. Эти клетки переносят питательные вещества из крови в нейроны.
Астроциты двух типов взаимосвязаны и образуют обширное трехмерное пространство, в которое погружены нейроны. Они часто делятся, образуя в случае повреждений центральной нервной ситемы рубцовую ткань.
Олигодендроциты локализованы в сером и белом веществе. Они мельче астроцитов и содержат одно сферическое ядро. От тела клетки отходит небольшое число тонких веточек, а само оно содержит цитоплазму с большим количеством рибосом. Шванновские клетки -это специализированные олигодендроциты, синтезирующие миелиновую оболочку миелинизированных волокон .
Клетки микроглии локализованы и в сером, и в белом
134
Рис. 30. Глия. Виды глии.
А — волокнистый астроцит; Б — протоплазматический астроцит; В — микроглия; Г — олигодендроглиоциты. (Doctor- V.ru)
веществе, но в сером веществе их больше. От каждого конца маленького продолговатого тела клетки, содержащей лизосомы и хорошо развитый аппарат Гольджи , отходит по толстому отростку. От всех его ветвей отходят более мелкие боковые веточки. При повреждении мозга эти клетки превращаются в фагоциты и, перемещаясь при помощи амебоидного движения, противостоят вторжению чужеродных частиц.
Глия является системой трофического обеспечения нервной системы, а также принимает активное участие в специфическом функционировании нервной ткани: в норме тормозит гиперактивность нейронов, способствует активному поглощению из синаптической щели и утилизации медиаторов и других агентов, участвующих в повреждении нейронов. В условиях ишемии микроглиальные клетки индуцируют синтез не только нейротоксичных веществ, но и сигнальных молекул, клеточных регуляторов, трофических факторов, способствующих выживаемости нейронов и уменьшающих процессы постишемического рубцевания
Микроглия - единственный иммунокомпетентный компартамент в центральной нервной системе.
135
3.2.4. Стволовые клетки
Завершая свою книгу "Дегенерация и регенерация нервной системы", Рамон - и - Кахал писал: "Во взрослом мозге нервные пути -это нечто фиксированное, законченное, неизменяемое. Все может погибнуть, ничто не может регенерировать". В течение многих лет это положение было одной из общепринятых основ нейробиологии; Однако в 90 - х г.г. многочисленными исследователями было показано на грызунах наличие во взрослом мозге клеток - предшественников (в основном дифференцирующихся в гранулярные клетки), мигрирующих в различные отделы мозга в течение всей жизни.
В1981 году американский биолог Мартин Эванс впервые выделил недифференцированные плюрипотентные линии стволовых клеток— бластоцисты мыши. В 1998 году Д. Томпсон и Д. Герхарт выявили бессмертную линию эмбриональных стволовых клеток (ЭСК).
В1999 году журнал Science признал открытие эмбриональных стволовых клеток третьим по значимости событием в биологии после расшифровки двойной спирали ДНК и про-
граммы «Геном человека». (http://pO3 .org.ru/publ/l-1 -0-28)
Стволовые клетки — иерархия особых клеток живых организмов, каждая из которых способна впоследствии изменяться (дифференцироваться) особым образом (то есть получать специализацию и далее развиваться как обычная клетка). Стволовые клетки способны асимметрично делиться, из-за чего при делении образуется клетка, подобная материнской (самовоспроизведение), а также новая клетка, которая способна дифференцироваться.
Термин «стволовая клетка» был введён в научный обиход русским гистологом Александром Максимовым (1874—1928). Он постулировал существование стволовой кроветворной клетки. На заседании Общества Гематологов в Берлине 1 июня 1909 года он ввел понятие «Stammzelle», подразумевая под этим определением лимфоцит в более широком значении этого слова, как клетку, способную быть стволовой в современном понимании этого слова.
В60-е годы прошлого столетия Тил и Маккулох, а
136
также Меткаф и его сотрудники показали, что внутривенное введение костномозговых клеток от здоровой сингенной к летально облученной мыши приводит к образованию колоний из клеток всех направлений гемопоэтической дифференцировки в селезенке. С разработкой клонального метода для выявления клеток предшественников in-vitro, так называемых колониеобразующих единиц (КОЕ), стало возможным проследить за дифференцировкой всех миелоидных ростков.
Фриденштейн А. Я. и его сотрудники впервые показали, что в костном мозге, помимо гемопоэтических имеются стромальные стволовые клетки, которые при культивировании формировали колонии фибробластноподобных клеток. Пересадка таких колоний под капсулу почки мыши в диффузион-
Рис. 31. Стволовые клетки.
ной камере приводило к формированию костной или адипозной ткани.
Корнем иерархии стволовых клеток является тотипотентная зигота. Первые несколько делений зиготы сохраняют тотипотентность и при потере целостности зародыша это может приводить к появлению монозиготных близнецов. К ветвям иерархии относятся плюрипотентные (омнипотентные) и мультипотентные (бластные) стволовые клетки. Листьями (конечными элементами) иерархии являются зрелые унипотентные клетки тканей организма.
137
Нишами стволовых клеток называются места в ткани, где постоянно залегают стволовые клетки, делящиеся по мере надобности для дальнейшей дифференциации.
Стволовые клетки размножаются путём деления, как и все остальные клетки. Отличие стволовых клеток состоит в том, что они могут делиться неограниченно, а зрелые клетки обычно имеют ограниченное количество циклов деления.
Когда происходит созревание стволовых клеток, то они проходят несколько стадий. В результате, в организме имеется ряд популяций стволовых клеток различной степени зрелости. В нормальном состоянии, чем более зрелой является клетка, тем меньше вероятность того, что она сможет превратиться в клетку другого типа.
Но всё же это возможно благодаря феномену трансдифференцировки клеток (англ. Trans differentiation).
ДНК во всех клетках одного организма (кроме половых), в том числе и стволовых, одинакова. Клетки различных органов и тканей, например, клетки кости и нервные клетки, различаются только тем, какие гены у них включены, а какие выключены, то есть регулированием экспрессии генов, например, путем метилирования ДНК. Фактически, с осознанием существования зрелых и незрелых клеток был обнаружен новый уровень управления клетками. То есть, геном у всех клеток идентичен, но режим работы, в котором он находится— различен.
В различных органах и тканях взрослого организма существуют частично созревшие стволовые клетки, готовые быстро дозреть и превратиться в клетки нужного типа. Они называются бластными клетками. Например, частично созревшие клетки мозга— это нейробласты, кости — остеобласты и так далее. Дифференцировку могут запускать как внутренние причины, так и внешние. Любая клетка реагирует на внешние раздражители, в том числе и на специальные сигналы цитокины. Например, есть сигнал (вещество), служащий признаком перенаселённости. Если клеток становится очень много, то этот сигнал сдерживает деление. В ответ на сигналы клетка может регулировать экспрессию генов.
Характеристики эмбриональных стволовых клеток
138
1.Тотипотентность— способность образовывать любую из 350 тканей организма;
2.Хоуминг— способность стволовых клеток, при введении их в организм, находить зону повреждения и фиксироваться там, исполняя утраченную функцию;
Факторы, которые определяют уникальность стволовых клеток находятся не в ядре, а в цитоплазме. Это избыток мРНК всех 3 тысяч генов, которые отвечают за раннее развитие зародыша;
Теломеразная активность.
При каждой репликации часть теломер утрачивается (лимит Хейфлика или биочасы). В стволовых, половых и опухолевых клетках есть теломеразная активность, концы их хромосом надстраиваются, то есть эти клетки способны проходить потенциально бесконечное количество клеточных делений, они бессмертны. Стволовых клеток в нашем организме очень мало:
• у эмбриона — 1 клетка на 10 тысяч,
• у человека в 60-80 лет — 1 клетка на 5-8 миллионов. В мозге стволовые клетки спонтанно дифференцируют-
ся в три основных типа клеточных элементов ЦНС - нейроны, астроциты и олигодендроглию. В последние годы стали интенсивно изучаться свойства различных типов стволовых клеток и возможность их применения в медицине, в частности при болезнях ЦНС.
На моделях травмы спинного мозга (ТСМ) было показано, что нейрональные региональные стволовые клетки могут выживать, интегрироваться с мозгом хозяина, а также и дифференцироваться в нейроны и глию. Развитие и дифференцировка трансплантированных нейрональных стволовых клеток (выделенных из эпендимиальных оболочек) совпадала с восстановлением функции экспериментальных животных с ТСМ. Было высказано предположение, что плюрипотентные свойства неирональных стволовых клеток позволяют использовать их в качестве источника донорских клеток для трансплантации при травме спинного мозга.
Vacanti С.А также использовал региональные нейрональные стволовые клетки, выделенные из спинного мозга
139
крысы. Клетки пересаживались в геле из плюроника Р-200, при этом трансплантат во время операции закрыл диастаз перерезанного спинного мозга длинной 4 мм. В группе животных с трансплантацией неирональных стволовых клеток наступило восстановление функции задних конечностей, тогда как в контрольной группе существенных изменений не произошло. Из линии клеток, человеческой тератокарциномы (hNT клетки от Layton Bioscience Inc) после воздействия ретиноевой кислотой была получена гомогенная п опуляция неирональных клеток предшественников NT2N. Это клетки (NT2N) характеризовались стабильностью нейрохимических, физиологических и морфологических свойств в культуре. NT2N клетки были успешно пересажены в мозг иммунодефицитных мышей, хорошо прижились, не образовывали опухоли, не отторгались, не некротизировались и не подверглись апоптозу в течение одного года.
Более того, было показано, что пересаженные NT2N клетки, интегрировали с окружающей нейрональной тканью хозяина, посредством дендритных и аксональных отростков. Эти клетки были использованы на животных моделях, имитирующих следующие неврологические состояния: инсульт, болезнь Паркинсона и нейротравма. Недавно, показано, что трансплантированные NT2N клетки интегрируют с клетками хозяина в спинном мозге мыши и дают спрутинг аксонов на протяжении более чем 2 см. Использование в клинике NT2N клеток сдерживается потенциальной возможностью малигнизации этих клеток.
Другая нейрональная линия стволовых клеток RN33B была получена из ядер шва эмбрионального мозга крысы. Эти клетки были трансфецированы ретровирусным вектором, несущим ген, кодирующим температурно чувствительный мутантный протеин SV40 большого Т-антигена. На модели ушиба спинного мозга крысы было продемонстрировано, что пересаженные после трансфекции клетки развиваются и дифференцируются в биполярные нейроны, а также интегрируют с окружающей тканью спинного мозга хозяина. К сожалению подобных попыток применения этих клеток в клинике не было из-за развития хромосомных аберраций в экспе-
140
рименте. McDonald и соавторы использовали линию эмбриональных стволовых клеток D3, обработанных ретиноевой кислотой, для трансплантации в область повреждения спинного мозга крыс. Перед трансплантацией клетки были трансфецированы LacZ, экспрессирующим гамма-галактозидазу. Спустя две недели, методом двойного окрашивания были обнаружены пересаженные клетки. При этом было показано, что трансплантированные клетки окрашиваются также специфическими маркерами на нейроны (NeuN - нейрон-специфический ядерный протеин), астроциты (GFAP- глиально-фибрилярный кислый протеин), олигоденроциты (АРС СС-1 аденоматозный полипозный генетический продукт палочек). В группе животных с трансплантацией клеток было выявлено некоторое улучшение функции паретичных конечностей по сравнению с контролем.
Клоны стволовых клеток можно поддерживать in vitro до двух лет. Новые нейроны, мигрирующие в структуры взрослого мозга, посылают аксоны к надлежащим м и- шеням, окружены функциональными синапсами и не отличаются от окружающих клеток, включаясь, таким образом, в существующие системы нейронов.
Нейрогенез во взрослом мозге (анализируемый обычно на гранулярных клетках зубчатой фасции) может возрастать под влиянием различных факторов - как при патологических условиях, так и при повышении нормальной активности мозга. Показано увеличение генерации новых гранулярных клеток через 1 - 2 недели после 10 мин глобальной ишемии мозга, что рассматривается как компенсаторная реакция. При судорожной активности, вызванной посредством киндлинга, введения каиновой кислоты или пилокарпина, наблюдается повышение нейрогенеза в зубчатой фасции на 75 - 140 % (по другим данным - в 6 раз). У старых животных, в мозге которых наблюдается повышенное содержание кортикостерона, снижение кортикостерона приводит к усилению нейрогенеза.
Вместе с тем, убедительные данные о повышении нейрогенеза у взрослых и старых животных получены на мышах, содержащихся в обогащенной среде (включающей социальные взаимодействия, объекты, вызывающие усиление ориен-