

естественно
.PDF141
тировочно - исследовательской реакции, и условия для физической активности). Перемещение животных в такую среду вызывает усиление пролиферации нейронов вдвое у молодых взрослых крыс. Происходит также возрастание пролиферации астроцитов. Такое усиление продукции нейронов наблюдается даже у старых крыс, переводимых в обогащенную среду из стандартных условий в возрасте 20 мес, причем число пролиферирующих нейронов у них возрастает в 3 раза (по отношению к сниженному в этом возрасте фоновому уровню). Это сопровождается умеренным, но достоверным улучшением обучения в водном лабиринте Морриса (Виноградова О.С,).
3.2.5. Синапс
Вконце прошлого века Ч.Шеррингтон обосновал представление об отсутствии межклеточной непрерывности в нервной системе и ввел понятие синапс для обозначения структуры, которая опосредует передачу сигнала от окончаний аксона к эффекторной клетке — нейрону, мышечному волокну, секреторной клетке железы. Синапс состоит из пресинаптического окончания и постсинаптической мембраны, разделенных синоптической щелью, которая заполнена рыхлым коллагеноподобным веществом.
Существуют два способа синаптической передачи — электрический и химический.
Возможно и сочетание обоих механизмов, электрического
ихимического, в одном смешанном синапсе, однако в нервной системе млекопитаюших преобладают чисто химические синапсы.
Вэлектрических синапсах, количество которых в нервной системе относительно невелико, потенциал действия пресинаптических окончаний обеспечивает ток, который деполяризует постсинаптическую мембрану. Морфологическую основу электрической передачи составляет щелевой контакт, для которого характерны тесное прилегание пре- и постсинаптической мембран (ширина синаптической щели всего лишь 2 нм), большая площадь контакта этих мембран, наличие ультраструктур, снижающих электрическое сопротивление в области контакта, —
142
своего рода каналов, организованных в виде правильной сети между пре- и постсинаптической мембраной.
Электрофизиологическими критериями электрической синаптической передачи являются: 1) отсутствие синаптической задержки; 2) проведение возбуждения в обоих направлениях (хотя некоторые электрические синапсы обладают выпрямляющими свойствами, т.е. коэффициент связи от пресинаптической мембраны к постсинаптическому нейрону больше, чем в обратном направлении); 3) независимость от потенциала пресинаптической мембраны; 4) устойчивость к изменениям концентрации ионов кальция и магния в среде, к асфиксии, низкой температуре, некоторым фармакологическим воздействиям.
Функциональная роль электрических синапсов состоит в осуществлении срочной передачи сигналов (без синаптической задержки), обеспечивающей синхронизацию электрической активности группы нейронов, например группы мотонейронов во время прыжковых движений лягушки или плавательных движений рыбы. Электрические синапсы обнаруживаются между нервными клетками, однотипными по структуре и функциям.
Эволюция нервной системы сопровождается уменьшением числа электрических синапсов в пользу другого способа передачи
— химического. В химическом синапсе, нервный импульс вызывает освобождение из пресинаптических окончаний химического посредника — нейромедиатора, который диффундирует через синаптическую щель (имеющую ширину 10-50 нм) и вступает во взаимодействие с белками-рецепторами постсинаптической мембраны. В результате происходит генерация постсинаптического потенциала.
Химический механизм синаптической передачи по сравнению с электрическим более эффективно обеспечивает основные функции синапса: 1) одностороннее проведение сигнала; 2) усиление сигнала; 3) конвергенцию многих сигналов на одной постсинаптической клетке, пластичность передачи сигналов (обучение, память и т.д.),
Химические синапсы передают два вида сигналов — возбуждающий и тормозной (рис. 32). В возбуждающих синапсах нейромедиатор, освобождаемый из пресинаптических нервных
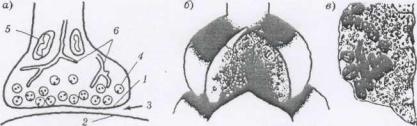
143
Рис. 32. Различные варианты изображения синапса: а — плоская (двухмерная) схема; б — объемная схема; в — фотография, полученная с помощью электронного микроскопа; 1 — пресинаптическая мембрана; 2 — постсинаптиче-ская мембрана; 3
— синаптическая щель; 4 — везикулы с медиатором; 5 — митохондрии; 6 — эндоплазматическая сеть
окончаний, вызывает в постсинаптической мембране возбуждающий постсинаптический потенциал — локальную деполяризацию, а в тормозных синапсах — тормозной постсинаптнгческий потенциал, как правило, —гиперполяризацию. Снижение сопротивления мембраны, происходящее во время тормозного постсинаптического потенциала, ведет к короткому замыканию возбуждающею постсинаптического тока, тем самым ослабляя или блокируя передачу возбуждения.
В течение многих десятилетий синапс рассматривался как, если и не единственная, то абсолютно доминирующая в нормальных условиях форма коммуникации между нейронами. Эта догма подверглась пересмотру в последние годы. Заметим попутно, что окончательно завершен и начатый еще Гольджи и Рамон-и-Кахалом спор о дискретности синапса. Электронная микроскопия и выделение синаптосом окончательно показали, что синаптическая щель - не пустое пространство, а сложная конструкция, соединяющая пре - и постсинаптический элемент. Однако, помимо локальных синаптических взаимодействий выявлены и множественные системы коммуникаций между нейронами. а) Объемное распространение трансмиттера
(volume conduction, spill - over, cross - talk, paracrine transmission). б) Щелевые контакты, формируемые прямой аппозицией олигомерных трансмембранных белков, в) Регуляция локального кровотока.
Синапс настолько узок, что его строение можно изучать только в электронный микроскоп. Он состоит из пресинапти-
144
ческой части (отросток, или тело нейрона) и постсинаптической части (отросток, или тело). Цитоплазма в месте контакта уплотнена с обеих сторон или только в постсинаптической клетке. Сигнал передается от пресинаптическои части к постсинаптической. Между ними находится синаптическая щель шириной 0,02—0,03 мкм. Диаметр синапса 1—2 мкм и менее
(рис. 32).
В пресинаптическом окончании находятся небольшие мембранные пузырьки — везикулы. Диаметр везикул может составлять 0,02—0,06 мкм и более; их форма сферическая или уплощенная. Везикулы наполнены физиологически активными веществами — медиаторами. Для каждого конкретного нейрона параметры образуемых им синапсов (размер щели, диаметр и форма везикул, количество молекул медиатора в везикуле) постоянны.
Возбуждение (электрический потенциал действия) нейрона в синапсе превращается из электрического импульса в импульс химический, контактирует с дендритами или телами сразу нескольких нейронов.
Иная картина при формировании так называемых дендритных шипиков: пресинаптических окончаний несколько, постсинаптическая же Другими словами, каждое возбуждение нейрона сопровождается выбросом в окончании его аксона порции биологически активного вещества - медиатора. Далее молекулы медиатора связываются с специальными белковыми молекулами, которые находятся на мембране другого нейрона. Эти молекулы называются рецепторами. Рецепторы устроены уникально и связывают только один тип молекул. В некоторых описаниях указывается, что они подходят, как «ключ к замку» (ключ подходит только к своему замку).
Рецептор состоит из двух частей. Одну можно назвать «узнающим центром», другую - «ионным каналом». Если молекулы медиатора заняли определенные места (узнающий центр) на молекуле рецептора, то ионный канал открывается и ионы начинают входить в клетку (ионы натрия) или выходить (ионы калия) из клетки. Другими словами, через мембрану протекает ионный ток, который вызывает изменение потенциала на мембране. Этот потенциал получил название постсинаптиче-
145
ского потенциала. Очень важным свойством описанных ионных каналов является то, что количество открытых каналов определяется количеством связанных молекул медиатора, а не потенциалом на мембране, как в случае с электровозбудимой мембраной нервного волокна. Таким образом, постсинаптические потенциалы имеют свойство градуальности: амплитуда потенциала определяется количеством молекул медиатора, связанного рецепторами. Благодаря этой зависимости амплитуда потенциала на мембране нейрона развивается пропорционально количеству открытых каналов.
С конца 70 - х г.г. появились многочисленные работы, показывающие сосуществование в нейронах и синапсах всех видов "классических" трансмиттеров с различными пептидами, обнаруженными первоначально как в самой нервной ткани, так и в других тканях организма. Тогда же было обнаружено активное участие пептидов в модуляции синаптической передачи. В 1986 г Хёкфельт с 19-тью соавторами представил сводный обзор по всей опубликованной к этому времени литературе с таблицами, показывающими ко-локализацию с трансмиттерами энкефалина, холецистокинина, нейропептида Y, вещества Р, соматостатина, мотилина, галанина, нейротензина и др. Хотя различные нейропептиды, иногда не один, а более, могут ко - локализоваться со всеми трансмиттерами, максимальное их разнообразие наблюдается в ГАМКергических клетках, причем их группы, различающиеся по морфологии и топографии (например, в гиппокампе), могут содержать разные нейропептиды.
Возможность выделения одним синапсом разных активных веществ при различных характеристиках входного сигнала была впервые прямо показана на мотонейроне В15 аплазии: при низком уровне возбуждения он работает как чисто - холинергический нейрон, а при повышении частоты разрядов как холинергический - пептидергический. В дальнейшем аналогичные частото-зависимые эффекты были показаны для синапсов гиппокампа и супрахиазмального ядра. Это привело к общему заключению, что одиночный нейрон представляет собою гибкую систему с вариабельными трансмиттерами, высвобождаемыми в разных сочетаниях при разных комбинациях воздейст-
146
вующих сигналов, причем допускается и выход разных сигнальных веществ на разных синапсах одного нейрона.
Наиболее распространены аксо-дендритные и аксосоматические синапсы — контакты между аксоном одной клетки (пресинаптическая часть) и дендритом либо телом другой клетки (постсинаптическая часть). Встречаются и другие варианты: дендро-дендритные, сомато-дендритные, аксоаксональные и прочие типы синапсов.
Синапс, состоящий из одного пре- и одного постсинаптического окончания, называют простым, но в ЦНС существуют и сложные синапсы. В этом случае можно наблюдать, как пресинаптическое окончание аксона образует «розетку» мембранных выростов и часть одна и образована характерным выростом дендрита.
Более сложную структуру имеют синаптические гломерулы — компактные скопления нервных отростков разных клеток, формирующие большое количество взаимных синапсов. Обычно такие гломерулы окружены оболочкой из глиальных клеток.
Синапс является элементарной структурой и функциональной единицей нервной ткани, в которой происходит передача сигналов с нейрона на нейрон. Взаимодействие эффектов рядом расположенных синапсов — один из важнейших элементов обработки информации в ЦНС, а существование сложных синапсов (особенно синаптических гломерул) позволяет осуществлять этот процесс особенно эффективно. Наибольшее количество сложных синапсов расположено именно в тех зонах мозга, где происходит максимально сложная обработка сигналов — в коре больших полушарий, коре мозжечка и таламусе.
Количество синапсов, образуемых одним нейроном, в среднем составляет 2—5 и более тысяч. Синапсы присутствуют на теле нейрона, его дендритах и в меньшей степени на аксоне. Наибольшее значение для деятельности нервных клеток имеют контакты с телом нейрона, основанием дендритов, а также точками их первого разветвления. Пресинаптическую функцию выполняют чаще всего конечные разветвления аксонов, реже — их коллатерали, а также тонкие ветвления денд-
147
ритов.
Отдельные нейроны, соединяясь синаптическими контактами, образуют нервные цепи и сети, состоящие из трех ос-
новных элементов — входных волокон, релейных клеток (тип Гольджи I — более крупные, обычно возбуждающие; имеют длинные аксоны и дают эфферентные выходы из сети к другим структурам мозга) и интернейронов (тип Гольджи II — мельче, чаще тормозные; имеют короткие аксоны и модулируют активность близлежащих нервных клеток). Каждый синапс может быть образован любыми двумя из этих элементов или всеми тремя. При этом формируются нейронные контуры, способные осуществлять передачу сигналов по сложным траекториям; соединять и разделять потоки сигналов. Такие сети образуют простейшие (типовые) нейронные модули, служащие для обработки информации, которые функционируют как единое целое и могут многократно повторяться в некоторой мозговой структуре (ядре).
Кроме синапсов в ЦНС, нервные клетки способны к образованию контактов с мышцами и внутренними органами более крупных структур размером до 50 мкм. Нервномышечные синапсы — это синапсы двигательной системы организма, срабатывание такого контакта вызывает сокращение мышечного волокна. На каждой поперечно-полосатой мышечной клетке имеется только один нервно-мышечный синапс, образованный мотонейроном ЦНС.
Синапсы с внутренними органами — это синапсы вегетативной нервной системы. Их образуют симпатические и парасимпатические нейроны с клетками гладких мышц или желез. Срабатывание таких контактов вызывает изменение секреции или тонуса мышечных волокон в стенках внутренних органов. В вегетативной нервной системе существуют синапсы между нервными клетками вне ЦНС •— в нервных сплетениях и ганглиях.
В последние годы произошла ревизия представлений о локальном действии трансмиттера в синапсе, являющемся его источником. При выбросе трансмиттера количество его молекул избыточно и не может быть связано внутрисинаптическими рецепторами. Лишь ограниченное число контактов в мозге
148
представлено синапсами "закрытого типа" (гломерулярные синапсы, контакты, изолированные глиальными отростками). В них избыточный трансмиттер убирается посредством обратного захвата, при действии транспортеров и метаболических ферментов. Большинство синапсов представляет собою открытые контакты, из которых трансмиттер может распространяться как трехмерный диффузионный сигнал по изотропной пористой среде. Эффективность этой передачи определяется существованием внесинаптических рецепторов на сома - дендритной мембране нейронов. Эта передача может быть близкой (но более величины синаптической щели), захватывая дополнительные внесинаптические рецепторы своего и прилежащих нейронов, и дальней - при диффузии через внеклеточную среду и ликвор. Функциональное значение этого типа передачи подчеркивается и тем фактом, что трансмиттеры разных типов могут выделяться аксонными варикозами, вообще не имеющими синаптических специализаций. Такая диффузная объемная передача показана для холинергических, серотонинергических, дофаминергических волокон (в последних 70 % варикоз - асинаптические, но способны выделять трансмиттер).
б) Щелевые контакты, формируемые прямой аппозицией олигомерных трансмембранных белков, позволяют непосредственный обмен ионами и малыми молекулами, представляя, таким образом, дополнительную систему метаболических
иэлектрических коммуникаций между нейронами. Эти контакты особенно важны и многочисленны в развивающейся нервной ткани, но во многих структурах они широко представлены
иу взрослых животных. Щелевые контакты имеют высоко - специализированную молекулярную структуру, основным элементом которой являются особые белки - коннексины. Меняя электрический потенциал клеток, щелевые контакты могут влиять на синаптические эффекты. Вместе с тем, сейчас накапливаются данные, что нейротрансмиттеры (ацетилхолин, дофамин, норадреналин, серотонин, глутамат), выделяемые из химических синапсов, могут регулировать состояние щелевых контактов (в основном снижая их проводимость) через различные системы вторичных посредников. Эта форма связи участвует в объединении нейронов, а также нейронов и астроцитов в
149
функциональные ансамбли, ее генетические и иные нарушения могут приводить к тяжелым формам патологии нервной системы.
в) Регуляция локального кровотока. Говоря о влияниях трансмиттеров, выделяемых нейронами, необходимо упомянуть еще один эффект. Многочисленные исследования показывают, что различные трансмиттерные системы могут влиять через рецепторы, находящиеся на капиллярах мозга или периваскулярной глии, посредством объемного проведения. Регуляторные влияния показаны в настоящее время для ацетилхолина (через никотиновые рецепторы и, по - видимому, с участием совместно синтезируемой окиси азота), серотонина (через 1А рецепторы), норадреналина, дофамина и глутамата. Существование и значение точной регуляции локального кровотока для осуществления функций мозга, включая высшие когнитивные процессы, стало очевидным в последние годы в связи с широким применением неинвазивных методов (позитронная эмиссионная томография и функциональный магнитный резонанс) для изучения динамической локализации процессов при различных когнитивных тестах. Оба метода основаны на оценке изменения кровотока и уровня оксигенации крови. Тот факт, что многие нейрофармакологические средства могут влиять не только посредством прямого действия на нейрональные рецепторы, но путем изменения кровоснабжения должен учитываться в экспериментальных исследованиях in vivo.
Этот раздел в целом можно заключить словами известного английского нейрохимика Лесли Айверсена: "Ушла в прошлое концепция жестко - связанных нервных цепей с быстрым включением и выключением химических сигналов. Вместо этого наступает эра "химического супа ".
3.3. Медиаторы нервной системы
Развитие представлений о химической медиации нервных импульсов началось в начале века в результате открытий О.Лёви, Дж.Эллиота, Г.Дейла, которые показали, что передача сигнала в нейроэффекторных соединениях опосредуется высвобождением ацетилхолина (АХ) или норадренадина (НА) из
150
нервных окончаний.
До 50-х годов к медиаторам относили две группы низкомолекулярных соединений, которые сейчас называются "классическими", "традиционными" медиаторами, — амины (АХ, адреналин, НА, дофамин, серотонин) и аминокислоты (глутамат, аспартат, ГАМК, глицин), В 60-е годы Дж.Бэрнсток открыл третью группу медиаторов — пуриновые нуклеотиды. В 1953 г. Ф.Лембек выдвинул предположение о медиаторной роли пептида — вещества Р, обнаруженного еще в 30-е годы в мозге в стенках кишечника в виде вещества, которое усиливало сокращения изолированной кишки и вызывало временную гипотензию. Свое название вещество Р получило от слова "powder", поскольку его первооткрыватели работали с высушенными в виде порошка экстрактами тканей. Разработка иммуноцитохимических и радиоиммунологических методов позволила в 70 — 80-е годы выявить в разных отделах нервной системы позвоночных и беспозвоночных множество пептидов (нейропептидов), участвующих в синаптической передаче. Нейропептиды составляют четвертую, самую многочисленную группу медиаторов и, кроме того, выступают как модуляторы действия других медиаторов.
Принцип Дейла. В 30-е годы Г.Дейл пришел к выводу, что идентификация медиатора в периферических окончаниях сенсорного нейрона позволяет судить о природе химической передачи в центральном синапсе этого нейрона. Со временем принцип Дейла стад толковаться как постулат, согласно которому каждый нейрон содержит единственное медиаторное вещество, которое высвобождается во всех окончаниях этого нейрона. В таком понимании принцип Дейла, безусловно, не соответствует действительности.
С современных позиций принцип Дейла соответствует, повидимому, формулировать как положение о метаболической зависимости аксона и его окончаний от тела клетки. Известный нейробиолог Дж.Экклс считает, что именно такой смысл вкладывал в свой вывод и сам Г.Дейл.
Многообразие синаптическях медиаторных функций. В
настоящее время представление о химическом кодировании сигналов в нервной системе основываются на принципе множественности химических сигналов: в индивидуальном нейроне синтезиру-