

системные механизмы
.pdf161
Глава 5 Влияние некоторых лигандов – тиролиберина и
норадреналина на электрогенез.
5.1. Влияние центрального введения ТЛ на электрогенез.
Несомненный интерес представляло исследовать влияние тиролиберина (ТЛ, thyrotropin-releasing hormone, ТЛ) первого выделенного и идентифицированного аденогипофизо-тропного нейрогормона (Burgus R. et al, 1972; Burgus R. et al., 1970; Nair R.M.G. Barret J.G., Bowers C.Y., Schally A.V., 1970) на электрогенез.
Исследование нейрофармакологических характеристик ТЛ показало, что его введение обусловливает выраженные центральные стимулирующие и антидепрессантные эффекты. Это позволило применить ТЛ в клинике в схемах лечения последствий повреждений головного и спинного мозга и депрессий различной этиологии (Ашмарин И. П., Кулашев А.
П., Чепурнов С. А., 1989; Bunevicius R., Matutevicius V., 1993; Horita A. 1998; Iversen E., 1995; Loosen P. T., Prange A. J., Wilson I.C., 1979; Marangell L. B., George M. S., Callahen A. M. et al. 1997). Известно (Ашмарин И. П., Кулашев А. П., Чепурнов С. А., 1989; Iversen E., 1995), что ТЛ, помимо гипоталамуса (где он продуцируется), присутствует в различных отделах головного и спинного мозга, локализуясь главным образом в нервных окончаниях. ТЛ выполняет ряд функций – нейрогормона, модулятора нейронной активности и нейротрофического фактора. Таким образом, каскадные регуляторные процессы, обеспечиваемые ТЛ весьма многообразны (Ашмарин И. П., Кулашев А. П., Чепурнов С. А., 1989). Ситуация с другими короткоживущими эндогенными регуляторными пептидами, видимо, аналогична.
Одно из основных свойств ТЛ, которое привлекало внимание клиницистов, ― его антидепрессивное действие
(Prange A.J., jun., Lara P.P., 1972). Как полагают,
нейрохимические механизмы ТЛ связаны с активацией дофаминэргической системы (Клуша В.Е., Муцениеце Р.К., Свиркис Ш.В., 1982).
С воздействием на нейромедиаторные системы в мозге162 связывают и антинаркотические (аналептические) свойства ТЛ проявляющиеся в виде антагонизма ТЛ с этанолом, барбитуратами, хлоралгидратом (Breese G.R., Cott J.M., Cooper B.R. et al., 1975; Prange A.J, Breese G.R., Cott J.M., vartin B.R., 1974; Prange A.J, et al., 1972).
Аналептическое действие ТЛ связывают с уровнем циклических нуклеотидов в мозге. ТЛ вызывает увеличение цГМФ в мозжечке и антагонизирует его снижение этанолом
(Mailman R.B., Frye G.D., Mueller R.A., Breese G.R., 1972).
При внутрижелудочковом и внутрибрюшинном введении ТЛ вызывает повышение всех биогенных моноаминов и их метаболитов в целом мозге (Зиле Р.К., Одынец Т.Г., Клуша В.Е., 1979).
Согласно данным ряда авторов (Carter C.S., Robertson L.C., Nordahl T.E. et al., 1996; Otto M.W., Yeo R.A., Doughes M.I., 1987), в формировании аффективных расстройств и, в частности, эндогенной депрессии, особо существенную роль играют патологические изменения в нейронных системах правого полушария. Эффекты эндогенных пептидных регуляторов, несмотря на двустороннюю представленность соответствующих нейронных систем, характеризуются той или иной степенью латерализации. В частности, с асимметричным действием этих регуляторов связывают развитие познотонической асимметрии при повреждениях мозжечка (Балабанов Ю.В., Варлинская Е.И., 1980). В то же время вопрос о латерализации влияний нейропептидов на структуры полушарий головного мозга до настоящего времени остается практически открытым.
Задачей настоящего фрагмента работы было выяснение особенностей изменений показателей электрокортикограммы крыс при левоили правостороннем интравентрикулярном введении ТЛ, в частности, выявление возможной латерализации получаемых эффектов. С учетом механизмов продуцирования и движения ликвора, а также анатомии внутримозговых полостей следовало полагать, что введение ТЛ в левый, либо правый боковой желудочек будет обеспечивать преимущественно унилатеральные непосредственные влияния данного агента на нейронные системы полушарий головного мозга.
ТЛ был синтезирован в Физико-химическом институте163 им. А.В. Богатского НАН Украины с использованием ранее описанного метода (Андронати С.А., Мазуров А.А., Коротенко Т.И., 1986). Полученный препарат по своим физикохимическим свойствам полностью соответствовал литературным данным (Schally A.V., Reddings T.W., Bowers
C.Y., Barrett J.F., 1969). Центральное влияние ТЛ исследовали при левостороннем и правостороннем введении.
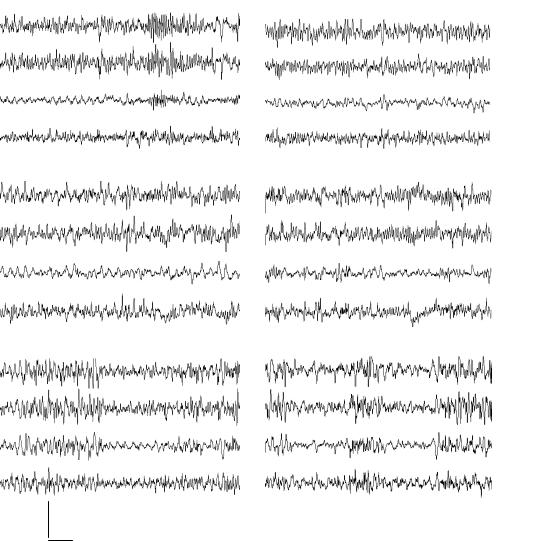
Изменения ЭКоГ после введения ТЛ в левый желудочек мозга. До введения ТЛ (рис. 5.1, А) в ЭКоГ эпизоды десинхронизации чередовались с эпизодами синхронизации, и в целом ЭКоГ носила синхронизированнодесинхронизированный характер. Через 6 минут после введения ТЛ (рис. 5.1, Б) в ЭКоГ преобладала низкоамплитудная дельта активность 1,5-2 в сек. Через 25 минут после введения ТЛ (рис. 5.1) в ЭКоГ отмечалось увеличение амплитуды, особенно в дельта диапазоне и снижение его частоты. Эта тенденция к 30-40 минутам после введения ТЛ (рис. 5.1, Г) возрастала.
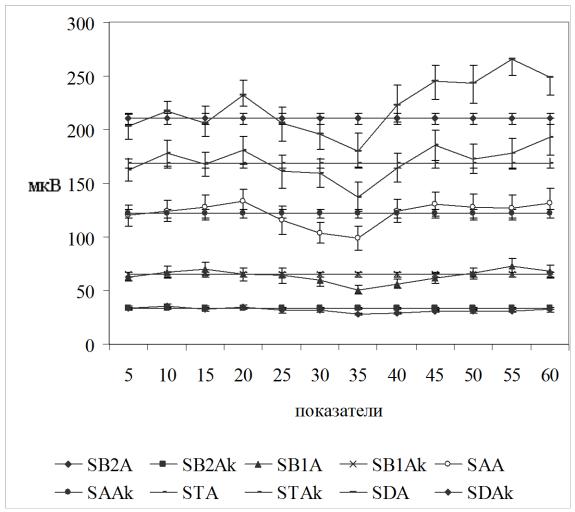
При цифровом анализе за 30-минутный период наблюдения перед введением ТЛ (табл. 5.1., рис. 5,2, 5,3) показатели фоновой ЭКоГ, отводимой от левого и правого полушарий, статистически значимо не изменялись. После введения ТЛ в левый боковой желудочек амплитуда дельтаритма в левом полушарии в первые пять минут после введения несколько уменьшалась, а затем с 15-й по 30-ю мин наблюдения возрастала, превышая исходные значения. Индекс длительности тета-ритма в левом полушарии после левостороннего введения ТЛ в интервале 20–30 мин значимо уменьшался. В правом полушарии наиболее характерным изменением после левостороннего введения ТЛ было уменьшение амплитуды дельта-ритма, наиболее выраженное начиная с пятой и до 25-й мин после введения. Индекс длительности бета-1-ритма в правом полушарии был повышенным на протяжении всех 30 мин после введения. Возрастал и индекс длительности альфа-ритма; этот эффект отмечался в интервале 5–25 мин.
Изменения ЭКоГ после введения ТЛ в правый желудочек мозга. До введения ТЛ (рис. 5.4, А) в ЭКоГ

эпизоды десинхронизации чередовались с эпизодами164 синхронизации,
|
1 |
|
2 |
|
3 |
|
4 |
А |
Б |
В |
Г |
Рисунок 5.1. Влияние на ЭКоГ центрального введения тиролиберина в левый латеральный желудочек мозга крысы.
Обозначения: 1 - фронтальная кора - затылочная кора слева, 2 - фронтальная кора - затылочная кора справа, 3 - фронтальная кора слева
– фронтальная кора справа, 4 -затылочная кора слева – затылочная кора справа. А – до введения, Б – через 362 сек. после введения, В – через 1514 сек. после введения, Г – через 1578 сек. после введения. Калибровка 500 мкВ, отметка времени 2 сек.
регистрировались альфа веретена. Через 2,5 минуты после введения ТЛ (рис. 5.4, Б) в ЭКоГ отмечался сдвиг в сторону десинхронизации. Через 12 минут после введения ТЛ (рис. 5.4, В) в ЭКоГ отмечалось увеличение представленности дельта активности. Через 17 минут после введения ТЛ в ЭКоГ отмечалось снижение представленности дельта активности и увеличение десинхронизации. Через 23 минуты после левостороннего введения ТЛ в ЭКоГ увеличилась представленность альфа ритма, через 30 минут после введения в ЭКоГ восстановился исходный фон. Перед центральным введением ТЛ в правый желудочек мозга почти все показатели фоновой ЭКоГ левого и правого полушарий в течение 30минутного периода наблюдения статистически значимо не менялись (табл. 5.2, рис. 5.5, 5,6).
Исключением было небольшое увеличение индекса длительности альфа-ритма в левом полушарии (на 14 %) и
средней амплитуды дельта-ритма в правом полушарии (на 27165 %) Эти изменения отмечались в конце периода наблюдения фоновой ЭКоГ перед введением гормона (на 25-й–30-й мин).
Рисунок 5.2. Динамика амплитуд ритмов ЭКоГ левого полушария до (5-30 мин) и после (35-60 мин) центрального введения тиролиберина в левый желудочек мозга.
Обозначения: SB2A – амплитуда бета-2-ритма, SB1Aамплитуда бета-1 ритма, SAA – амплитуда альфа ритма, STA – амплитуда тета ритма, SDA – амплитуда дельта ритма. обозначения амплитуд с буквой k отражают базовую величину сравнения – среднюю величину за 30-ти минутный период до введения препарата.
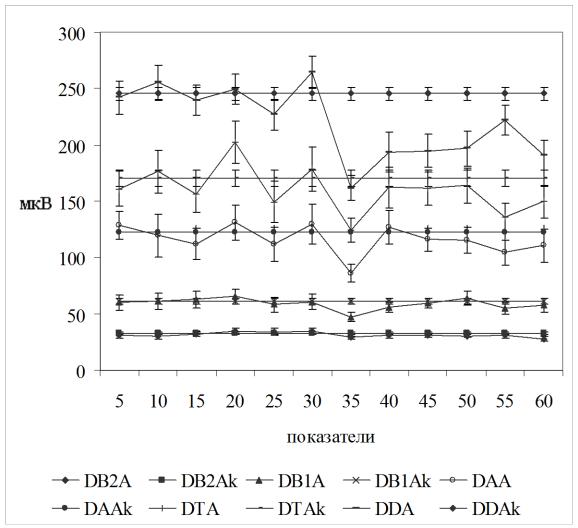
После правостороннего интравентрикулярного введения ТЛ на 5-й мин наблюдения уменьшались индексы длительности бета- 2 и альфа-ритмов и модальная частота бета-2-ритма в левом полушарии. В правом полушарии произошло уменьшение почти всех анализируемых показателей ЭКоГ за исключением
индексов длительности бета-1 и дельта-ритмов. Последний166 показатель значимо возрастал, причем наиболее выраженным
Рисунок 5.3. Динамика амплитуд ритмов ЭКоГ правого полушария до (5-30 мин) и после (35-60 мин) центрального введения TRH в левый желудочек мозга.
Обозначения: DB2A – амплитуда бета-2-ритма, DB1Aамплитуда бета-1 ритма, DAA – амплитуда альфа ритма, DTA
– амплитуда тета ритма, DDA – амплитуда дельта ритма. обозначения амплитуд с буквой k отражают базовую величину сравнения – среднюю величину за 30-ти минутный период до введения препарата.
было увеличение индекса длительности данного низкочастотного компонента ЭКоГ с 20-й по 30-ю мин наблюдения. Необходимо подчеркнуть, что это отмечалось как в левом, так и в правом полушарии.
167
1
2
3
4
А |
Б |
В |
Г |
Д |
Е |
Рисунок 5.4 Влияние на ЭКоГ центрального введения тиролиберина в правый латеральный желудочек мозга крысы. Обозначения: А – до введения, Б – через 150 сек. после введения, В – через 718 сек. после введения, Г – через 1058 сек. после введения, Д – через 1388 сек. после введения. Е – через 1800 сек. после введения. Остальные обозначения теже, что и на 3.1. Калибровка 513 мкВ, отметка времени 2 сек.
5.1.2. Изменения ФМПА после центрального введения ТЛ.
Изменения ФМПА после введения ТЛ в левый желудочек мозга. До центрального левостороннего введения ТЛ (табл. 5.3, рис. 5.7) коэффициенты ФМПА по амплитуде бета-2 альфа тета ритмов ЭКоГ были статистически не значимы, что свидетельствует об отсутствии латерализации.

Амплитуда бета-1 ритма была латерализована в левое, а168 дельта ритма – в правое полушарие. После введения ТЛ коэффициенты ФМПА амплитуд альфа, тета и дельта ритмов
Рисунок 5.5 Динамика амплитуд ритмов ЭКоГ левого полушария до (5-30 мин) и после (35-60 мин) центрального введения тиролиберина в правый желудочек мозга.
Обозначения: те же, что и на рис. 5.2.
ЭКоГ статистически значимо увеличились и были латерализованы в левое полушарие. Показатель ФМПА амплитуды бета-2 ритма также как и до введения ТЛ, был статистически не значим, а положительность показателя ФМПА амплитуды бета-1 ритма, хотя и увеличилась, но статистически не значимо. До введения ТЛ показатели ФМПА по частоте ритмов ЭКоГ были латерализованы в левое полушарие и выражались величинами в пределах 0,66 0,48% – 2,81 0,54%.
Статистически значимо изменились после введения ТЛ показатели ФМПА по частоте тета и дельта ритмов ЭКоГ.

Показатели ФМПА индексов длительности бета-2, бета-1,169 альфа и тета ритмов ЭКоГ до введения ТЛ были латерализованы в левое полушарие и выражались величинами от 5,25 2,83% до 13,65 1,45%, а дельта ритма - в правое (- 13,00 1,56%). После введения ТЛ все показатели МПА
Рисунок 5.6. Динамика амплитуд ритмов ЭКоГ правого полушария до (5-30 мин) и после (35-60 мин) центрального введения тиролиберина в правый желудочек мозга.
Обозначения: те же, что и на рис. 5.3.
индексов длительности статистически значимо уменьшились по модулю. Показатели ФМПА индексов длительности бета-2, бета-1, альфа и дельта ритмов ЭКоГ выражались отрицательными величинами в пределах от (-0,86 0,71%) до (– 3,89 1,9%), т.е. были латерализованы в правое полушарие.
Показатель ФМПА индекса длительности тета ритмвыражался положительной, статистически не значимой величиной –
1,09 0,97%.
Изменения ФМПА после введения ТЛ в правый170
желудочек мозга. До центрального правостороннего введения ТЛ (табл. 5.4, рис. 5.8) коэффициенты МПА по амплитуде бета-2, бета-1, альфа и тета ритмов ЭКоГ выражались отрицательными величинами в пределах (-1,80 1,79% - - 4,36 2,33%.). Коэффициент МПА амплитуды дельта ритма выражался а
Таблица 5.3.
Показатели межполушарной асимметрии ритмов ЭКоГ до и после центрального введения ТЛ в левый желудочек мозга.
Показател |
Периоды наблюдения |
||
и ритмов |
До введения ТЛ |
После введения |
|
ЭКоГ |
|
|
ТЛ |
Бет |
А |
0,99 1,94 |
0,51 1,45 |
а-2 |
Ч |
1,23 0,69 |
-0,21 0,15 |
|
И |
5,25 2,83 |
-3,89 1,9* |
Бет |
А |
2,61 1,13 |
4,72 2,16 |
а-1 |
Ч |
0,66 0,48 |
-0,26 0,61 |
|
И |
7,03 1,93 |
-3,33 1,06* |
Аль |
А |
-0,66 2,48 |
5,87 1,56* |
фа |
Ч |
2,27 0,77 |
0,02 0,67 |
|
И |
11,66 1,62 |
-1,86 1,16* |
Тет |
А |
-0,52 1,74 |
6,9 2,13* |
а |
Ч |
1,88 0,6 |
0,04 0,51* |
|
И |
13,65 1,45 |
1,09 0,97* |
Дел |
А |
-8,02 1,6 |
9,4 1,2* |
ьта |
Ч |
2,81 0,54 |
-0,02 0,69* |
|
И |
-13 1,56 |
-0,86 0,71* |
незначительной, статистически незначимой положительной величиной 0,22 2,37%. После центрального введения ТЛ все коэффициенты МПА по амплитуде ритмов ЭКоГ выражались положительными величинами. Статистически значимыми определялись изменения коэффициентов МПА альфа и тета ритмов.
Показатели МПА частоты ритмов ЭКоГ до введения ТЛ до введения ТЛ были положительны в пределах 1,87 0,82% - 3,72 0,39%. После введения ТЛ эти показатели также определялись положительными в пределах 2,74% 0,69% - 5,00 1,28%. Статистически значимых изменений