

системные механизмы
.pdfкоэффициентов МПА частоты ритмов после центрального171 введения ТЛ не определялось.
15
10
5
% 0
-5
-10
-15
до введения TRH |
после введения TRH |
показатели

 B2A
B2A 
 B1A
B1A  AA
AA 
 TA
TA  DA
DA
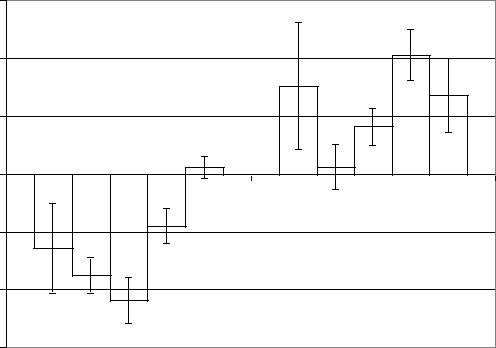
Рис. 5.7. Коэффициенты межполушарной асимметрии по амплитуде до (5-10) и после (15-50) центрального введения TRH в левый латеральный желудочек мозга.
Показатели ФМПА индексов длительности бета-1, альфа, тета и дельта ритмов ЭКоГ до введения ТЛ до введения ТЛ, были положительны в пределах 0,16 1,76% - 4,80 1,82%.
Показатель ФМПА индекса длительности бета-2 ритма до введения ТЛ выражался отрицательной, статистически не значимой величиной 0,1 1,42%. После введения ТЛ все показатели ФМПА индексов длительности определялись положительными в пределах 2,74% 0,69% - 5,00 1,28%. Статистически значимых изменений коэффициентов ФМПА индексов длительности ритмов после центрального введения ТЛ не определялось.
Таблица 5.4172 Показатели межполушарной асимметрии ритмов ЭКоГ до и
после центрального введения TRH в правый желудочек мозга.
Показате |
Периоды наблюдения |
||
ли |
|
До введения |
После введения |
ритмов |
ТЛ |
ТЛ |
|
ЭКоГ |
|
|
|
Бет |
А |
-2,54 1,57 |
3,03 2,2 |
а-2 |
Ч |
1,96 0,69 |
2,74 0,69 |
|
И |
-0,1 1,42 |
0,36 1,31 |
Бет |
А |
-3,5 0,99 |
0,24 2,34 |
а-1 |
Ч |
2,98 0,6 |
3,62 0,78 |
|
И |
0,16 1,76 |
0,67 1,41 |
Ал |
А |
-4,36 2,33 |
1,63 2,88* |
ьфа |
Ч |
1,87 0,82 |
3,28 0,63 |
|
И |
1,19 1,07 |
2,1 1,18 |
Тет |
А |
-1,8 1,79 |
4,11 3,33* |
а |
Ч |
1,82 0,6 |
4,78 0,89 |
|
И |
2,63 1,23 |
4,77 1,32 |
Дел |
А |
0,22 2,37 |
2,7 3,75 |
ьта |
Ч |
3,72 0,39 |
5,00 1,28 |
|
И |
4,8 1,82 |
4,05 2,43 |
5.1.3.Взаимоотношения амплитуд ритмов ЭКоГ до и после центрального введения ТЛ.
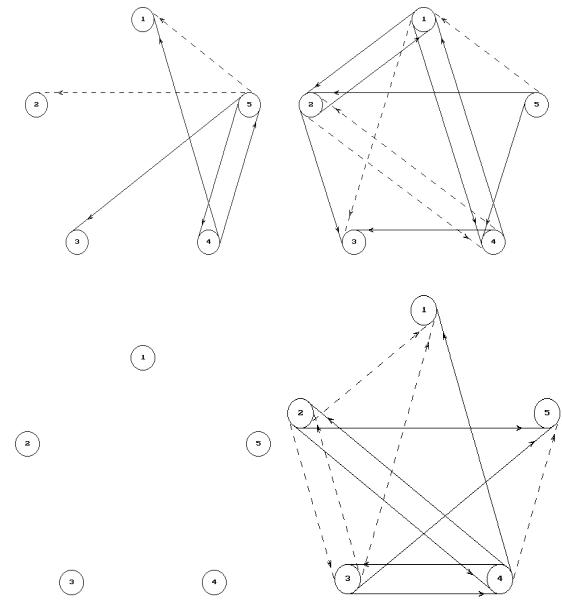
До введения ТЛ в левый желудочек мозга в левом полушарии (рис. 5.9, А. Б) верифицировалось 6 связейотношений, из них 4 положительные и 2 отрицательные. После введения ТЛ количество связей-отношений в левом полушарии увеличилось на 6 связей-отношений и составило 12, из них 8 положительных и 4 отрицательные.
До введения ТЛ в левый желудочек мозга в правом (рис. 5.9, В, Г) полушарии связи не опрделялись. После введения ТЛ связи-отношения в правом полушарии определялись в количестве 12, из них положительных - 7 и отрицательных - 5.
До введения ТЛ в правый желудочек мозга в левом173 полушарии (рис. 5.10, А. Б) верифицировались 3 связиотношения. После
6
4
2
% 0
-2
-4
-6
до введения TRH |
после введения TRH |
показатели

 B2A
B2A  B1A
B1A 
 AA
AA  TA
TA 
 DA
DA
Рис. 5.8. Коэффициенты межполушарной асимметрии по амплитуде до (5-10) и после (15-50) центрального введения TRH в правый латеральный желудочек мозга.
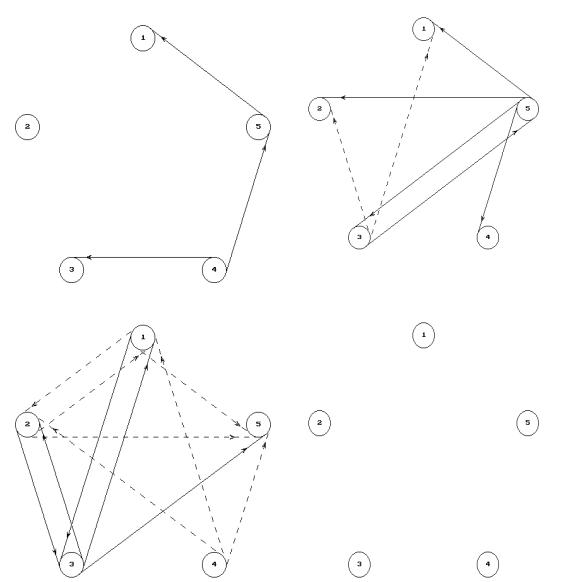
введения ТЛ количество связей-отношений в левом полушарии увеличилось до 12, из них 5 положительных и 7 отрицательных. До введения ТЛ в правый желудочек мозга в правом (рис. 5.10, В, Г) полушарии определялось 12 связейотношений, из них 5 положительных и 7 отрицательных. После введения ТЛ связи-отношения в правом полушарии не определялись.
Таким образом, после центрального введения ТЛ в желудочек левого полушария количество связей-отношений, как в левом, так и в правом полушарии увеличилось. При введении ТЛ в желудочек мозга правого полушария в нем
количество связей-отношений уменьшилось, а в левом174 полушарии, напротив, возросло.
А |
Б |
В |
Г |
Рисунок 5.9. Математические модели отражающая взаимоотношения между амплитудами ритмов ЭКоГ левого и правого полушарий до и после центрального введения ТЛ в левый желудочек мозга.
Обозначения: 1 – бета –2 ритм, 2 – бета 1 ритм, 3 – альфа ритм, 4 – тета ритм, 5 – дельта ритм; А – левое полушарие до введенияя ТЛ, Б - левое полушарие после введенияя ТЛ, В – правое полушарие до введения ТЛ, Г – правое полушарие после введения ТЛ.
175
5.1.4. Изменения коэффициентов активации до и после центрального введения ТЛ.
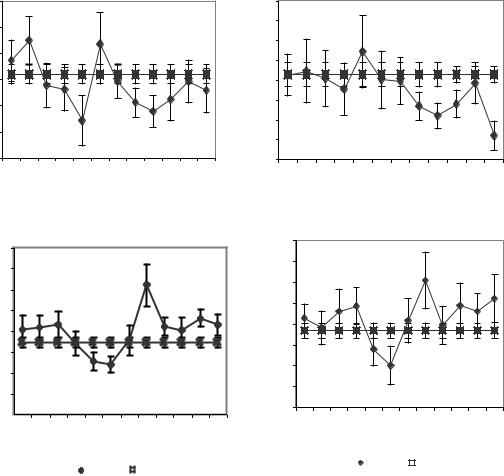
После центрального введения ТЛ в левый желудочек КА в ЭКоГ левого и правого полушария (рис. 5.11) уменьшились,
апосле введения ТЛ в правый желудочек КА в ЭКоГ левого и правого полушария увеличивались.
Резюмируя изложенное можно заключить, что после центрального левостороннего введения ТЛ в левом полушарии уменьшение амплитуд ритмов отмечено только к первому пятиминутному периоду наблюдения. Наиболее постоянным являлось увеличение амплитуды дельта ритма – с 15 по 30 минуты наблюдения. На протяжении всего периода наблюдения после введения ТЛ в левый латеральный желудочек мозга в правом полушарии амплитуда дельта диапазона наблюдалась статистически значимо уменьшенной,
аиндекс длительности бета – 1 ритма – увеличенным. Достаточно выраженным было увеличение индекса длительности альфа-ритма (к 5, 10, 15, 25 и 30 минутам наблюдения).
После центрального правостороннего введения ТЛ, в левом полушарии, наиболее часто определялось увеличение индекса длительности дельта ритма, а в правом полушарии наиболее характерным изменением ЭКоГ являлось уменьшение амплитуд ритмов в первые 10 минут после введения и увеличение индекса длительности дельта ритма к 10, 20, 25 и 30 минутам после введения.
После введения ТЛ в левый боковой желудочек амплитуда дельта-ритма в левом полушарии в первые пять минут после введения несколько уменьшалась, а затем с 15-й по 30-ю мин наблюдения возрастала, превышая исходные значения.
Индекс длительности тета-ритма в левом полушарии после левостороннего введения ТЛ в интервале 20–30 мин значимо уменьшался. В правом полушарии наиболее характерным изменением после левостороннего введения ТЛ было уменьшение амплитуды дельта-ритма, наиболее выраженное начиная с пятой и до 25-й мин после введения.
Индекс длительности бета-1-ритма в правом полушарии был повышенным на протяжении всех 30 мин после введения.
Возрастал и индекс длительности альфа-ритма; этот эффект176 отмечался в интервале 5–25 мин.
А |
Б |
В |
Г |
Рисунок 5.10. Математические модели отражающая взаимоотношения между амплитудами ритмов ЭКоГ левого и правого полушарий до и после центрального введения ТЛ в правый желудочек мозга.
Обозначения: те же, что и на рис. 5.9.
После правостороннего интравентрикулярного введения ТЛ на 5-й мин наблюдения уменьшались индексы длительности бета- 2 и альфа-ритмов и модальная частота бета-2-ритма в левом полушарии. В правом полушарии произошло уменьшение почти всех анализируемых показателей ЭКоГ за исключением индексов длительности бета-1 и дельта-ритмов.
Последний показатель значимо возрастал, причем177 наиболее выраженным было увеличение индекса длительности
0.55 |
|
|
|
|
|
|
|
|
|
|
|
0.50 |
|
|
|
|
|
|
|
|
|
|
|
0.45 |
|
|
|
|
|
|
|
|
|
|
|
КА 0.40 |
|
|
|
|
|
|
|
|
|
|
|
0.35 |
|
|
|
|
|
|
|
|
|
|
|
0.30 |
|
|
|
|
|
|
|
|
|
|
|
0.25 |
|
|
|
|
|
|
|
|
|
|
|
5 |
10 |
15 |
20 |
25 |
30 |
35 |
40 |
45 |
50 |
55 |
60 |
|
|
|
|
|
время (мин) |
|
|
|
|
|
|
|
|
|
|
|
Ska2 |
А |
Ska2k |
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
0,65 |
|
|
|
|
|
|
|
|
|
|
|
0,60 |
|
|
|
|
|
|
|
|
|
|
|
0,55 |
|
|
|
|
|
|
|
|
|
|
|
0,50 |
|
|
|
|
|
|
|
|
|
|
|
КА 0,45 |
|
|
|
|
|
|
|
|
|
|
|
0,40
0,35
0,30
0,25 

5 |
10 |
15 |
20 |
25 |
30 |
35 |
40 |
45 |
50 |
55 |
60 |
|||
|
|
|
|
|
время (мин) |
|
|
|
|
|||||
|
|
|
|
|
Ska2 |
|
|
|
Ska2k |
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
||||
0.65 |
|
|
|
|
|
|
|
|
|
|
|
|
0.60 |
|
|
|
|
|
|
|
|
|
|
|
|
0.55 |
|
|
|
|
|
|
|
|
|
|
|
|
0.50 |
|
|
|
|
|
|
|
|
|
|
|
|
КА 0.45 |
|
|
|
|
|
|
|
|
|
|
|
|
0.40 |
|
|
|
|
|
|
|
|
|
|
|
|
0.35 |
|
|
|
|
|
|
|
|
|
|
|
|
0.30 |
|
|
|
|
|
|
|
|
|
|
|
|
0.25 |
|
|
|
|
|
|
|
|
|
|
|
|
|
5 |
10 |
15 |
20 |
25 |
30 |
35 |
40 |
45 |
50 |
55 |
60 |
|
|
|
|
|
|
время (мин) |
|
|
|
|
||
|
|
|
|
|
|
Dka2 |
|
БDka2k |
|
|
|
|
|
0.65 |
|
|
|
|
|
|
|
|
|
|
|
|
0.60 |
|
|
|
|
|
|
|
|
|
|
|
|
0.55 |
|
|
|
|
|
|
|
|
|
|
|
|
0.50 |
|
|
|
|
|
|
|
|
|
|
|
КА |
0.45 |
|
|
|
|
|
|
|
|
|
|
|
0.40
0.35
0.30
0.25
5 |
10 |
15 |
20 |
25 |
30 |
35 |
40 |
45 |
50 |
55 |
60 |
|||
|
|
|
|
|
время (мин) |
|
|
|
|
|||||
|
|
|
|
|
Dka2 |
|
|
|
Dka2k |
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
||||
В |
Г |
Рисунок 5.11. Динамика КА при центральном введении TRH. Обозначения: ka2 – КА; ka2k – средние величины Ка до введения TRH. Обозначения с литерой S - левое полушарие, D - правое полушарие; обозначения амплитуд с литерой k отражают базовую величину сравнения – среднюю величину за тридцатиминутный период наблюдения до введения препарата. А и Б введение TRH в левый желудочек мозга, В и Г – в правый. Интервал времени 5-30 минут до введения TRH 35-60 после введения
данного низкочастотного компонента ЭКоГ с 20-й по 30-ю мин наблюдения. Необходимо подчеркнуть, что это отмечалось как в левом, так и в правом полушарии.
До введения ТЛ в левый латеральный желудочек мозга показатели ФМПА бета-2, и бета-1 ритмов выражались положительными значениями, а альфа, тета и дельта ритмов – отрицательными. После введения ТЛ все показатели МПА амплитуд ритмов выражались положительными величинами.
Статистически значимо измененными определялись178 показатели ФМПА альфа, тета и дельта амплитуд.
До центрального правостороннего введения TЛ коэффициенты ФМПА по амплитуде бета-2, бета-1, альфа и тета ритмов ЭКоГ выражались отрицательными величинами. Коэффициент ФМПА амплитуды дельта ритма выражался незначительной, статистически незначимой положительной величиной. После центрального введения ТЛ все коэффициенты ФМПА по амплитуде ритмов ЭКоГ выражались положительными величинами. Статистически значимыми определялись изменения коэффициентов МПА альфа и тета ритмов.
Таким образом, показатели ФМПА для амплитуд почти всех ритмов ЭКоГ после введения ТЛ возрастали вне зависимости от стороны введения, причем часто это происходило с инверсией знака. Иначе говоря, асимметрия этих показателей становилась явно левосторонней
При структурном анализе связей-отношений между амплитудами ритмов ЭКоГ определённых использованием множественного регрессионного метода анализа выявилось, что до введения ТЛ у крыс с левосторонней канюлей количество регрессионных связей-отношений преобладало в левом полушарии, а с правосторонней - было таким же, как и у неканюлированных крыс – в правом полушарии определялось больше связей-отношений, чем в левом.
После введения ТЛ в левый желудочек мозга количество связей-отношений, как в левом, так и в правом полушарии увеличилось, и в обеих полушариях было одинаково. При введении ТЛ в желудочек мозга правого полушария в нем количество связей-отношений уменьшилось, а в левом полушарии, напротив, возросло.
По данным иммуногистохимического анализа наибольшее число тиролиберин-нейронов сосредоточено в ядрах переднего гипоталамуса: перивентрикулярном и медиальном мелкоклеточных частях паравентрикулярного ядра (Lechan R.M., Jackson I.M.D., 1982; Nishiyama T., et al.,
1985). Также такие нейроны содержатся в перивентрикулярном, дорсо- и вентромедиальном, супрахиазматическом ядрах, в преоптической и перифорникальной области. Иммуногистохимически
обнаружены два тракта тиреолиберин-волокон,179 проецирующихся в срединное возвышение из паравентрикулярного ядра – латеральный и медиальный
(Nishiyama T., et al., 1985).
Количество тиролиберина, локализованного в гипоталамусе, составляет всего 30-32% от его содержания в мозге. Остальная часть тиролиберина (около 70%) приходится на внегипоталамические области мозга (передний мозг, задняя часть промежуточного мозга, задний мозг, двигательные нейроны спинного мозга, ядра черепных нервов, нейрогипофиз, эпифиз).
Тиролиберин, выявляемый в различных частях ЦНС, не является продуктом секреции клеток гипоталамуса. Помимо своей гипофизарной функции, тиролиберин в других областях ЦНС выполняет роль нейропередатчика или нейромодулятора и оказывает в основном стимулирующее влияние.
Было установлено (Болдырева Г.Н., Шарова Е.В., Добронравова Н.С., 2000), что поражение гипоталамуса сопровождается усилением, главным образом, дистантной синхронизации в коре. Это выражается в генерализации альфаритма, проявляющегося без типичного для ЭЭГ здоровых людей амплитудного градиента в затылочно-лобном направлении. Как полагают авторы, эти изменения обусловлены усилением синхронизирующих влияний на кору со стороны включенных в патологический процесс диэнцефальных структур и сопровождаются ослаблением активирующего влияния на кору со стороны ретикулярной формации мозгового ствола, что в свою очередь, отражает реципрокный характер изменения активности структур, оказывающих синхронизирующий и десинхронизирующий эффект в формировании корковой активности.
В наших опытах после введения ТЛ в левый боковой желудочек увеличение амплитуды дельта-ритма в левом полушарии в сочетании со снижением величин коэффициента активации, свидетельствующего о ослаблении активирующего влияния на кору со стороны ретикулярной формации мозгового ствола, может рассматриваться как усиление ингибиторных влияний. Как известно, синхронизация у крысы реализуется на более низком частотном диапазоне, чем у
человека (Ефремова Т.М., Труш В.Д., 1971; Жадин М.Н. 1984;180 Жадин М.Н., Бахарев Б.В., 1976)
Латерализация амплитуд в левое полушарие свидетельствует об относительном, правого полушария, усилении механизмов синхронизации в левом полушарии. Тем не менее, наблюдалось увеличение регрессионных связейотношений и уравнивание их количества в обеих полушарии. Аналогичная ситуация наблюдается на препарате изолированного переднего мозга. На препарате изолированного переднего мозга, амплитуда ритмов ЭКоГ также латерализована в левое полушарие и количество связейотношений в левом и правом полушарии также одинаково.
Таким образом, можно заключить, что при левостороннем введении ТЛ в ЭКоГ крысы возникают элементы подобия с ЭКоГ препарата изолированного переднего мозга.
Поэтому можно предположить, что центральное применение ТЛ в левый желудочек мозга, в какой-то мере, моделирует гиперактивность гипоталамических структур.
После правостороннего интравентрикулярного введения ТЛ в правом полушарии произошло уменьшение почти всех анализируемых показателей ЭКоГ за исключением индексов длительности бета-1 и дельта-ритмов как в левом, так и в правом полушарии при увеличении показателей коэффициента активации. При этом связи отношения в правом полушарии, т.е. на стороне введения уменьшились а в левом возросли.
Требует дополнительно анализа факт увеличения показателей коэффициента активации в правом полушарии при правостороннем введении ТЛ и уменьшение количества регрессионных связей-отношений, с одной стороны и уменьшение коэффициента активации при левостороннем введении ТЛ и увеличение количества регрессионных связейотношений. Поэтому был проведен двумерный корреляционный анализ синхронности изменений показателей амплитуды ЭКоГ в левом и правом полушариях при лево- и правостороннем применении ТЛ.
При расчете коэффициентов двумерной корреляции между амплитудами ритмов ЭКоГ при левостороннем введении ТЛ в левом полушарии определялось до введения 3 статистически значимых коэффициента корреляции, а после