

системные механизмы
.pdfсинхрониизации определялось по 16 связей-отношений.211 После введении ПФв в левом полушарии в периоды десинхронизации определялось 18, а синхронизации – 14 связей-отношений.
Таким образом, в условиях несегментированной ЭКоГ количество определяемых связей-отношений после применения ПФв уменьшается, а при сегментации, хоть и незначительно, увеличивается в правом полушарии.
Можно высказать предположение, что при применении множественного регрессионного анализа в условиях несегментированной ЭКоГ происходит искажение реальной ситуации в связи с неравнозначностью периодов синхронизации и десинхронизации.
Как известно, полифенольные вещества имеют антиоксидантну действие (Барабой В.А., 1965;. Барабой В.А., 1968; Барабой В.А., Сутковский Д.А., 1997). Система биологически активных фенолов, к которым відносят аминокислоти тирозин, триптофан, катехоламини адреналин и норадреналин функционируют, как одно целое и сыграют роль буферной антиоксидантной системы, так как, в физиологических условиях равновесие внутри нее сдвинуто влево (Барабой В.А., Сутковский Д.А. 1997)
Можно предположить что антиоксидантное действие полифенолов, в какой-то мере, реализуеться через активацию норадренергичних систем.
Многократно предпринимались попытки объяснить действие растительных полифенолов их адреналоподобным действием (Барабой, 1968). Как полагают адреналиноподобное действие фенолов может быть обусловлено: 1) стабилизацией эндогенного или введенного одновременно извне адреналина; 2) стимуляцией мозгового вещества надпочечников и окончаний нервной системы, с освобождением и выходом в кровь пирокатехинаминов; 3) блокадой путей инактивации адреналина; 4) непосредственным адреналиноподобным действием на клетки и ткани (Барабой, 1968).
В последнее время показано, что цис и транс-resveratrol (c-RESV и t-RESV), а также (-)-trans-epsilon-viniferin (эпсилонviniferin), димер ресвератрола, в концентрации 5-200 microM дозозависимо ингибировали обратный захват (3H) норадреналин и (3H) 5-HT в синаптосомах мозга крысы и
обратный захват (3H) 5-HT человеческими пластинками212
(тромбоцитами). c-RESV, t-RESV и эпсилон-viniferin в
концентрации 5-200 microM дозозависимо ингибировал ферментативную активность коммерческой - человеческий рекомбинантный ген - моноаминоксидазы (MAO) изоформы MAO-A и MAO. Выявленные особенности в действии исследованного препарата позволяют отнести его к антидепрессантам, принимая во внимание, что основные группы антидепрессантов реализуют обратный захват noradrenaline/5- HT или ингибицию MAO. В данном случае эти два важных биохимических действия объединены в одной химической структуре (Yanez M, Fraiz N, Cano E, Orallo F. 2006a; Yanez M, Fraiz N , Cano E, Orallo F. 2006b)
В этой связи представляло интерес сопоставить эффекты внутрибрюшинного введения ПФв и центрального введения НА.
6.2. Нейрофизиологический анализ нейрохимических механизмов действия ПФв
В ЭКоГ левых полушарий после применения ПФв и НА (табл. 6.6) статистически значимо отрицательно коррелировали при Р<0,05 изменения амплитуд бета-2 и альфа ритмов, а также дельта ритма, но при 0,05<Р<0,1. В правом полушарии статистически значимо положительно коррелировали только изменения частот дельта ритма при 0,05<Р<0,1.
Квадрат расстояния Маханолобиса между разностными матрицами показателей левых полушарий после применения ПФв и НА составлял 2,00 при Р=0,11, а правых 2,29 при Р=0,068. Отсутствие статистически значимых различий (Р>0,1) квадрата расстояния Маханолобиса между матрицами данных свидетельствует о подобии показателей ЭКоГ, и наоборот, статистически значимое различие (P<0,1) матриц данных, судя по квадрату расстояния Маханолобиса, свидетельствует об отсутствии подобия.
Таким образом, по результатам дискриминантного анализа выявляется подобие в изменении показателей ЭКоГ после применения ПФв и НА в левых полушариях и отсутствие подобия в изменении показателей ЭКоГ правых полушарий.
При исследовании подобия-различия изменений показателей ЭКоГ после центрального левостороннего введении

ТЛ и ПФв (табл. 6.6) в левом полушарии статистически213 значимо положительно коррелировали при Р<0,05 изменения индексов длительности бета-2 и альфа ритмов и статистически значимо при 0,05<Р<0,1 изменения амплитуд бета-1 ритма. Изменения амплитуды бета-2 и индекса длительности бета-1 ритма коррелировали статистически значимо при Р<0,05 отрицательно. В правом
Таблица 6.6. Результаты корреляционного анализа изменений ЭКоГ при
внутрибрюшинном применении ПФв и центральном ТЛ и NA.
Центрально примененные препараты сравнения
|
|
|
ТЛ |
|
|
|
|
|
||
|
|
Левостороннее |
Правосторон-нее |
|
NA |
|
||||
|
|
введение |
|
введение |
|
|
|
|
||
Показатели |
|
|
|
Полушария |
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
ЭКоГ |
|
|
Пра- |
|
|
|
|
|
|
Пра- |
|
|
Левое |
вое |
|
Левое |
Пра-вое |
|
Левое |
|
вое |
|
|
|
|
|
|
|
|
|
|
|
|
А |
-0,90** |
-0,42 |
|
0,62 |
0,41 |
|
-0,86** |
|
0,24 |
Бета –2 |
Ч |
0,39 |
0,43 |
|
0,26 |
0,18 |
|
-0,54 |
|
-0,6 |
|
И |
0,78** |
0,25 |
|
0,27 |
0,07 |
|
-0,55 |
|
-0,57 |
|
А |
-0,72* |
-0,51 |
|
-0,97** |
0,11 |
|
-0,65 |
|
-0,11 |
Бета – 1 |
Ч |
0,29 |
0,41 |
|
0,31 |
0,25 |
|
0,58 |
|
0,62 |
|
И |
-0,89** |
-0,53 |
|
0,54 |
0,42 |
|
0,39 |
|
-0,27 |
Альфа |
А |
0,2 |
0,08 |
|
-0,85** |
0,35 |
|
-0,76** |
|
0,27 |
|
Ч |
0,47 |
-0,05 |
|
-0,08 |
-0,15 |
|
0,49 |
|
0,07 |
|
И |
0,78** |
-0,54 |
|
0,20 |
0,17 |
|
0,26 |
|
0,27 |
Тета |
А |
0,13 |
-0,58 |
|
-0,63 |
0,1 |
|
-0,55 |
|
-0,1 |
|
Ч |
0,21 |
-0,02 |
|
0,33 |
-0,07 |
|
0,02 |
|
0,4 |
|
И |
0,47 |
0,86** |
|
0,70* |
-0,76** |
|
-0,15 |
|
-0,51 |
|
А |
-0,61 |
-0,05 |
|
-0,57 |
0,06 |
|
-0,68* |
|
0,09 |
Дельта |
Ч |
0,28 |
-0,19 |
|
0,27 |
-0,31 |
|
0,52 |
|
0,70* |
|
И |
-0,35 |
0,66 |
|
-0,68* |
-0,43 |
|
0,57 |
|
-0,19 |
полушарии статистически значимо положительно (при Р<0,05) коррелировали изменения индексов длительности тета ритма ЭКоГ.
При центральном правостороннем введении ТЛ в левом полушарии статистически значимо отрицательно
коррелировали изменения амплитуд бета-1, и альфа ритмов.214 В правом полушарии статистически значимо отрицательно коррелировали изменения индексов длительности тета ритма.
Обращает на себя внимание, тот факт, что не зависимо от стороны введения ТЛ большинство корреляционных отношений показателей ЭКоГ после применения ПФв и ТЛ выявлялись в левом полушарии - 6, а в правом только 1.
Таким образом, при применении ПФв и ТЛ подобно изменялись во времени три показателя ЭКоГ (положительные коэффициенты корреляции), а пять показателей ЭКоГ коррелировали отрицательно. Следует отметить, что при левостороннем введении ТЛ три показателя коррелировали положительно, а два отрицательно, а при правостороннем введении ТЛ положительных корреляций выявлено не было.
Квадраты расстояний Маханолобиса при левостороннем центральном введении ТЛ и ПФв между разностными матрицами показателей ЭКоГ составили между левыми полушариями 1,51 при Р=0,22, а между правыми полушариями 1,42 при Р=0,25, т.е. изменения показателей в своей совокупности не различались.
При центральном правостороннем введении ТЛ и ПФв квадрат расстояний Маханолобиса между левыми полушариями составил 2,76 при Р=0,035, а между правыми полушариями – 2,16, при Р=0,09, т.е. изменения показателей в своей совокупности были подобны при Р<0,1.
Таким образом, при левостороннем центральном введении ТЛ изменения ЭКоГ в обоих полушариях в целом соответствовали таковым при введении ПФв, а при правостороннем не соответствовали.
Полученные результаты могут свидетельствовать о неравноценности в физиологическом взаимодействии ПФв с ТЛ продуцируемым левой и правой половинами гипоталамуса.
В целом, полученные результаты позволяют высказать гипотезу, что нейрофизиологическое действие ПФв может реализовываться через стимуляцию норадренергических и тиролиберинэргических нейрохимических механизмов.
215
6.3. Изучение влияния ПФв в условиях ИПМ
Основное действующее начало ПФв - растительные полифенолы у которых выявлено антиоксидантное действие (Г.А. Барабой, 1990; Г.А Барабой., Орел., И.М Карнаух., 1991; Г.А Барабой., Д.А. Сутковский, 1997). Развитие антиоксидантного действия сопровождается активацией структур антиэпилептической системы (L.S Godlewsky., A.A Shandra., A.A Oleinik., et all, 2002), активация которых вызывает десинхронизацию ЭКоГ (Крижановский Г.Н., Тупеев И.Р., Никушкин Е.Г., 1987). Одним из важных звеньев антиэпилептической системы является мозжечок (Г.Н. Крыжановский, Р.Ф. Макулькин, Б.А., Лобасюк 1981; Г.Н. Крыжановский, А.А. Шандра, Л.С. Годлевский, 1990).
Поэтому, для целей выявления нейрофизиологических механизмов действия ПФв, целесообразно исследовать влияние ПФв на ЭКоГ в условиях ИПМ и удаления полушарий мозжечка.
В связи с тем, что исследование влияние ПФв на ЭКоГ ИПМ и АП проводилось с применением процедуры сегментации, исследовали влияние ПФв на ЭКоГ у интактных животных также в условиях сегментации.
Следует подчеркнуть, что исследование влияния ПФв на ЭКоГ АП без применения сегментации не представляется возможным.
6.3.1. Сопоставление показателей ЭКоГ, отводимых от препаратов ИМ и ИПМ при применении ПФв.
Анализ влияния на ЭКоГ ПФв в условиях ИПМ проводился в условиях несегментированной ЭКоГ и при использовании сегментации. Мы полагаем, что использование этих двух подходов позволит более корректно проанализировать данную экспериментальную ситуацию.
До введения ПФв в ЭКоГ крысы (рис. 6.9, А) преобладала синхронизация в альфа диапазоне. К 5 минуте после введения ПФв (рис. 6.9, Б) увеличилось количество возникающих альфаверетен, их длительность. В последующие периоды выраженность альфа активности не отличалась от исходного фона.
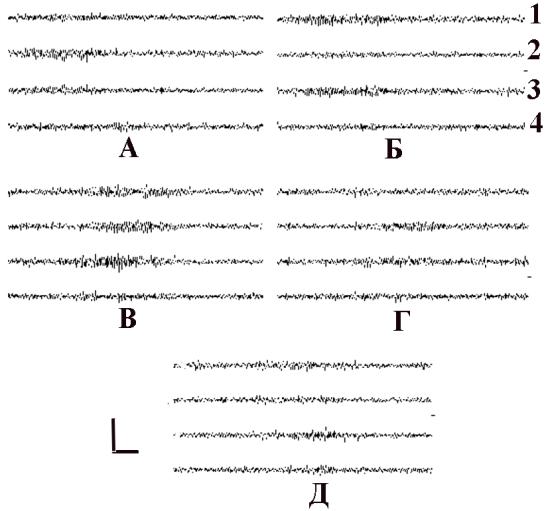
Сопоставление показателей несегментированных216 ЭКоГ до и после применения ПФв у ИПМ. На препарате ИПМ (табл 6.7, рис. 6.10, 6.11) амплитуды, частоты и индексы дллительности бета-2, бета-1 и альфа ритма к пятой минуте наблюдение после введения ПФв наблюдались статистически значимо увеличенными в правом полушарии, а в левом - статистически значимо не измененными.
К 10 минуте наблюдения в правом полушарии статистически значимо увеличенною определялись только амплитуда, частота и индекс длительности бета-2 ритма, а в левом полушарии статистически значимо измененных показателей ритмов ЭКоГ не
Рис. 6.9. Влияние на ЭКоГ ИПМ внутрибрюшинного введения ПФв.
Обозначения: А – ЭКоГ до введения, Б, В, Г, Д – через 5, 15, 20 и 30 минут после введения ПФв. Отметка времени 1 сек., калибровка 517 мкв.
определялось. К 15 минуте наблюдения в левом полушарии217 статистически значимо уменьшенными определялись показатели амплитуды бета-1 и альфа ритма, частоты бета-2, бета-1 и альфа ритма, индекс длительности бета-1 ритма, частота и индекс длитеельности тета ритма, частота дельта ритма. К 15 и 20 минутам после введения ПФв в правом полушарии, статистически значимо измененным, увеличенным, определялся только индекс длительности бета-2 ритма. С 20 по 40 минуты наблюдения в левом полушарии статистически значимо уменьшились показатели амплитуды, частоты и индекса длительности бета-2 ритма. Амплитуда бета-1 ритма определялась уменьшенною к 20, 25, 35 и 40-й минуте после введения ПФв, частота бета-1 ритма - к 20, 30 и 35 минутам после введения ПФР, а индекс длительности бета-1 ритма с 20 по 50 минуту наблюдения.
Остальные показатели ЭКоГ статистически значимо измененными определялись спорадически: к 40 и 50 минутам в левом полушарии частота альфа ритма, к 35 минуте в левом полушарии увеличилась частота дельта ритма, и к 50 минуте наблюдения в правом полушарии увеличилась амплитуда дельта ритма.
Сопоставление показателей сегментированных ЭКоГ ИПМ до и после применения ПФв. До введения ПФв в левом полушарии ЭКоГ крысы периоды синхронизации составляли 13,41% от общего времени регистрации, а в правом полушарии
– 26,67%. После введения ПФв в левом полушарии ЭКоГ крысы периоды синхронизации составляли 13,62% от общего времени регистрации, а в правом полушарии –5,98 %.
В ЭКоГ левого полушария во фрагментах синхронизации после введения ПФв (табл. 6.8) амплитуды бета-1, бета-2, альфа и тета ритмов ЭКоГ определялись большими, чем во фрагментах десинхронизации. Амплитуда дельта ритма во фрагментах синхронизации левого полушария определялась большей, а правого - меньшей, чем во фрагментах десинхронизации. Частоты и индексы длительности бета-2 и бета-1 ритмов левого и правого полушарий и частота и индекс длительности альфа ритма в правом полушарии, а также индекс длительности дельта ритма левого полушария определялись во фрагментах синхронизации увеличенными по сравнению с аналогичными величинами фрагментов десинхронизации.
Индексы длительности тета ритма левого полушария и218 дельта ритма правого определялись во фрагментах синхронизации уменьшенными по сравнению с аналогичными величинами фрагментов десинхронизации.
Во фрагментах десинхронизации после введения ПФв амплитуды бета-2, бета-1, ритмов левого полушария определялись уменьшенными а тета и дельта ритмов – увеличенными, а в правом, наоборот - амплитуды бета-2, бета-1 и альфа ритмов определялись увеличенными, а тета и дельта ритмов – уменьшенными по сравнению с фрагментами
180 |
|
|
|
|
|
|
|
|
|
|
|
160 |
|
|
|
|
|
|
|
|
|
|
|
140 |
|
|
|
|
|
|
|
|
|
|
|
120 |
|
|
|
|
|
|
|
|
|
|
|
100 |
|
|
|
|
|
|
|
|
|
|
|
мкВ |
|
|
|
|
|
|
|
|
|
|
|
80 |
|
|
|
|
|
|
|
|
|
|
|
60 |
|
|
|
|
|
|
|
|
|
|
|
40 |
|
|
|
|
|
|
|
|
|
|
|
20 |
|
|
|
|
|
|
|
|
|
|
|
0 |
|
|
|
|
|
|
|
|
|
|
|
5 |
10 |
15 |
20 |
25 |
30 |
35 |
40 |
45 |
50 |
55 |
60 |
|
|
|
|
|
время (мин) |
|
|
|
|
|
|
|
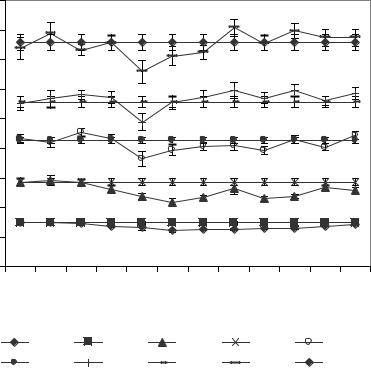
SB2A |
|
SB2A |
|
SB1A |
|
SB1A |
SAA |
|||
|
SAA |
|
STA |
|
|
STA |
|
SDA |
|
SDA |
|
Рис. 6.10. Динамика амплитуд ритмов ЭКоГ левого полушария ИПМ до (5 и 10 минуты) и после (15-60 минуты) введения ПФв.
десинхронизации до введения ПФГ. Частоты и индексы длительности бета-2, бета-1 и альфа ритмов ЭКоГ в левом полущарии после введения ПФв во фрагментах десинхронизации определялись уменьшенными, а в правом полушарии – увеличенными, по сравнению, с аналогичными фрагментами ЭКоГ до введения ПФГ. Частота и ндекс длительности тета ритма в левом полушарии, также определялись увеличенными, а индекс длительности дельта ритма в правом полушарии – уменьшенным.
219
140 |
|
|
|
|
|
|
|
|
|
|
|
120 |
|
|
|
|
|
|
|
|
|
|
|
100 |
|
|
|
|
|
|
|
|
|
|
|
80 |
|
|
|
|
|
|
|
|
|
|
|
мкВ |
|
|
|
|
|
|
|
|
|
|
|
60 |
|
|
|
|
|
|
|
|
|
|
|
40 |
|
|
|
|
|
|
|
|
|
|
|
20 |
|
|
|
|
|
|
|
|
|
|
|
0 |
|
|
|
|
|
|
|
|
|
|
|
5 |
10 |
15 |
20 |
25 |
30 |
35 |
40 |
45 |
50 |
55 |
60 |
|
|
|
|
время (мин) |
|
|
|
|
|||
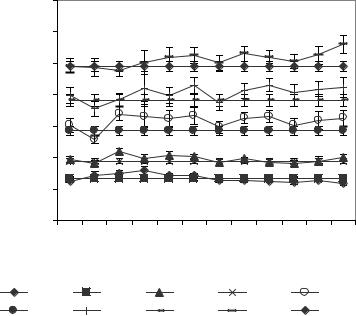
DB2A |
|
DB2Ak |
|
DB1A |
|
DB1Ak |
|
DAA |
|||
DAAk |
|
DTA |
|
|
DTAk |
|
DDA |
|
DDAk |
||
Рис. 6.11. Динамика амплитуд ритмов ЭКоГ правого полушария ИПМ до (5 и 10 минуты) и после (15-60 минуты) введения ПФв.
Во фрагментах синхронизации, также как и во фрагментах десинхронизации, после введения ПФв амплитуды бета-2, бета-1 и альфа ритмов левого полушария определялись уменьшенными а тета и дельта ритмов – увеличенными, а в правом, наоборот - амплитуды бета-2, бета-1 и альфа ритмов определялись увеличенными, а тета и дельта ритмов – уменьшенными по сравнению с фрагментами синхронизации до введения ПФв. Частоты и индексы длительности бета-2 и бета-1 ритмов, а также индекс длительности альфа ритма ЭКоГ в левом полущарии после введения ПФв во фрагментах десинхронизации определялись уменьшенными, а в правом полушарии – увеличенными, по сравнению, с аналогичными фрагментами ЭКоГ до введения ПФГ. Частота альфа ритма определялась увеличенной в правом полушарии, а частота дельта ритма - сниженной, также, в правом полушарии.
6.3.2. Изменения индексов ФМПА ЭКоГ при применении ПФв в условиях ИПМ.
ФМПА показателей ЭКоГ ИПМ без применения процедуры сегментации. Все показатели ФМПА амплитуды ИПМ (табл. 6.9, рис 6.12) были положительные в границах от 5,65±0,10% до 21,36±0,50%. После введения ПФР показатели

ФМПА амплитуд бета-1, альфа, тета и дельта ритма220 статистически значим уменьшили позитивность, а показатель ФМПА бета-2 ритма определялся отрицательным, но статистически незначимым.
Показатели ФМПА частот ритмов ЭКоГ препарата излолированного мозга до введенния ПФв также как и показатели ФМПА амплитуд ритмов, были положительными в границах от 0,74±0,0001% до 16,57±0,01%. После введения ПФв показатели ФМПА частот альфа, и дельта ритма статистически значим уменьшили позитивность, а показатель ФМПА бета-2 ритма определялся статистически значимо отрицательным. Изменения показателей частоты ФМПА бета-1 и тета ритмов определялись статистически не значимыми.
Таблица 6.9.
Показатели межполушарной асимметрии ритмов ЭКоГ ИПМ до и после введения ПФв (без применения сегментации).
Показате |
Периоды наблюдения |
||
ли |
|
|
|
ритмов |
До введения |
После |
|
ЭКоГ |
|
ПФв |
введения ПФв |
Бета-2 |
А |
5,65±0,01 |
-0,08±1,66* |
|
Ч |
0,74±0,0001 |
-2,89±0,92* |
|
И |
20,3±0,18 |
2,83±2,40* |
Бета-1 |
А |
20,52±0,06 |
11,83±1,23* |
|
Ч |
3,58±0,0001 |
1,87±0,97 |
|
И |
18,44±0,05 |
10,09±1,31* |
Альфа |
А |
19,9±0,07 |
12,22±1,14* |
|
Ч |
8,1±0,01 |
3,71±1,11* |
|
И |
15,7±0,08 |
12,45±0,89* |
Тета |
А |
18,79±0,06 |
16,16±1,08* |
|
Ч |
6,27±0,0001 |
4,91±0,79 |
|
И |
7,89±0,04 |
6,84±1,25 |
Дель- |
А |
21,36±0,05 |
18,32±1,01* |
та |
Ч |
16,57±0,01 |
13,71±1,04* |
|
И |
-6,87±0,02 |
-8,35±1,29 |
Примечание: * P<0,05